Associations between Attention and Implicit Associative Learning in Healthy Adults: The Role of Cortisol and Salivary Alpha-Amylase Responses to an Acute Stressor


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
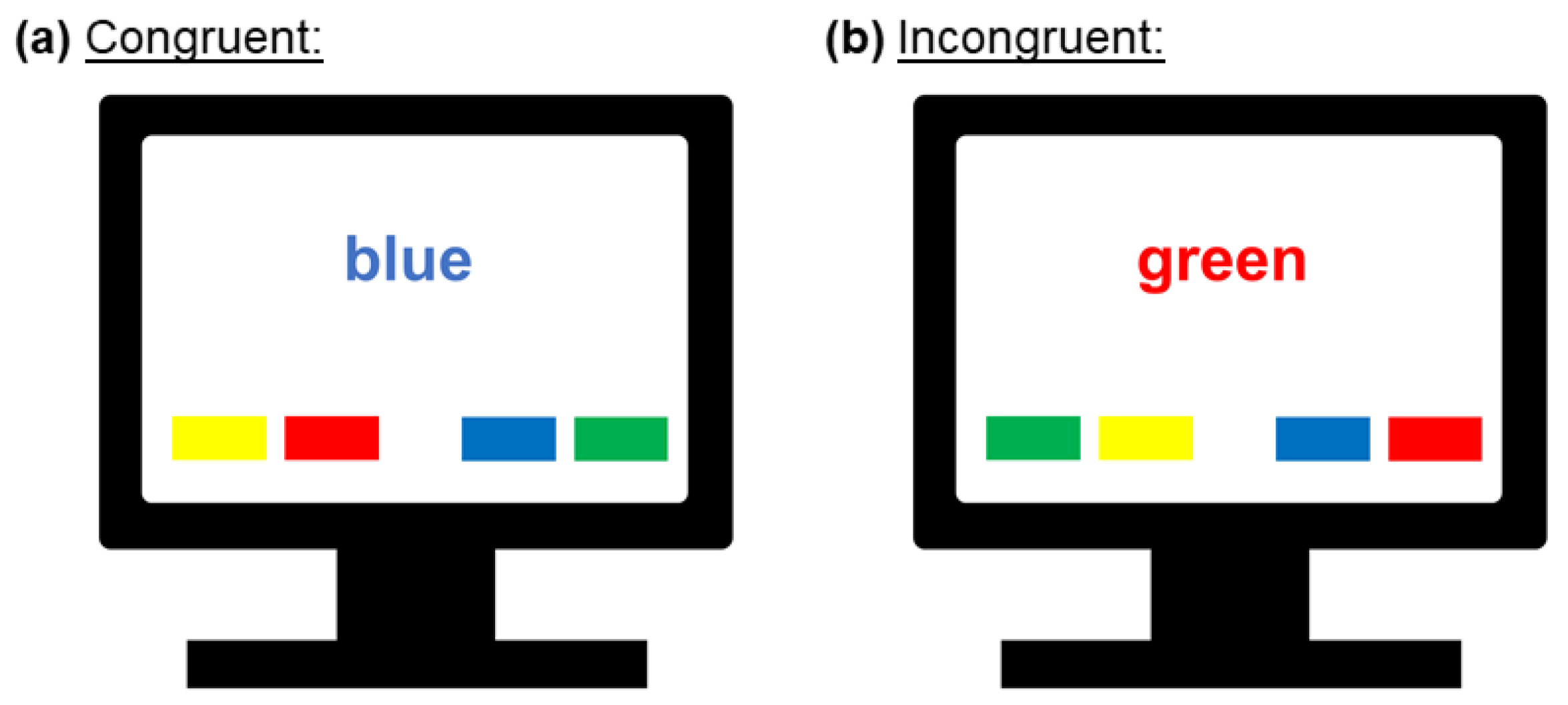
2.2. Cognitive Testing
2.3. Stress Induction and Control Condition
2.4. Saliva Sampling and Analysis
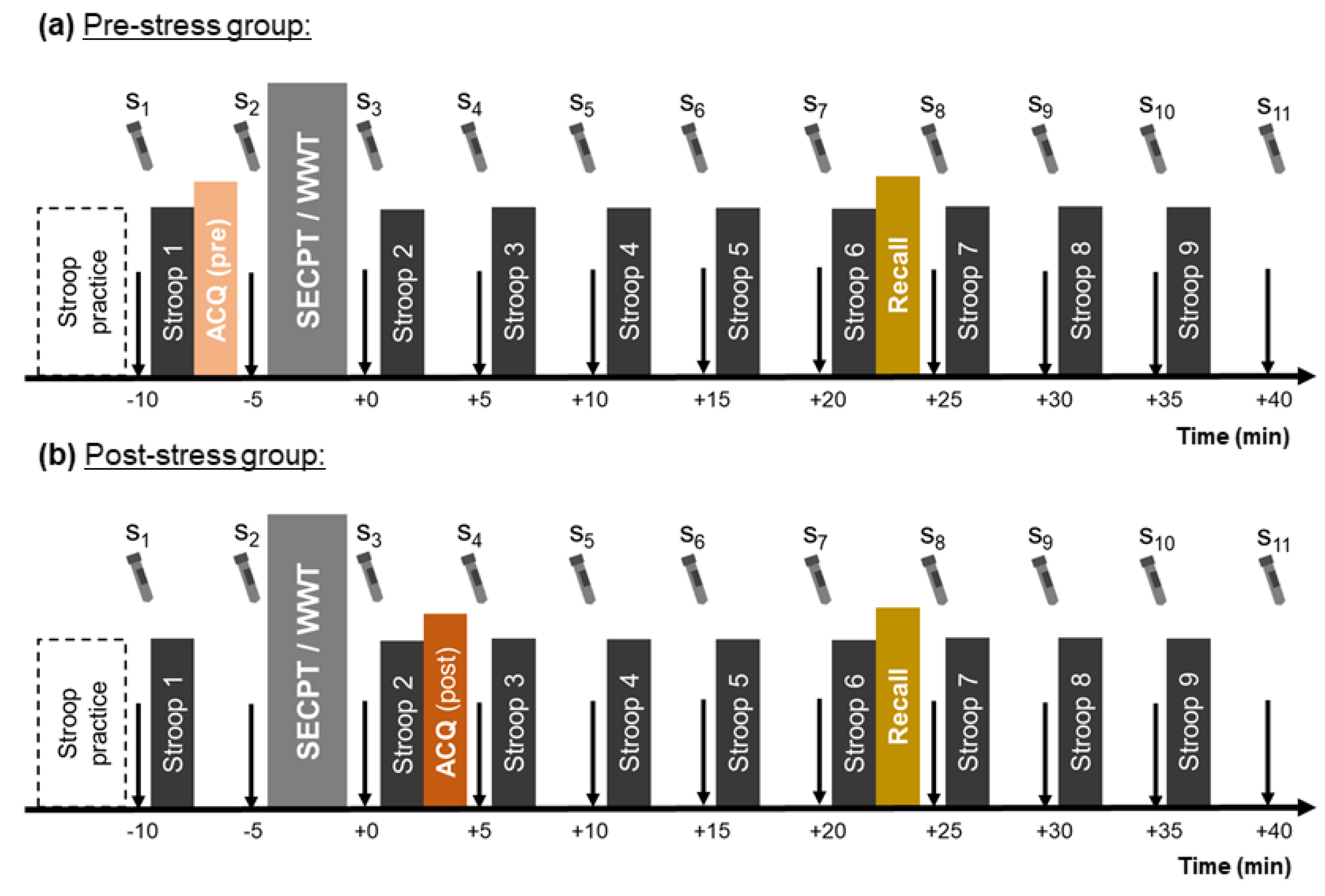
2.5. General Procedure
2.6. Statistical Data Analysis
3. Results
3.1. Baseline Characteristics
3.2. Stress Response
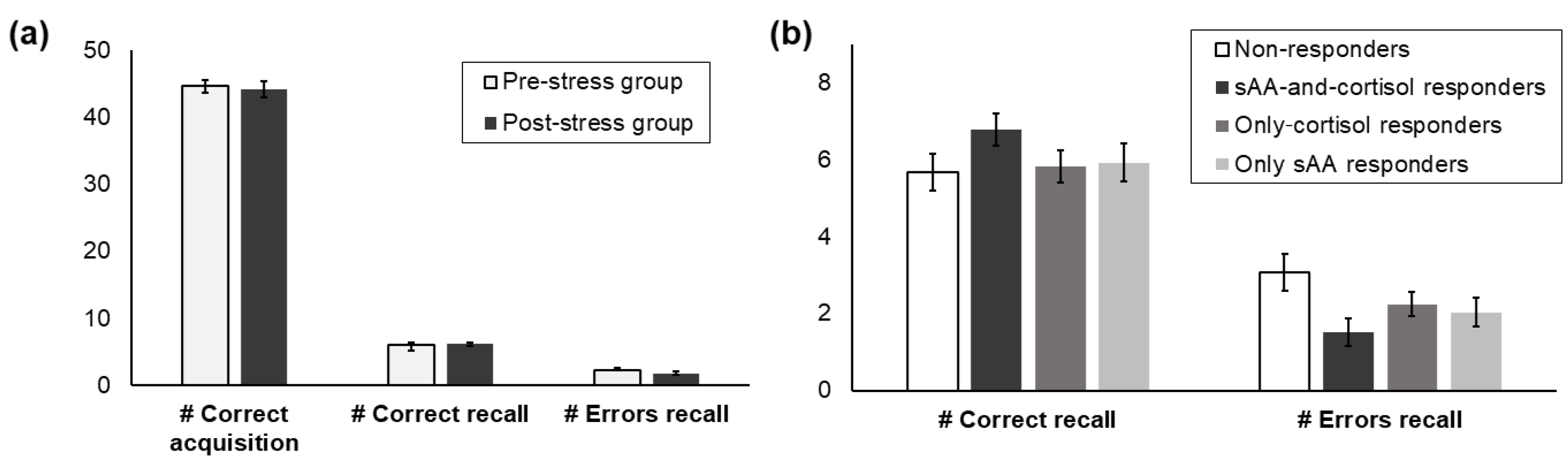
3.3. Memory Performance
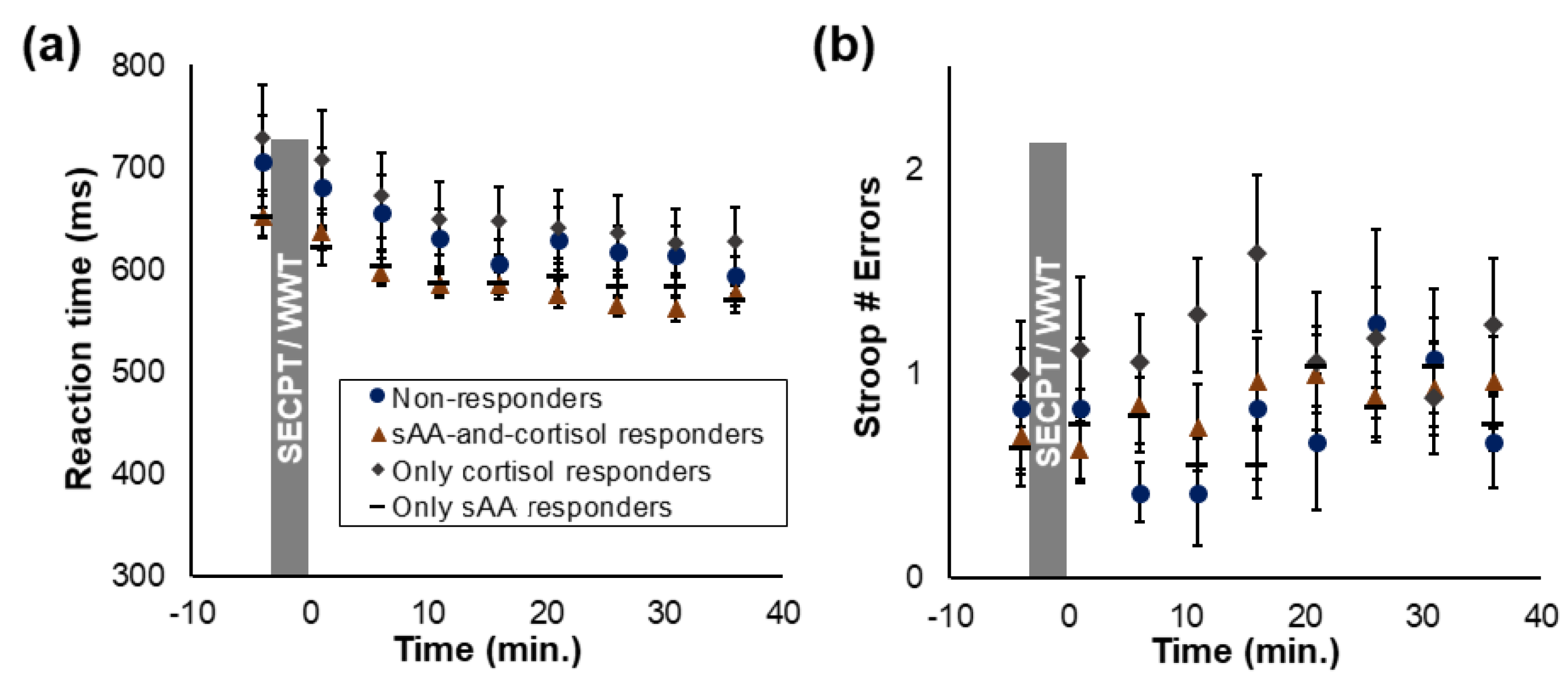
3.4. Attentional Time Course
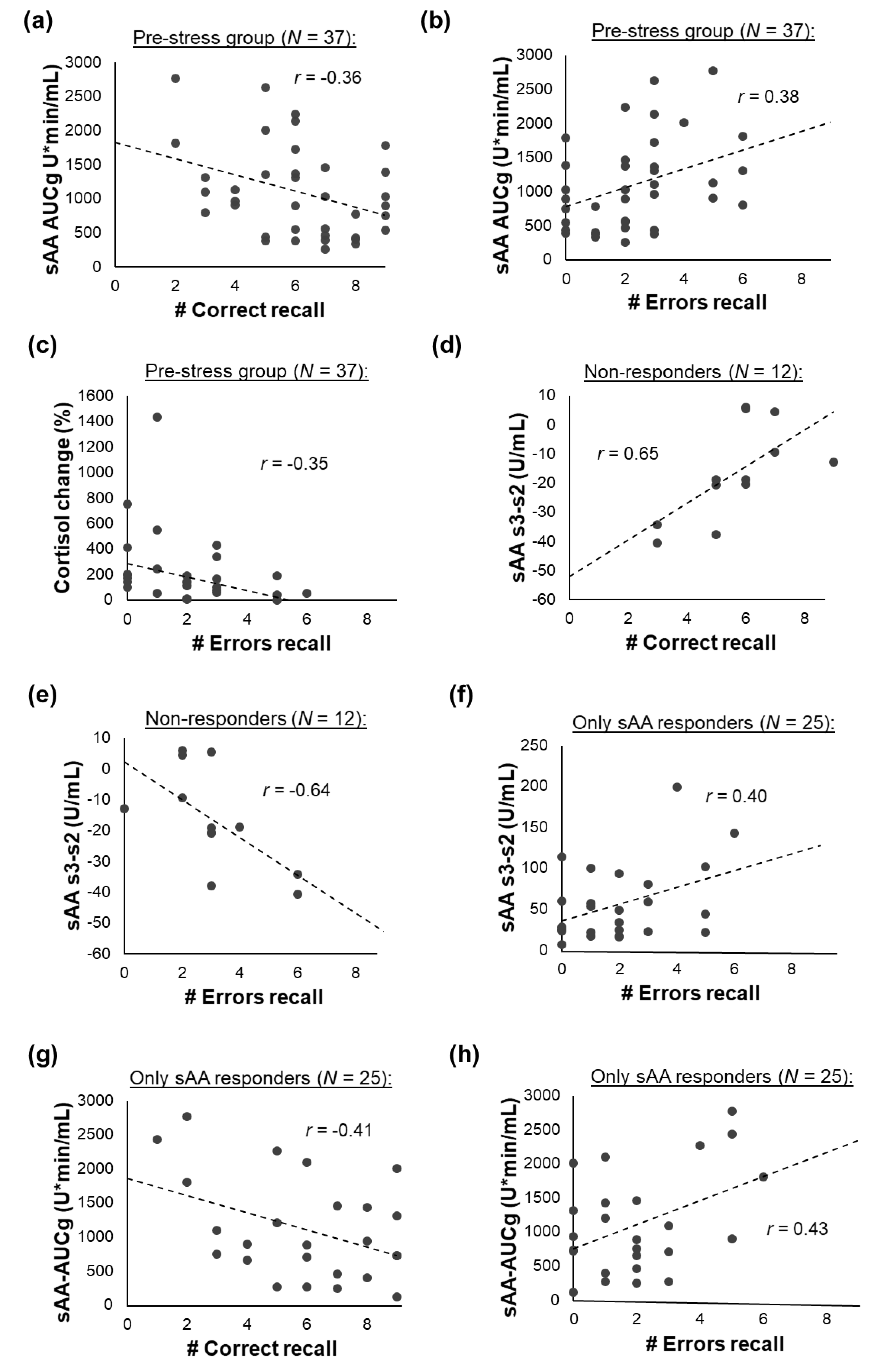
3.5. Associations between Memory Recall and the Stress Response
3.5.1. All Participants
3.5.2. Pre- vs. Post-Stress Group
3.5.3. Responder Groups
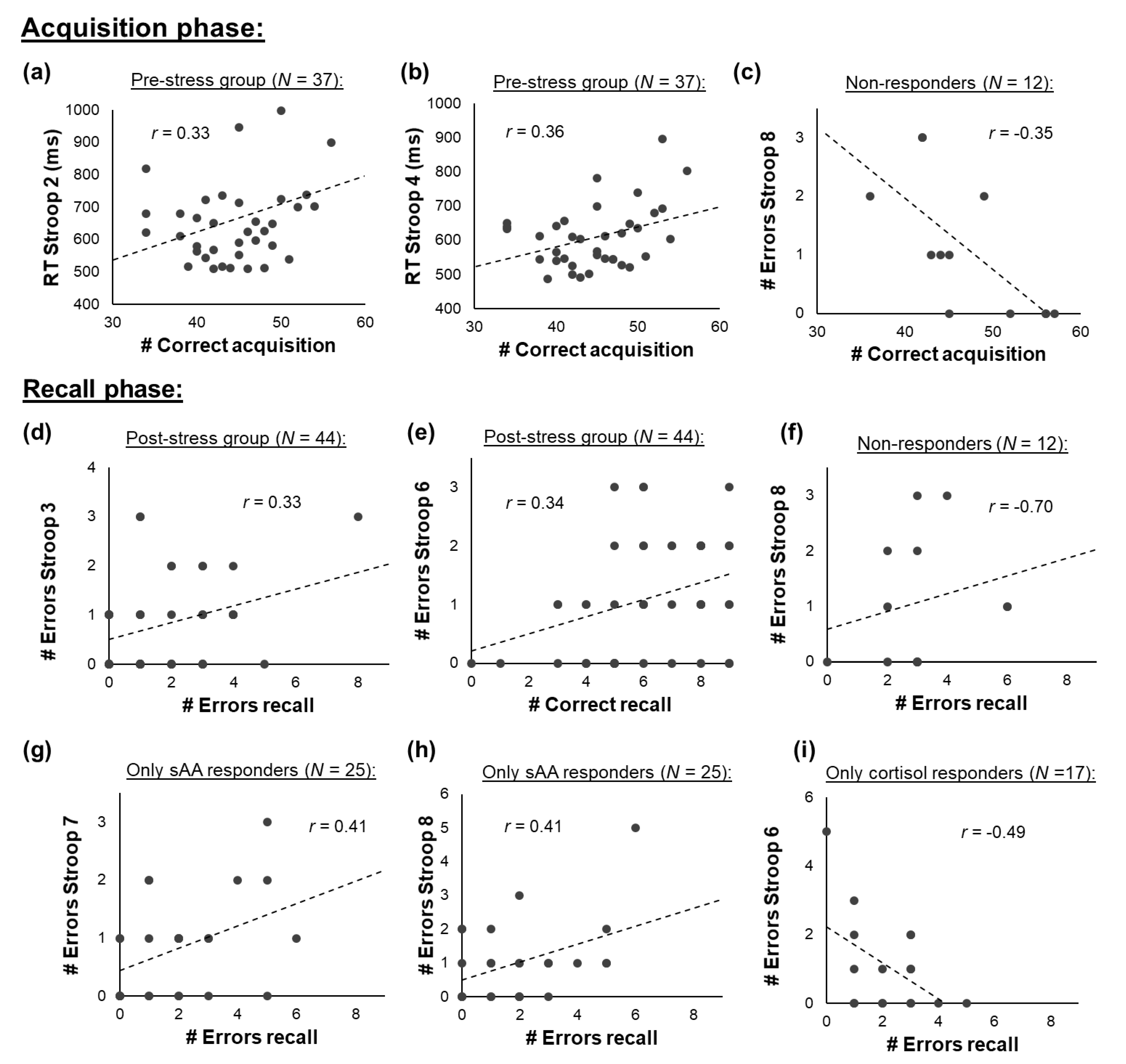
3.6. Associations between Attention and Implicit Learning
3.6.1. All Participants
3.6.2. Pre- vs. Post-Stress Group
3.6.3. Responder Groups
4. Discussion
4.1. Summary and Discussion of Main Findings
4.2. Summary and Discussion of Further Findings
4.3. Physiological Responses to the Stress and the Control Task
4.4. Limitations and Directions for Future Research
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Squire, L.R.; Zola-Morgan, S. Memory: Brain systems and behavior. Trends Neurosci. 1988, 11, 170–175. [Google Scholar] [CrossRef]
- Henke, K. A model for memory systems based on processing modes rather than consciousness. Nat. Rev. Neurosci. 2010, 11, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Henke, K.; Buck, A.; Weber, B.; Wieser, H.G. Human hippocampus establishes associations in memory. Hippocampus 1997, 7, 249–256. [Google Scholar] [CrossRef]
