Optimising Cognitive Enhancement: Systematic Assessment of the Effects of tDCS Duration in Older Adults

{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
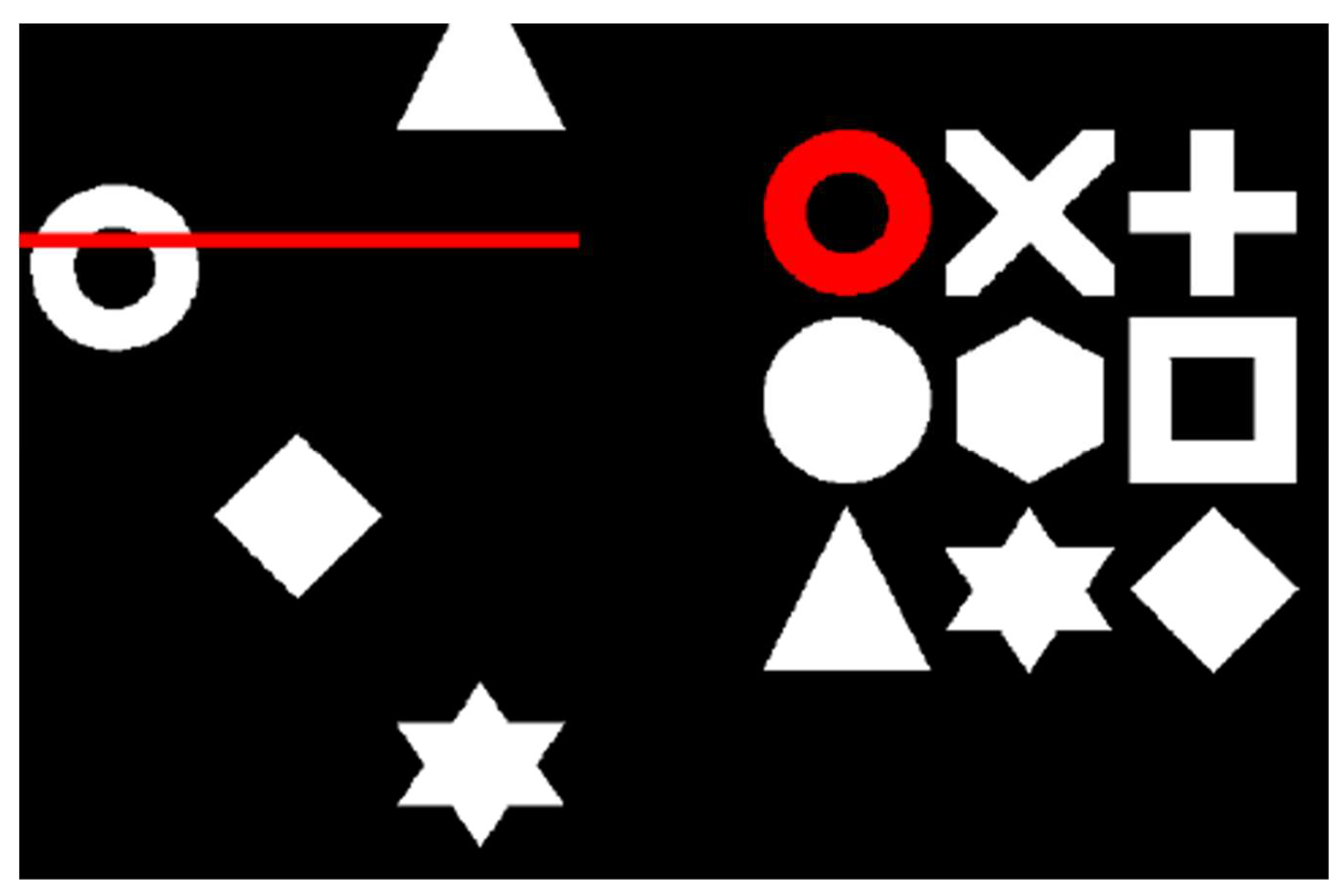
2.2. Task-Switching Paradigm
2.3. Transcranial Direct Current Stimulation
2.4. Experimental Procedure
2.5. Data Analysis
3. Results
3.1. Adverse Effects Questionnaire
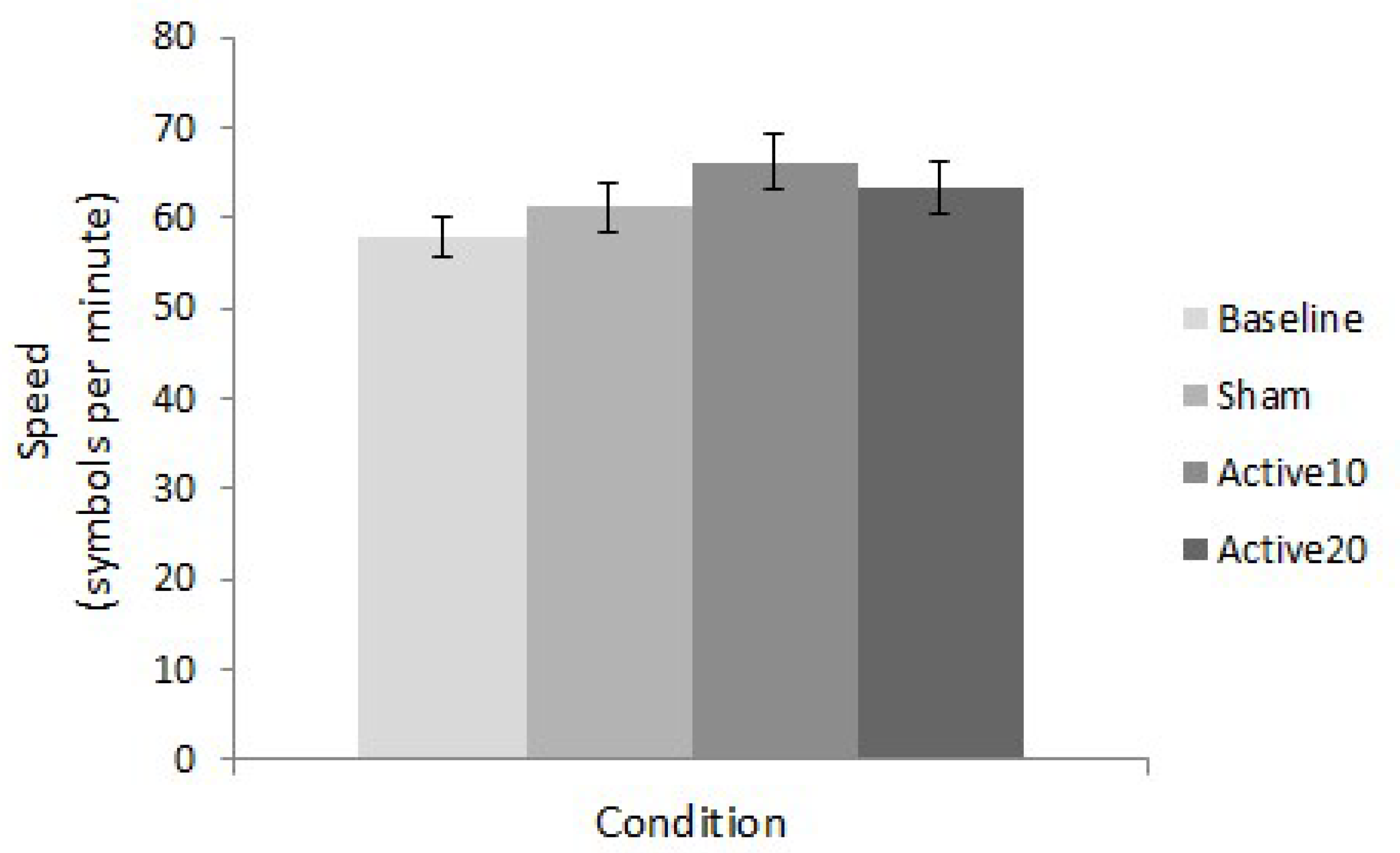
3.2. Task-Switching Speed
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sylvester, C.-Y.C.; Wager, T.; Lacey, S.C.; Hernandez, L.; Nichols, T.E.; E Smith, E.; Jonides, J. Switching attention and resolving interference: fMRI measures of executive functions. Neuropsychologia 2003, 41, 357–370. [Google Scholar] [CrossRef]
- Cieslik, E.C.; Mueller, V.I.; Eickhoff, C.R.; Langner, R.; Eickhoff, S.B. Three key regions for supervisory attentional control: Evidence from neuroimaging meta-analyses. Neurosci. Biobehav. Rev. 2015, 48, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Logsdon, R.G.; Gibbons, L.E.; McCurry, S.M.; Teri, L. Assessing quality of life in older adults with cognitive impairment. Psychosom. Med. 2002, 64, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Emilien, G.; Durlach, C.; Minaker, K.L.; Winblad, B.; Gauthier, S.; Maloteaux, J.-M. Mild Cognitive Impairment. Lancet 2006, 367, 1262–1270. [Google Scholar]
- Shimada, H.; Makizako, H.; Doi, T.; Lee, S.; Tsutsumimoto, K.; Hotta, R.; Bae, S.; Nakakubo, S.; Harada, K.; Suzuki, T. Impact of cognitive frailty on daily activities in older persons. J. Nutr. Health Aging 2016, 20, 729–735. [Google Scholar] [CrossRef]
- Christensen, K.; Doblhammer, G.; Rau, R.; Vaupel, J.W. Ageing populations: The challenges ahead. Lancet 2009, 374, 1196–1208. [Google Scholar] [CrossRef]
- Howdon, D.; Rice, N. Health care expenditures, age, proximity to death and morbidity: Implications for an ageing population. J. Health Econ. 2018, 57, 60–74. [Google Scholar] [CrossRef]
- Whalley, L.J.; Deary, I.J.; Appleton, C.L.; Starr, J.M. Cognitive reserve and the neurobiology of cognitive aging. Ageing Res. Rev. 2004, 3, 369–382. [Google Scholar] [CrossRef]
- Fritsch, T.; McClendon, M.J.; Smyth, K.A.; Lerner, A.J.; Friedland, R.P.; Larsen, J.D. Cognitive functioning in healthy aging: The role of reserve and lifestyle factors early in life. Gerontologist 2007, 47, 307–322. [Google Scholar] [CrossRef]
- Kane, M.R.L.; Butler, P.M.; Fink, M.H.A. Interventions to prevent age-related cognitive decline, mild cognitive impairment, and clinical Alzheimer’s-type dementia. J. Comp. Eff. Rev. 2017, 188. [Google Scholar] [CrossRef]
