Enhancing Memory for Relationship Actions by Transcranial Direct Current Stimulation of the Superior Temporal Sulcus

 ,
,  ,
,
Abstract
:1. Introduction
Moderation of tDCS Effect by Approach and Avoidance Personality Traits
2. Study 1—Memorization of Approach/Avoidance Sentences
2.1. Method
2.1.1. Participants
2.1.2. Stimuli
2.1.3. Design
2.1.4. Procedure
2.2. Results
3. Study 2—tDCS Effects on Memorization of Relationship Action-Sentences
3.1. Method
3.1.1. Participants
3.1.2. Approach and Avoidance Behavioral Scales (BIS/BAS)
3.1.3. Design
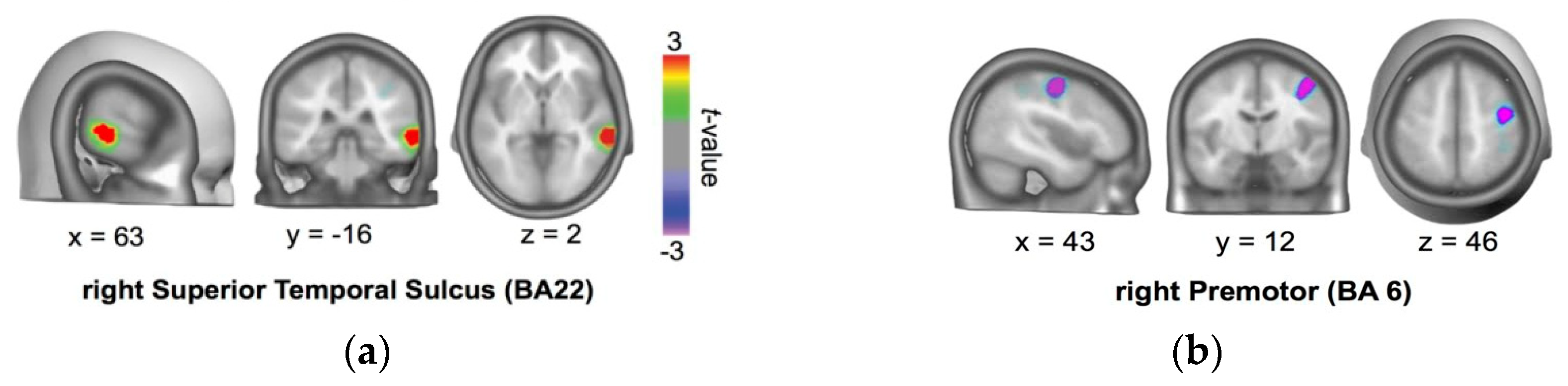
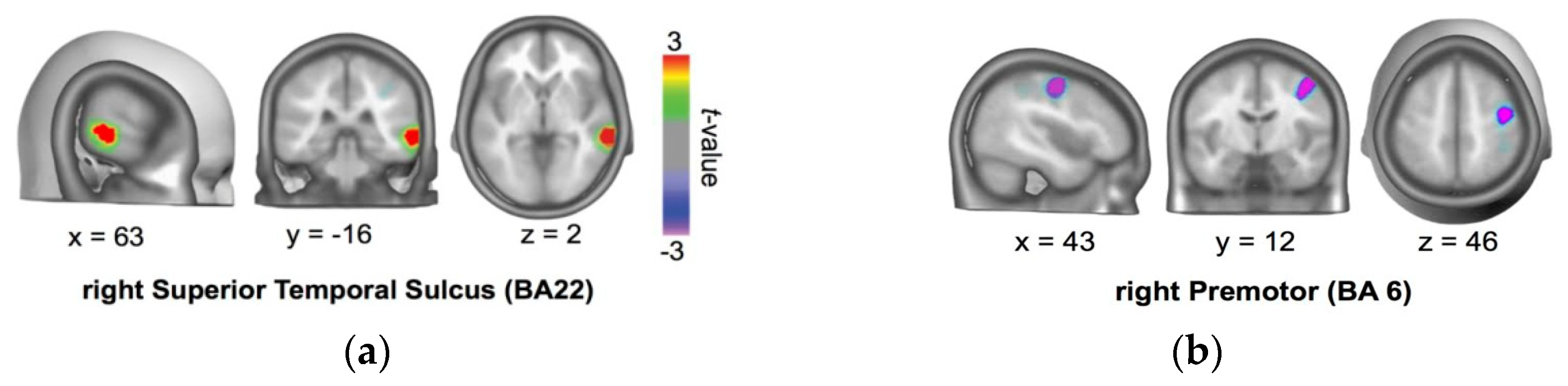
3.1.4. Protocol for tDCS Application
3.1.5. tDCS Procedure
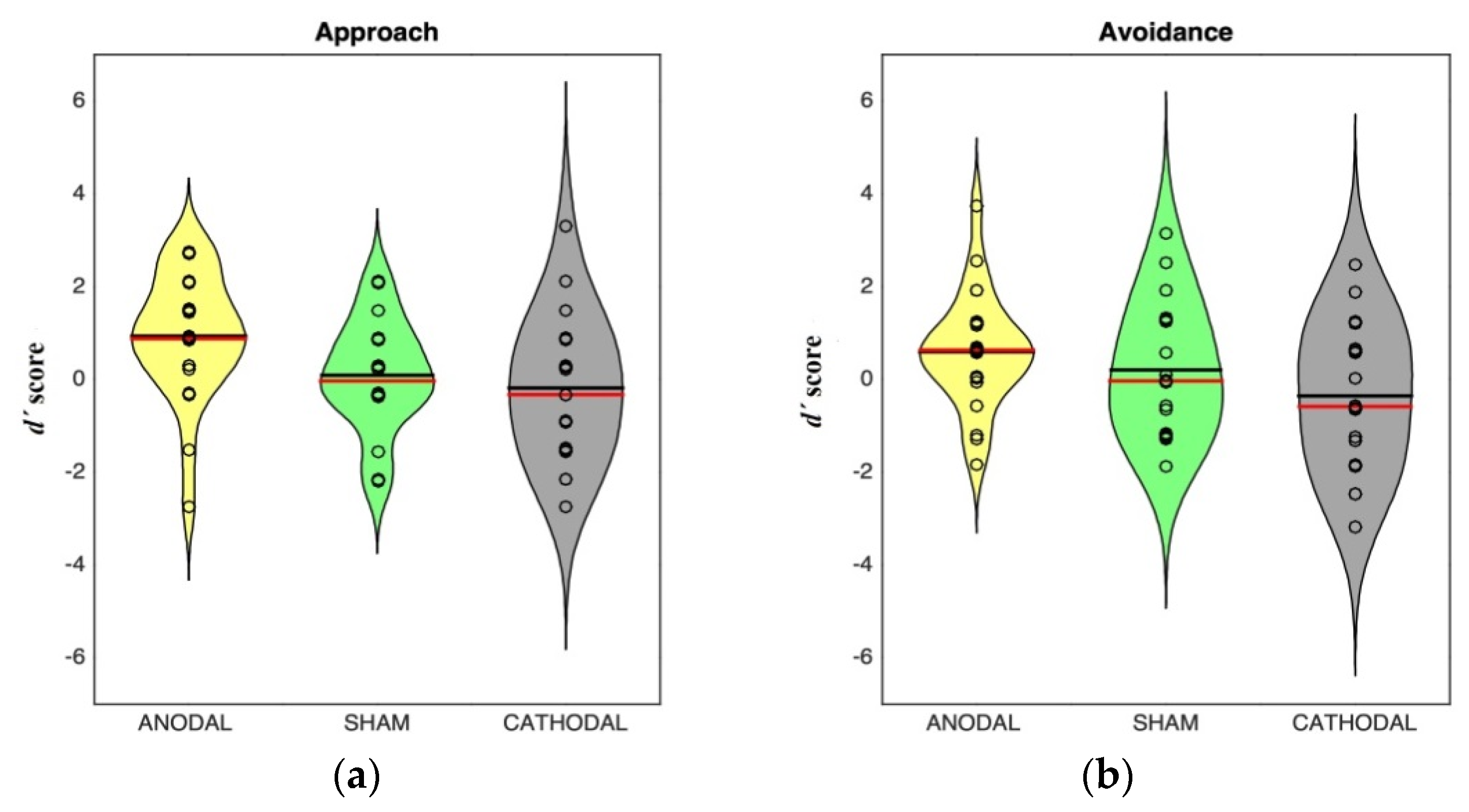
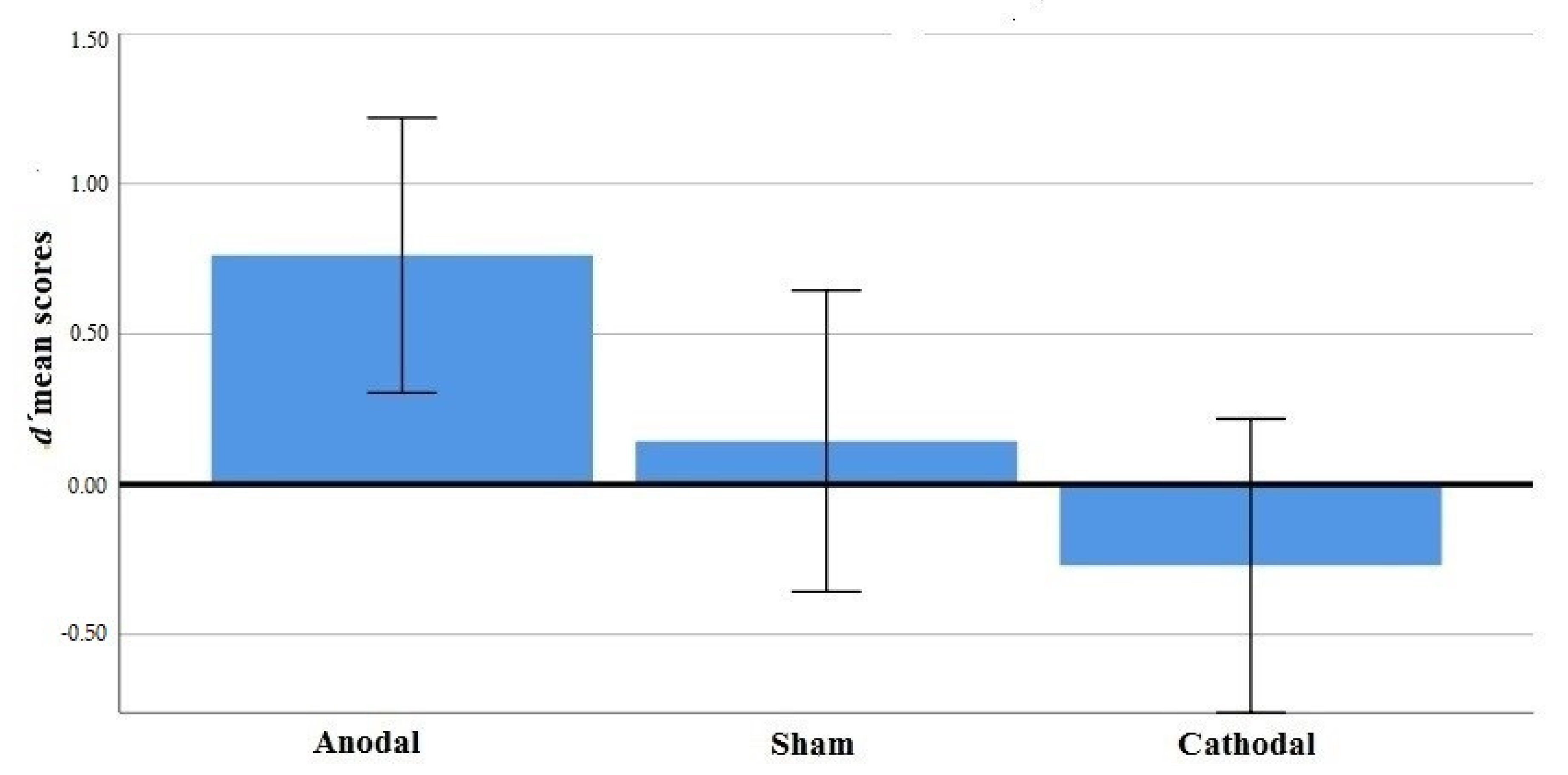
3.2. Results
Moderation of tDCS Effect by Affective Traits
4. General Discussion
4.1. tDCS Effect on Memorization is Modulated by Approach/Avoidance Trait
4.2. Limitations, Contributions and Future Directions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dodell-Feder, D.; Koster-Hale, J.; Bedny, M.; Saxe, R. fMRI item analysis in a theory of mind task. Neuroimage 2011, 55, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.P.; Adolphs, R. The social brain in psychiatric and neurological disorders. Trends Cogn. Sci. 2012, 16, 559–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spunt, R.P.; Falk, E.B.; Lieberman, M.D. Dissociable neural systems support retrieval of how and why action knowledge. Psychol. Sci. 2010, 21, 1593–1598. [Google Scholar] [CrossRef]
- Gobbini, M.I.; Koralek, A.C.; Bryan, R.E.; Montgomery, K.J.; Haxby, J.V. Two takes on the social brain: A comparison of theory of mind tasks. J. Cogn. Neurosci. 2007, 19, 1803–1814. [Google Scholar] [CrossRef]
- Watson, R.; Latinus, M.; Charest, I.; Crabbe, F.; Belin, P. People-selectivity, audiovisual integration and heteromodality in the superior temporal sulcus. Cortex 2014, 50, 125–136. [Google Scholar] [CrossRef]
- Wong, C.; Gallate, J. The function of the anterior temporal lobe: A review of the empirical evidence. Brain Res. 2012, 1449, 94–116. [Google Scholar] [CrossRef]