- Preston, A.R.; Gabrieli, J.D.E. Dissociation between explicit memory and configural memory in the human medial temporal lobe. Cereb. Cortex 2008, 18, 2192–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schott, B.H.; Sellner, D.B.; Lauer, C.-J.; Habib, R.; Frey, J.U.; Guderian, S.; Heinze, H.-J.; Düzel, E. Activation of midbrain structures by associative novelty and the formation of explicit memory in humans. Learn. Mem. 2004, 11, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Luethi, M.; Meier, B.; Sandi, C. Stress effects on working memory, explicit memory, and implicit memory for neutral and emotional stimuli in healthy men. Front. Behav. Neurosci. 2008, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, M.; Todd, R. Acute psychophysiological stress impairs human associative learning. Neurobiol. Learn. Mem. 2017, 145, 84–93. [Google Scholar] [CrossRef]
- Meyer, T.; Smeets, T.; Giesbrecht, T.; Quaedflieg, C.W.E.M.; Merckelbach, H. Acute stress differentially affects spatial configuration learning in high and low cortisol-responding healthy adults. Eur. J. Psychotraumatology 2013, 4, 19854. [Google Scholar] [CrossRef]
- Chrousos, G.P. The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. JAMA 1992, 267, 1244–1252. [Google Scholar] [CrossRef]
- Stratakis, C.A.; Chrousos, G.P. Neuroendocrinology and pathophysiology of the stress system. Ann. N. Y. Acad. Sci. 1995, 771, 1–18. [Google Scholar] [CrossRef]
- Arnsten, A.F.T. Stress signalling pathways that impair prefrontal cortex structure and function. Nat. Rev. Neurosci. 2009, 10, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Ramos, B.P.; Arnsten, A.F.T. Adrenergic pharmacology and cognition: Focus on the prefrontal cortex. Pharmacol. Ther. 2007, 113, 523–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupien, S.; Maheu, F.; Tu, M.; Fiocco, A.; Schramek, T. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007, 65, 209–237. [Google Scholar] [CrossRef] [Green Version]
- De Kloet, E.R.; Reul, J.M.; Sutanto, W. Corticosteroids and the brain. J. Steroid Biochem. Mol. Boil. 1990, 37, 387–394. [Google Scholar] [CrossRef]