- Northey, J.M.; Cherbuin, N.; Pumpa, K.L.; Smee, D.J.; Rattray, B. Exercise interventions for cognitive function in adults older than 50: A systematic review with meta-analysis. Br. J. Sports Med. 2018, 52, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Pisoni, A.; Mattavelli, G.; Papagno, C.; Rosanova, M.; Casali, A.G.; Romero Lauro, L.J. Cognitive enhancement induced by anodal tDCS drives circuit-specific cortical plasticity. Cereb. Cortex 2018, 28, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Meinzer, M.; Lindenberg, R.; Antonenko, D.; Flaisch, T.; Flöel, A. Anodal transcranial direct current stimulation temporarily reverses age-associated cognitive decline and functional brain activity changes. J. Neurosci. 2013, 33, 12470–12478. [Google Scholar] [CrossRef] [PubMed]
- Antonenko, D.; Nierhaus, T.; Meinzer, M.; Prehn, K.; Thielscher, A.; Ittermann, B.; Flöel, A. Age-dependent effects of brain stimulation on network centrality. NeuroImage 2018, 176, 71–82. [Google Scholar] [CrossRef]
- Antonenko, D.; Schubert, F.; Bohm, F.; Ittermann, B.; Aydin, S.; Hayek, D.; Grittner, U.; Flöel, A. tDCS-induced modulation of GABA levels and resting-state functional connectivity in older adults. J. Neurosci. 2017, 37, 4065–4073. [Google Scholar] [CrossRef]
- Porges, E.C.; Woods, A.J.; Edden, R.A.; Puts, N.A.; Harris, A.D.; Chen, H.; Garcia, A.M.; Seider, T.R.; Lamb, D.G.; Williamson, J.B.; et al. Frontal gamma-aminobutyric acid concentrations are associated with cognitive performance in older adults. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 38–44. [Google Scholar] [CrossRef]
- Park, S.H.; Seo, J.H.; Kim, Y.H.; Ko, M.H. Long-term effects of transcranial direct current stimulation combined with computer-assisted cognitive training in healthy older adults. Neuroreport 2014, 25, 122–126. [Google Scholar] [CrossRef]
- Jones, K.T.; Gözenman, F.; Berryhill, M.E. The strategy and motivational influences on the beneficial effect of neurostimulation: A tDCS and fNIRS study. NeuroImage 2015, 105, 238–247. [Google Scholar] [CrossRef]
- Stephens, J.A.; Berryhill, M.E. Older adults improve on everyday tasks after working memory training and neurostimulation. Brain Stimul. 2016, 9, 553–559. [Google Scholar] [CrossRef]
- Boggio, P.S.; Campanhã, C.; Valasek, C.A.; Fecteau, S.; Pascual-Leone, A.; Fregni, F. Modulation of decision-making in a gambling task in older adults with transcranial direct current stimulation. Eur. J. Neurosci. 2010, 31, 593–597. [Google Scholar] [CrossRef]