- Johnson, M.H.; Senju, A.; Tomalski, P. The two-process theory of face processing: Modifications based on two decades of data from infants and adults. Neurosci. Biobehav. Rev. 2015, 50C, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Pelphrey, K.A.; Morris, J.P. Brain mechanisms for interpreting the actions of others from biological motion cues. Curr. Dir. Psychol. Sci. 2006, 15, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Pelphrey, K.A.; Carter, E.J. Brain mechanisms for social perception: Lessons from autism and typical development. Ann. N. Y. Acad. Sci. 2008, 1145, 283–299. [Google Scholar] [CrossRef] [Green Version]
- Saitovitch, A.; Bargiacchi, A.; Chabane, N.; Brunelle, F.; Samson, Y.; Boddaert, N.; Zilbovicius, M. Social cognition and the superior temporal sulcus: Implications in autism. Rev. Neurol. 2012, 168, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.Y.; Rosenblau, G.; Keifer, C.; Pelphrey, K.A. An integrative neural model of social perception, action observation, and theory of mind. Neurosci. Biobehav. Rev. 2015, 51, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, L.E.; Eckstrand, K.L.; Silk, J.S.; Allen, N.B.; Ambrosia, M.; Healey, K.L.; Forbes, E.E. Adolescents’ neural response to social reward and real-world emotional closeness and positive affect. Cogn. Affect. Behav. Neurosci. 2018, 18, 705–717. [Google Scholar] [CrossRef]
- Pelphrey, K.A.; Viola, R.J.; McCarthy, G. When strangers pass. Psychol. Sci. 2004, 15, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Ross, L.A.; Olson, I.R. Social cognition and the anterior temporal lobes. NeuroImage 2004, 49, 3452–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, P.; Lawrence, A.D.; Barnard, P.J. Paying attention to social meaning: An FMRI study. Cereb. Cortex 2008, 18, 1876–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacoboni, M.; Lieberman, M.D.; Knowlton, B.J.; Molnar-Szakacs, I.; Moritz, M.; Throop, C.; Fiske, A.P. Watching social interactions produces dorsomedial prefrontal and medial parietal BOLD fMRI signal increases compared to a resting baseline. NeuroImage 2004, 21, 1167–1173. [Google Scholar] [CrossRef]