- Lupien, S.J.; De Leon, M.J.; De Santi, S.; Convit, A.; Tarshish, C.; Nair, N.P.V.; Thakur, M.; McEwen, B.S.; Hauger, R.L.; Meaney, M.J. Cortisol levels during human aging predict hippocampal atrophy and memory deficits. Nat. Neurosci. 1998, 1, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, T.W.; Lovallo, W.R. Enhanced memory for emotional material following stress-level cortisol treatment in humans. Psychoneuroendocrinology 2001, 26, 307–317. [Google Scholar] [CrossRef]
- Joëls, M.; Fernández, G.; Roozendaal, B. Stress and emotional memory: A matter of timing. Trends Cogn. Sci. 2011, 15, 280–288. [Google Scholar] [CrossRef]
- Wolf, O. Stress and memory retrieval: Mechanisms and consequences. Curr. Opin. Behav. Sci. 2017, 14, 40–46. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Wolf, O.; May, M.; Wippich, W.; Hellhammer, D. Stress and treatment-induced elevations of cortisol levels associated with impaired declarative memory in healthy adults. Life Sci. 1996, 58, 1475–1483. [Google Scholar] [CrossRef]
- Roozendaal, B.; Okuda, S.; De Quervain, D.-F.; McGaugh, J. Glucocorticoids interact with emotion-induced noradrenergic activation in influencing different memory functions. Neuroscience 2006, 138, 901–910. [Google Scholar] [CrossRef]
- Barsegyan, A.; MacKenzie, S.M.; Kurose, B.D.; McGaugh, J.L.; Roozendaal, B. Glucocorticoids in the prefrontal cortex enhance memory consolidation and impair working memory by a common neural mechanism. Proc. Natl. Acad. Sci. USA 2010, 107, 16655–16660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smeets, T.; Otgaar, H.; Candel, I.; Wolf, O. True or false? Memory is differentially affected by stress-induced cortisol elevations and sympathetic activity at consolidation and retrieval. Psychoneuroendocrinology 2008, 33, 1378–1386. [Google Scholar] [CrossRef]