- Harty, S.; Robertson, I.H.; Miniussi, C.; Sheehy, O.C.; Devine, C.A.; McCreery, S.; O’Connell, R.G. Transcranial direct current stimulation over right dorsolateral prefrontal cortex enhances error awareness in older age. J. Neurosci. 2014, 34, 3646–3652. [Google Scholar] [CrossRef] [PubMed]
- Hanley, C.J.; Tales, A. Anodal tDCS improves attentional control in older adults. Exp. Gerontol. 2019, 115, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Batsikadze, G.; Moliadze, V.; Paulus, W.; Kuo, M.F.; Nitsche, M.A. Partially non-linear stimulation intensity-dependent effects of direct current stimulation on motor cortex excitability in humans. J. Physiol. 2013, 591, 1987–2000. [Google Scholar] [CrossRef] [PubMed]
- Jamil, A.; Batsikadze, G.; Kuo, H.-I.; Labruna, L.; Hasan, A.; Paulus, W.; Nitsche, M.A. Systematic evaluation of the impact of stimulation intensity on neuroplastic after-effects induced by transcranial direct current stimulation. J. Physiol. 2017, 595, 1273–1288. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilpour, Z.; Marangolo, P.; Hampstead, B.M.; Bestmann, S.; Galletta, E.; Knotkova, H.; Bikson, M. Incomplete evidence that increasing current intensity of tDCS boosts outcomes. Brain Stimul. 2018, 11, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Laakso, I.; Tanaka, S.; Koyama, S.; De Santis, V.; Hirata, A. Inter-subject variability in electric fields of motor cortical tDCS. Brain Stimul. 2015, 8, 906–913. [Google Scholar] [CrossRef]
- Tatti, E.; Rossi, S.; Innocenti, I.; Rossi, A.; Santarnecchi, E. Non-invasive brain stimulation of the aging brain: State of the art and future perspectives. Ageing Res. Rev. 2016, 29, 66–89. [Google Scholar] [CrossRef]
- Mahdavi, S.; Towhidkhah, F. Alzheimer’s Disease Neuroimaging Initiative. Computational human head models of tDCS: Influence of brain atrophy on current density distribution. Brain Stimul. 2018, 11, 104–107. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef]
- Stagg, C.J.; Nitsche, M.A. Physiological basis of transcranial direct current stimulation. Neuroscientist 2011, 17, 37–53. [Google Scholar] [CrossRef]
- Thomas, C.; Datta, A.; Woods, A. Effect of aging on current flow due to transcranial direct current stimulation. Brain Stimul. 2017, 10, 469. [Google Scholar] [CrossRef]
- Fujiyama, H.; Hyde, J.; Hinder, M.; Kim, S.-J.; McCormack, G.H.; Vickers, J.C.; Summers, J.J. Delayed plastic responses to anodal tDCS in older adults. Front. Aging Neurosci. 2014, 6, 115. [Google Scholar] [CrossRef] [PubMed]