- Lahnakoski, J.M.; Glerean, E.; Salmi, J.; Jääskeläinen, I.P.; Sams, M.; Hari, R.; Nummenmaa, L. Naturalistic FMRI mapping reveals superior temporal sulcus as the hub for the distributed brain network for social perception. Front. Hum. Neurosci. 2012, 6, 233. [Google Scholar] [CrossRef] [Green Version]
- Frith, U.; Frith, C.D. Development and neurophysiology of mentalizing. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Zahn, R.; Moll, J.; Krueger, F.; Huey, E.D.; Garrido, G.; Grafman, J. Social concepts are represented in the superior anterior temporal cortex. Proc. Natl. Acad. Sci. USA 2007, 104, 6430–6435. [Google Scholar] [CrossRef] [Green Version]
- Lambon Ralph, M.A.; Jefferies, E.; Patterson, K.; Rogers, T.T. The neural and computational bases of semantic cognition. Nat. Rev. Neurosci. 2017, 18, 42–55. [Google Scholar] [CrossRef]
- Marrero, H.; Urrutia, M.; Beltrán, D.; Gámez, E.; Díaz, J.M. Understanding approach and avoidance in verbal descriptions of everyday actions: An ERP study. Cogn. Affect. Behav. Neurosci. 2017, 17, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Marrero, H.; Gámez, E.; Diaz, J.M.; Urrutia, M.; de Vega, M. Carefully encoding approach and avoidance body locomotion with interpersonal conduct in narrated interactions. Can. J. Exp. Psychol. 2015, 69, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Marrero, H.; Gámez, E.; Urrutia, M.; Beltrán, D.; Diaz, J.M.; Yagual, S.N. Brain Encoding of Social Approach: Is it Associated with Spatial Ability? Front. Behav. Neurosci. 2019, 13, 179. [Google Scholar] [CrossRef] [Green Version]
- Grave de Peralta, R.; Andino, S.; Lantz, G.; Michel, C.M.; Landis, T. Noninvasive localization of electromagnetic epileptic activity. I. Method descriptions and simulations. Brain Topogr. 2001, 14, 131–137. [Google Scholar] [CrossRef]
- Logan, G.D. An instance theory of attention and memory. Psychol. Rev. 2002, 109, 376–400. [Google Scholar] [CrossRef] [PubMed]
- Macmillan, N.A.; Kaplan, H.L. Detection theory analysis of group data: Estimating sensitivity from average hit and false-alarm rates. Psychol. Bull. 1985, 98, 185–199. [Google Scholar] [CrossRef]
- Gray, J.A. A critique of Eysenck’s theory of personality. In A Model for Personality; Eysenck, H.J., Ed.; Springer-Verlag: Berlin, Germany, 1981; pp. 246–276. [Google Scholar]