- Beckner, V.; Tucker, D.M.; Delville, Y.; Mohr, D. Stress facilitates consolidation of verbal memory for a film but does not affect retrieval. Behav. Neurosci. 2006, 120, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, L.; Wolf, O. Stress and multiple memory systems: From ‘thinking’ to ‘doing’. Trends Cogn. Sci. 2013, 17, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.P.; Howland, J.G.; Robillard, J.M.; Ge, Y.; Yu, W.; Titterness, A.K.; Brebner, K.; Liu, L.; Weinberg, J.; Christie, B.R.; et al. Hippocampal long-term depression mediates acute stress-induced spatial memory retrieval impairment. Proc. Natl. Acad. Sci. USA 2007, 104, 11471–11476. [Google Scholar] [CrossRef] [Green Version]
- Oei, N.Y.L.; Everaerd, W.T.A.M.; Elzinga, B.M.; Van Well, S.; Bermond, B. Psychosocial stress impairs working memory at high loads: An association with cortisol levels and memory retrieval. Stress 2006, 9, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, S.A.; Wagner, A.D. Acute stress and episodic memory retrieval: Neurobiological mechanisms and behavioral consequences. Ann. N. Y. Acad. Sci. 2016, 1369, 55–75. [Google Scholar] [CrossRef]
- Smeets, T. Acute stress impairs memory retrieval independent of time of day. Psychoneuroendocrinology 2011, 36, 495–501. [Google Scholar] [CrossRef]
- Dinse, H.R.; Kattenstroth, J.; Lenz, M.; Tegenthoff, M.; Wolf, O. The stress hormone cortisol blocks perceptual learning in humans. Psychoneuroendocrinology 2017, 77, 63–67. [Google Scholar] [CrossRef]
- Dierolf, A.M.; Fechtner, J.; Böhnke, R.; Wolf, O.; Naumann, E. Influence of acute stress on response inhibition in healthy men: An ERP study. Psychophysiology 2017, 54, 684–695. [Google Scholar] [CrossRef]
- Schoofs, D.; Preuß, D.; Wolf, O. Psychosocial stress induces working memory impairments in an n-back paradigm. Psychoneuroendocrinology 2008, 33, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Becker, L.; Rohleder, N. Time course of the physiological stress response to an acute stressor and its associations with the primacy and recency