- McLaren, M.E.; Nissim, N.R.; Woods, A.J. The effects of medication use in transcranial direct current stimulation: A brief review. Brain Stimul. 2018, 11, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H.; Bédirian, V. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriat. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.; Creelman, C.D. PEST: Efficient estimates on probability functions. J. Acoust. Soc. Am. 1967, 41, 782–787. [Google Scholar] [CrossRef]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef]
- Filmer, H.L.; Dux, P.E.; Mattingley, J.B. Applications of transcranial direct current stimulation for understanding brain function. Trends Neurosci. 2014, 37, 742–753. [Google Scholar] [CrossRef]
- Bestmann, S.; de Berker, A.O.; Bonaiuto, J. Understanding the behavioural consequences of noninvasive brain stimulation. Trends Cogn. Sci. 2015, 19, 13–20. [Google Scholar] [CrossRef]
- Shekhawat, G.S.; Stinear, C.M.; Searchfield, G.D. Transcranial direct current stimulation intensity and duration effects on tinnitus suppression. Neurorehabil. Neural Repair 2013, 27, 164–172. [Google Scholar] [CrossRef]
- Shekhawat, G.S.; Sundram, F.; Bikson, M.; Truong, D.Q.; De Ridder, D.; Stinear, C.; Welch, D.; Searchfield, G. Intensity, duration, and location of high-definition transcranial direct current stimulation for tinnitus relief. Neurorehabil. Neural Repair 2016, 30, 349–359. [Google Scholar] [CrossRef]
- Summers, J.J.; Kang, N.; Cauraugh, J.H. Does transcranial direct current stimulation enhance cognitive and motor functions in the ageing brain? A systematic review and meta-analysis. Ageing Res. Rev. 2016, 25, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Antal, A.; Kincses, T.Z.; Nitsche, M.A.; Bartfai, O.; Paulus, W. Excitability changes induced in the human primary visual cortex by transcranial direct current stimulation: Direct electrophysiological evidence. Invest. Ophthal. Vis. Sci. 2004, 45, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Antal, A.; Paulus, W. Transcranial direct current stimulation and visual perception. Perception 2008, 37, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, S.; Larochelle-Brunet, F.; Lafleur, L.P.; El Mouderrib, S.; Lepage, J.F.; Théoret, H. Systematic assessment of duration and intensity of anodal transcranial direct current stimulation on primary motor cortex excitability. Eur. J. Neurosci. 2016, 44, 2184–2190. [Google Scholar] [CrossRef]
- Horvath, J.C.; Carter, O.; Forte, J.D. Transcranial direct current stimulation: Five important issues we aren’t discussing (but probably should be). Front. Syst. Neurosci. 2014, 8, 2. [Google Scholar] [CrossRef]
- Monte-Silva, K.; Kuo, M.-F.; Hessenthaler, S.; Fresnoza, S.; Liebetanz, D.; Paulus, W.; Nitsche, M.A. Induction of late LTP-like plasticity in the human motor cortex by repeated non-invasive brain stimulation. Brain Stimul. 2013, 6, 424–432. [Google Scholar] [CrossRef]