- Metuki, N.; Sela, T.; Lavidor, M. Enhancing cognitive control components of insight problem solving by anodal tDCS of the left dorsolateral prefrontal cortex. Brain Stimul. 2012, 5, 110–115. [Google Scholar] [CrossRef]
- Sela, T.; Ivry, R.B.; Lavidor, M. Prefrontal control during a semantic decision task that involves idiom comprehension: A transcranial direct current stimulation study. Neuropsychologia 2012, 50, 2271–2280. [Google Scholar] [CrossRef]
- Blumenthal, T.D. Extraversion, attention, and startle response reactivity. PAID 2001, 30, 495–503. [Google Scholar] [CrossRef]
- Giakoumaki, S.G.; Roussos, P.; Tsapakis, E.M.; Koiliari, E.; Pasparakis, E.; Zouraraki, C.; Bitsios, P. Cognitive and personality analysis of startle reactivity in a large cohort of healthy males. Biol. Psychol. 2013, 94, 582–591. [Google Scholar] [CrossRef]
- LaRowe, S.D.; Patrick, C.J.; Curtin, J.J.; Kline, J.P. Personality correlates of startle habituation. Biol. Psychol. 2006, 72, 257–264. [Google Scholar] [CrossRef]
- Eysenck, M.W.; Derakshan, N.; Santos, R.; Calvo, M.G. Anxiety and cognitive performance: Attentional control theory. Emotion 2007, 7, 336–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, L.E.; Cacioppo, J.T. Learning where to look for danger: Integrating affective and spatial information. Psychol. Sci. 2002, 13, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.K.; Cacioppo, J.T.; Larsen, J.T.; Chartrand, T.L. May I have your attention, please: Electrocortical responses to positive and negative stimuli. Neuropsychologia 2003, 41, 171–183. [Google Scholar] [CrossRef]
- LaBar, K.S.; Phelps, E.A. Arousal-mediated memory consolidation: Role of the medial temporal lobe in humans. Psychol. Sci. 1998, 9, 490–493. [Google Scholar] [CrossRef]
- Sharot, T.; Delgado, M.R.; Phelps, E.A. How emotion enhances the feeling of remembering. Nat. Neurosci. 2004, 7, 1376–1380. [Google Scholar] [CrossRef]
- Díez, E.; Gómez-Ariza, C.J.; Díez-Álonso, A.; Alonso, M.A.; Fernandez, A. The processing of semantic relatedness in the brain: Evidence from associative and categorical false recognition effects following transcranial direct current stimulation of the left anterior temporal lobe. Cortex 2017, 93, 133–145. [Google Scholar] [CrossRef]