effect of the serial position curve. PLoS ONE 2019, 14, e0213883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedhara, K.; Hyde, J.; Gilchrist, I.D.; Tytherleigh, M.; Plummer, S. Acute stress, memory, attention and cortisol. Psychoneuroendocrinology 2000, 25, 535–549. [Google Scholar] [CrossRef]
- Shields, G.S.; Bonner, J.C.; Moons, W.G. Does cortisol influence core executive functions? A meta-analysis of acute cortisol administration effects on working memory, inhibition, and set-shifting. Psychoneuroendocrinology 2015, 58, 91–103. [Google Scholar] [CrossRef]
- Chun, M.M.; Turk-Browne, N.B. Interactions between attention and memory. Curr. Opin. Neurobiol. 2007, 17, 177–184. [Google Scholar] [CrossRef]
- Seger, C.A. Implicit learning. Psychol. Bull. 1994, 115, 163–196. [Google Scholar] [CrossRef]
- Rausei, V.; Makovski, T.; Jiang, Y.V. Attention dependency in implicit learning of repeated search context. Q. J. Exp. Psychol. 2007, 60, 1321–1328. [Google Scholar] [CrossRef] [Green Version]
- Chun, M.M.; Jiang, Y. Contextual cueing: Implicit learning and memory of visual context guides spatial attention. Cogn. Psychol. 1998, 36, 28–71. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Chun, M.M. Selective attention modulates implicit learning. Q. J. Exp. Psychol. Sect. A 2001, 54, 1105–1124. [Google Scholar] [CrossRef]
- Chajut, E.; Algom, D. Selective attention improves under stress: Implications for theories of social cognition. J. Pers. Soc. Psychol. 2003, 85, 231–248. [Google Scholar] [CrossRef] [Green Version]
- Booth, R.; Sharma, D. Stress reduces attention to irrelevant information: Evidence from the Stroop task. Motiv. Emot. 2009, 33, 412–418. [Google Scholar] [CrossRef]
- Sänger, J.; Bechtold, L.; Schoofs, D.; Blaszkewicz, M.; Wascher, E. The influence of acute stress on attention mechanisms and its electrophysiological correlates. Front. Behav. Neurosci. 2014, 8, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belanoff, J.K.; Gross, K.; Yager, A.; Schatzberg, A.F. Corticosteroids and cognition. J. Psychiatr. Res. 2001, 35, 127–145. [Google Scholar] [CrossRef]