- Krause, B.; Márquez-Ruiz, J.; Cohen Kadosh, R. The effect of transcranial direct current stimulation: A role for cortical excitation/inhibition balance? Front. Human Neurosci. 2013, 7, 602. [Google Scholar] [CrossRef]
- Kidgell, D.; Daly, R.M.; Young, K.; Lum, J.; Tooley, G.; Jaberzadeh, S.; Zoghi, M.; Pearce, A.J. Different current intensities of anodal transcranial direct current stimulation do not differentially modulate motor cortex plasticity. Neural Plast. 2013, 2013, 603502. [Google Scholar] [CrossRef]
- Bastani, A.; Jaberzadeh, S. Differential Modulation of Corticospinal Excitability by Different Current Densities of Anodal Transcranial Direct Current Stimulation. PLoS ONE 2013, 8, e72254. [Google Scholar] [CrossRef]
- Lisman, J.E. Three Ca2+ levels affect plasticity differently: The LTP zone, the LTD zone and no man’s land. J. Physiol. 2001, 532, 285. [Google Scholar] [CrossRef]
- Opitz, A.; Paulus, W.; Will, S.; Antunes, A.; Thielscher, A. Determinants of the electric field during transcranial direct current stimulation. NeuroImage 2015, 109, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Bortoletto, M.; Pellicciari, M.C.; Rodella, C.; Miniussi, C. The interaction with task-induced activity is more important than polarization: A tDCS study. Brain Stimul. 2015, 8, 269–276. [Google Scholar] [CrossRef] [PubMed]
- López-Alonso, V.; Fernández-del-Olmo, M.; Costantini, A.; Gonzalez-Henriquez, J.J.; Cheeran, B. Intra-individual variability in the response to anodal transcranial direct current stimulation. Clin. Neurophysiol. 2015, 126, 2342–2347. [Google Scholar] [CrossRef] [PubMed]
- Wiethoff, S.; Hamada, M.; Rothwell, J.C. Variability in response to transcranial direct current stimulation of the motor cortex. Brain Stimul. 2014, 7, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Puri, R.; Hinder, M.R.; Fujiyama, H.; Gomez, R.; Carson, R.G.; Summers, J.J. Duration-dependent effects of the BDNF Val66Met polymorphism on anodal tDCS induced motor cortex plasticity in older adults: A group and individual perspective. Front. Aging Neurosci. 2015, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, B.; Reis, J.; Martinowich, K.; Schambra, H.M.; Ji, Y.; Cohen, L.G.; Lu, B. Direct current stimulation promotes BDNF-dependent synaptic plasticity: Potential implications for motor learning. Neuron 2010, 66, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Nathan, P.J.; Cobb, S.R.; Lu, B.; Bullmore, E.T.; Davies, C.H. Studying synaptic plasticity in the human brain and opportunities for drug discovery. Curr. Opin. Pharmacol. 2011, 11, 540–548. [Google Scholar] [CrossRef]