- Carver, C.S.; White, T.L. Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: The BIS/BAS scales. J. Personal. Soc. Psychol. 1994, 67, 319–333. [Google Scholar] [CrossRef]
- Jung, Y.J.; Kim, J.H.; Im, C.H. COMETS: A MATLAB toolbox for simulating local electric fields generated by transcranial direct current stimulation (tDCS). Biomed. Eng. Lett. 2013, 3, 39–46. [Google Scholar] [CrossRef]
- Zwissler, B.; Sperber, C.; Aigeldinger, S.; Schindler, S.; Kissler, J.; Plewnia, C. Shaping memory accuracy by left prefrontal transcranial direct current stimulation. J. Neurosci. 2014, 34, 4022e–4026e. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Paulus, W. Transcranial electrical stimulation (tES-tDCS; tRNS, tACS) methods. Neuropsychol. Rehabil. 2011, 21, 602–617. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.R.; Amadera, J.; Berbel, B.; Volz, M.S.; Rizzerio, B.G.; Fregni, F. A systematic review on reporting and assessment of adverse effects associated with transcranial direct current stimulation. Int. J. Neuropsychopharmacol. 2011, 14, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Marrero, H.; Gámez, E.; Díaz, J.M. BIS/BAS individual differences and the verification of conditional hypotheses. Pers. Individ. Differ. 2008, 45, 296–301. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichenbaum, H.; Yonelinas, A.P.; Ranganath, C. The medial temporal lobe and recognition memory. Annu. Rev. Neurosci. 2007, 30, 123–152. [Google Scholar] [CrossRef] [Green Version]
- Ranganath, C. A unified framework for the functional organization of the medial temporal lobes and the phenomenology of episodic memory. Hippocampus 2010, 20, 1263–1290. [Google Scholar] [CrossRef]
- Dedoncker, J.; Brunoni, A.R.; Baeken, C.; Vanderhasselt, M.A. A Systematic Review and Meta-Analysis of the Effects of Transcranial Direct Current Stimulation (tDCS) Over the Dorsolateral Prefrontal Cortex in Healthy and Neuropsychiatric Samples: Influence of Stimulation Parameters. Brain Stimul. 2016, 9, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Batsikadze, G.; Moliadze, V.; Paulus, W.; Kuo, M.F.; Nitsche, M.A. Partially non-linear stimulation intensity-dependent effects of direct current stimulation on motor cortex excitability in humans. J. Physiol. 2013, 591, 1987–2000. [Google Scholar] [CrossRef]