- Lupien, S.J.; Gillin, C.J.; Hauger, R.L. Working memory is more sensitive than declarative memory to the acute effects of corticosteroids: A dose-response study in humans. Behav. Neurosci. 1999, 113, 420. [Google Scholar] [CrossRef]
- Putman, P.; Hermans, E.J.; Van Honk, J. Cortisol administration acutely reduces threat-selective spatial attention in healthy young men. Physiol. Behav. 2010, 99, 294–300. [Google Scholar] [CrossRef]
- Stroop, J.R. Studies of interference in serial verbal reactions. J. Exp. Psychol. 1935, 18, 643–662. [Google Scholar] [CrossRef]
- Wechsler, D. Manual for the Wechsler Adult Intelligence Scale. Psychological Corp.: New York, NY, USA, 1955. [Google Scholar]
- Becker, L.; Schade, U.; Rohleder, N. Evaluation of the socially evaluated cold-pressor group test (SECPT-G) in the general population. PeerJ 2019, 7, e7521. [Google Scholar] [CrossRef] [Green Version]
- Minkley, N.; Schröder, T.P.; Wolf, O.; Kirchner, W.H. The socially evaluated cold-pressor test (SECPT) for groups: Effects of repeated administration of a combined physiological and psychological stressor. Psychoneuroendocrinology 2014, 45, 119–127. [Google Scholar] [CrossRef]
- Schwabe, L.; Haddad, L.; Schachinger, H. HPA axis activation by a socially evaluated cold-pressor test. Psychoneuroendocrinology 2008, 33, 890–895. [Google Scholar] [CrossRef]
- Nater, U.M.; Rohleder, N.; Schlotz, W.; Ehlert, U.; Kirschbaum, C. Determinants of the diurnal course of salivary alpha-amylase. Psychoneuroendocrinology 2007, 32, 392–401. [Google Scholar] [CrossRef]