- Fathi, D.; Ueki, Y.; Mima, T.; Koganemaru, S.; Nagamine, T.; Tawfik, A.; Fukuyama, H. Effects of aging on the human motor cortical plasticity studied by paired associative stimulation. Clin. Neurophysiol. 2010, 121, 90–93. [Google Scholar] [CrossRef]
- Cabeza, R. Hemispheric asymmetry reduction in older adults: The HAROLD model. Psychol. Aging 2002, 17, 85. [Google Scholar] [CrossRef]
- Reuter-Lorenz, P.A.; Park, D.C. How does it STAC up? Revisiting the scaffolding theory of aging and cognition. Neuropsychol. Rev. 2014, 24, 355–370. [Google Scholar] [CrossRef]
- Cabeza, R.; Albert, M.; Belleville, S.; Craik, F.I.M.; Duarte, A.; Grady, C.L.; Lindenberger, U.; Nyberg, L.; Park, D.C.; Reuter-Lorenz, P.A.; et al. Maintenance, reserve and compensation: The cognitive neuroscience of healthy ageing. Nat. Rev. Neurosci. 2018, 19, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Seider, T.R.; Fieo, R.A.; O’Shea, A.; Porges, E.C.; Woods, A.J.; Cohen, R.A. Cognitively engaging activity is associated with greater cortical and subcortical volumes. Front. Aging Neurosci. 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Matura, S.; Fleckenstein, J.; Deichmann, R.; Engeroff, T.; Füzéki, E.; Hattingen, E.; Hellweg, R.; Lienerth, B.; Pilatus, U.; Schwarz, S.; et al. Effects of aerobic exercise on brain metabolism and grey matter volume in older adults: Results of the randomised controlled SMART trial. Transl. Psychiatry 2017, 7, e1172. [Google Scholar] [CrossRef] [PubMed]
- Indahlastari, A.; Albizu, A.; O’Shea, A.; Forbes, M.A.; Nissim, N.R.; Kraft, J.N.; Evangelista, N.D.; Hausman, H.K.; Woods, A.J.; Initiative, A.D.N. Modeling transcranial electrical stimulation in the aging brain. Brain Stimul. 2020, 13, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Bikson, M.; Rahman, A.; Datta, A. Computational models of transcranial direct current stimulation. Clin. EEG Neurosci. 2012, 43, 176–183. [Google Scholar] [CrossRef]
- Rampersad, S.M.; Janssen, A.; Lucka, F.; Aydin, U.; Lanfer, B.; Lew, S.; Wolters, C.; Stegeman, D.F.; Oostendorp, T.F. Simulating transcranial direct current stimulation with a detailed anisotropic human head model. IEEE Trans. Neural Syst. Rehabil. 2014, 22, 441–452. [Google Scholar] [CrossRef]
- Mikkonen, M.; Laakso, I.; Tanaka, S.; Hirata, A. Cost of focality in TDCS: Interindividual variability in electric fields. Brain Stimul. 2020, 13, 117–124. [Google Scholar] [CrossRef]