- Brasil-Neto, J.P. Learning, memory, and transcranial direct current stimulation. Front. Psych. 2012, 3, 80. [Google Scholar] [CrossRef] [Green Version]
- Boggio, P.S.; Fregni, F.; Valasek, C.; Ellwood, S.; Chi, R.; Gallate, J.; Pascual-Leone, A.; Snyder, A. Temporal lobe cortical electrical stimulation during the encoding and retrieval phase reduces false memories. PLoS ONE 2009, 4, e4959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Urbina, G.N.; Mendez Joya, M.F.; Nitsche, M.A.; Molero-Chamizo, A. Anodal tDCS over Wernicke’s area improves verbal memory and prevents the interference effect during words learning. Neuropsychology 2019, 33, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Sparing, R.; Dafotakis, M.; Meister, I.G.; Thirugnanasambandam, N.; Fink, G.R. Enhancing language performance with noninvasive brain stimulation—a transcranial direct current stimulation study in healthy humans. Neuropsychologia 2008, 46, 261–268. [Google Scholar] [CrossRef]
- Fan, I.; Mao, W.; Jin, J.; Ma, Q. Modulating activity in the dorsolateral prefrontal cortex alter corruption behavior: A transcranial direct current stimulation study. Behav. Brain Res. 2020, 382, 112479. [Google Scholar] [CrossRef] [PubMed]
- Pedroarena-Leal, N.; Heidemeyer, L.; Trenado, C.; Ruge, D. Human Depotentiation following Induction of Spike Timing Dependent Plasticity. Biomedicines 2018, 6, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkman, E.T.; Lieberman, M.D. Approaching the Bad and Avoiding the Good: Lateral Prefrontal Cortical Asymmetry Distinguishes between Action and Valence. J. Cogn. Neurosci. 2010, 22, 1970–1979. [Google Scholar] [CrossRef] [Green Version]
- Carver, C.S.; Harmon-Jones, E. Anger is an approach-related affect: Evidence and implications. Psychol. Bull. 2009, 135, 183–204. [Google Scholar] [CrossRef] [PubMed]
- Bonner, M.F.; Price, A.R. Where is the anterior temporal lobe and what does it do? J. Neurosci. 2013, 33, 4213–4215. [Google Scholar] [CrossRef] [Green Version]