- Thoma, M.V.; Kirschbaum, C.; Wolf, J.M.; Rohleder, N. Acute stress responses in salivary alpha-amylase predict increases of plasma norepinephrine. Boil. Psychol. 2012, 91, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.A.; De Geus, E.; Veerman, E.C.I.; Hoogstraten, J.; Amerongen, A.V.N. Innate secretory immunity in response to laboratory stressors that evoke distinct patterns of cardiac autonomic activity. Psychosom. Med. 2003, 65, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Rohleder, N.; Nater, U.M. Determinants of salivary α-amylase in humans and methodological considerations. Psychoneuroendocrinology 2009, 34, 469–485. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Plessow, F.; Kirschbaum, C.; Stalder, T. Classification Criteria for distinguishing cortisol responders from nonresponders to psychosocial stress. Psychosom. Med. 2013, 75, 832–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwabe, L.; Schächinger, H. Ten years of research with the Socially Evaluated Cold Pressor Test: Data from the past and guidelines for the future. Psychoneuroendocrinology 2018, 92, 155–161. [Google Scholar] [CrossRef]
- Pruessner, J.C.; Kirschbaum, C.; Meinlschmid, G.; Hellhammer, D.H. Two formulas for computation of the area under the curve represent measures of total hormone concentration versus time-dependent change. Psychoneuroendocrinology 2003, 28, 916–931. [Google Scholar] [CrossRef]
- Mauchly, J.W. Significance test for sphericity of a normal n-variate distribution. Ann. Math. Stat. 1940, 11, 204–209. [Google Scholar] [CrossRef]
- Greenhouse, S.W.; Geisser, S. On methods in the analysis of profile data. Psychometrika 1959, 24, 95–112. [Google Scholar] [CrossRef]
- Morris, S.B. Estimating effect sizes from pretest-posttest-control group designs. Organ. Res. Methods 2007, 11, 364–386. [Google Scholar] [CrossRef]
- Sinclair, J.; Taylor, P.J.; Hobbs, S.J. Alpha level adjustments for multiple dependent variable analyses and their applicability—A review. Int. J. Sports Sci. Eng. 2013, 7, 17–20. [Google Scholar]
- Hubbard, R.; Lindsay, R.M. Why p values are not a useful measure of evidence in statistical significance testing. Theory Psychol. 2008, 18, 69–88. [Google Scholar] [CrossRef]