- Laakso, I.; Mikkonen, M.; Koyama, S.; Hirata, A.; Tanaka, S. Can electric fields explain inter-individual variability in transcranial direct current stimulation of the motor cortex? Sci. Rep. 2019, 9, 626. [Google Scholar] [CrossRef]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef]
- Hsu, W.Y.; Ku, Y.; Zanto, T.P.; Gazzaley, A. Effects of noninvasive brain stimulation on cognitive function in healthy aging and Alzheimer’s disease: A systematic review and meta-analysis. Neurobiol. Aging 2015, 36, 2348–2359. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Vanderhasselt, M.A. Working memory improvement with non-invasive brain stimulation of the dorsolateral prefrontal cortex: A systematic review and meta-analysis. Brain Cogn. 2014, 86, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.T.; Stephens, J.A.; Alam, M.; Bikson, M.; Berryhill, M.E. Longitudinal neurostimulation in older adults improves working memory. PLoS ONE 2015, 10, e0121904. [Google Scholar] [CrossRef] [PubMed]
- E O’Connell, N.; Cossar, J.; Marston, L.; Wand, B.M.; Bunce, D.; Moseley, G.L.; De Souza, L.H. Rethinking Clinical Trials of Transcranial Direct Current Stimulation: Participant and Assessor Blinding Is Inadequate at Intensities of 2mA. PLoS ONE 2012, 7, e47514. [Google Scholar] [CrossRef] [PubMed]
- Fonteneau, C.; Mondino, M.; Arns, M.; Baeken, C.; Bikson, M.; Brunoni, A.R.; Poulet, E. Sham tDCS: A hidden source of variability? Reflections for further blinded, controlled trials. Brain Stimul. 2019, 12, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Goodwill, A.M.; Reynolds, J.; Daly, R.M.; Kidgell, D.J. Formation of cortical plasticity in older adults following tDCS and motor training. Front. Aging Neurosci. 2013, 5, 87. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanley, C.J.; Alderman, S.L.; Clemence, E. Optimising Cognitive Enhancement: Systematic Assessment of the Effects of tDCS Duration in Older Adults. Brain Sci. 2020, 10, 304. https://doi.org/10.3390/brainsci10050304
Hanley CJ, Alderman SL, Clemence E. Optimising Cognitive Enhancement: Systematic Assessment of the Effects of tDCS Duration in Older Adults. Brain Sciences. 2020; 10(5):304. https://doi.org/10.3390/brainsci10050304
Chicago/Turabian StyleHanley, Claire J., Sophie L. Alderman, and Elinor Clemence. 2020. "Optimising Cognitive Enhancement: Systematic Assessment of the Effects of tDCS Duration in Older Adults" Brain Sciences 10, no. 5: 304. https://doi.org/10.3390/brainsci10050304
APA StyleHanley, C. J., Alderman, S. L., & Clemence, E. (2020). Optimising Cognitive Enhancement: Systematic Assessment of the Effects of tDCS Duration in Older Adults. Brain Sciences, 10(5), 304. https://doi.org/10.3390/brainsci10050304