- Saxe, R.; Kanwisher, N. People thinking about thinking people. The role of the temporo-parietal junction in “theory of mind”. NeuroImage 2003, 19, 1835–1842. [Google Scholar] [CrossRef]
- Cosmides, L. The logic of social exchange: Has natural selection shaped how humans reason? Studies with the Wason selection task. Cognition 1989, 31, 187–276. [Google Scholar] [CrossRef]
- Higgins, E.T. Promotion and prevention: Regulatory focus as a motivational principle. Adv. Exp. Soc. Psychol. 1998, 30, 1–45. [Google Scholar]
- Couto, B.; Manes, F.; Montañés, P.; Matallana, D.; Reyes, P.; Velázquez, M.; Yoris, A.; Baez, S.; Ibáñez, A. Structural neuroimaging of social cognition in progressive non-fluent aphasia and behavioural variant of frontotemporal dementia. Front. Hum. Neurosci. 2013, 7, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Memorization | Approach/Avoidance List | Same-Changed Judgement Task | Same/Changed Sentence |
|---|---|---|---|
| Pedro admitió a Rosa en el Whatsapp. (Pedro accepted Rosa in Whatsapp.) | Approach list 1 | Pedro admitió a Rosa en el Whatsapp. (Pedro accepted Rosa in Whatsapp.) | Same |
| Pedro bloqueó a Rosa en el Whatsapp. (Pedro blocked Rosa on Whatsapp.) | Avoidance list 2 | Rosa bloqueó a Pedro en el Whatsapp. (Rosa blocked Pedro onWhatsapp.) | Changed |
| Inés rechazó a Roberto por su papel. (Ines rejected Roberto for his role.) | Avoidance list 2 | Inés rechazó a Roberto por su papel. (Inés rejected Roberto for his role.) | Same |
| Inés elogió a Roberto por su papel. (Inés praised Roberto for his role.) | Approach list 1 | Roberto elogió a Inés por su papel. (Roberto praised Inés for his role.) | Changed |
| Type of Effect | Severity | Percentage |
|---|---|---|
| Tingling | Mild | 29.23% |
| Itching | Mild | 20.00% |
| Sleepiness | Mild | 12.30% |
| Sentence | Same | Changed | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Direction (tDCS) | Approach | Avoidance | Approach | Avoidance | N | ||||
| Anodal | 71 | (15.69) | 73 | (13.98) | 60 | (17.56) | 56 | (16.89) | 24 |
| Sham | 70 | (12.76) | 70 | (16.22) | 48 | (14.09) | 53 | (17.80) | 20 |
| Cathodal | 62 | (18.33) | 62 | (13.64) | 50 | (14.30) | 52 | (17.00) | 21 |
| Direction | Approach | Avoidance | Averaged |
|---|---|---|---|
| Anodal | 0.936 (1.34) | 0.587 (1.21) | 0.761 (0.985) |
| Sham | 0.087 (1.18) | 0.200 (1.40) | 0.144 (0.941) |
| Cathodal | −0.182 (1.52) | −0.358 (1.58) | −0.270 (1.391) |
| (a) Pearson Corr. tDCS with d’ | (b) ANODAL | |||||||
|---|---|---|---|---|---|---|---|---|
| BIS/BAS Trait | d’ Approach | d’ Avoidance | d’ Averaged | N | d’ Approach | d’ Avoidance | d’ Averaged | N |
| Low BAS | 0.551 (p = 0.002) | 0.341 (p = 0.065) | 0.537 (p = 0.002) | 30 | 1.45 (1.01) | 0.87 (1.29) | 1.16 (0.84) | 13 |
| High BAS | 0.096 (p > 0.20) | 0.136 (p > 0.20) | 0.132 (p > 0.20) | 34 | 0.32 (1.46) | 0.25 (1.07) | 0.28 (0.95) | 11 |
| Low BIS | 0.511 (p = 0.005) | 0.410 (p = 0.027) | 0.556 (p = 0.002) | 29 | 1.30 (1.21) | 0.64 (.94) | 0.97 (0.83) | 10 |
| High BIS | 0.163 (p > 0.20) | 0.123 (p > 0.20) | 0.169 (p > 0.20) | 35 | 0.67 (1.40) | 0.54 (1.41) | 0.60 (1.40) | 14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marrero, H.; Yagual, S.N.; García-Marco, E.; Gámez, E.; Beltrán, D.; Díaz, J.M.; Urrutia, M. Enhancing Memory for Relationship Actions by Transcranial Direct Current Stimulation of the Superior Temporal Sulcus. Brain Sci. 2020, 10, 497. https://doi.org/10.3390/brainsci10080497
Marrero H, Yagual SN, García-Marco E, Gámez E, Beltrán D, Díaz JM, Urrutia M. Enhancing Memory for Relationship Actions by Transcranial Direct Current Stimulation of the Superior Temporal Sulcus. Brain Sciences. 2020; 10(8):497. https://doi.org/10.3390/brainsci10080497
Chicago/Turabian StyleMarrero, Hipólito, Sara Nila Yagual, Enrique García-Marco, Elena Gámez, David Beltrán, Jose Miguel Díaz, and Mabel Urrutia. 2020. "Enhancing Memory for Relationship Actions by Transcranial Direct Current Stimulation of the Superior Temporal Sulcus" Brain Sciences 10, no. 8: 497. https://doi.org/10.3390/brainsci10080497
APA StyleMarrero, H., Yagual, S. N., García-Marco, E., Gámez, E., Beltrán, D., Díaz, J. M., & Urrutia, M. (2020). Enhancing Memory for Relationship Actions by Transcranial Direct Current Stimulation of the Superior Temporal Sulcus. Brain Sciences, 10(8), 497. https://doi.org/10.3390/brainsci10080497