- Gignac, G.; Szodorai, E.T. Effect size guidelines for individual differences researchers. Pers. Individ. Differ. 2016, 102, 74–78. [Google Scholar] [CrossRef]
- Sara, S.J. The locus coeruleus and noradrenergic modulation of cognition. Nat. Rev. Neurosci. 2009, 10, 211–223. [Google Scholar] [CrossRef]
- Foote, S.L.; Morrison, J.H. Extrathalamic modulation of cortical function. Annu. Rev. Neurosci. 1987, 10, 67–95. [Google Scholar] [CrossRef]
- Valentino, R.J.; Van Bockstaele, E. Convergent regulation of locus coeruleus activity as an adaptive response to stress. Eur. J. Pharmacol. 2008, 583, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Roebuck, A.J.; Liu, M.C.; Lins, B.R.; Scott, G.A.; Howland, J.G. Acute stress, but not corticosterone, facilitates acquisition of paired associates learning in rats using touchscreen-equipped operant conditioning chambers. Behav. Brain Res. 2018, 348, 139–149. [Google Scholar] [CrossRef]
- Schacter, D.L. Priming and multiple memory systems: Perceptual mechanisms of implicit memory. J. Cogn. Neurosci. 1992, 4, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Cahill, L.; Prins, B.; Weber, M.; McGaugh, J.L. β-Adrenergic activation and memory for emotional events. Nature 1994, 371, 702–704. [Google Scholar] [CrossRef]
- Schoenfeld, B.J. Potential mechanisms for a role of metabolic stress in hypertrophic adaptations to resistance training. Sports Med. 2013, 43, 179–194. [Google Scholar] [CrossRef]
- Dickerson, S.S.; Gruenewald, T.L.; Kemeny, M.E. When the social self is threatened: Shame, physiology, and health. J. Pers. 2004, 72, 1191–1216. [Google Scholar] [CrossRef]
- Dickerson, S.S.; Kemeny, M.E. Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychol. Bull. 2004, 130, 355–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruenewald, T.L.; Dickerson, S.S.; Kemeny, M.E. The Self-Conscious Emotions: Theory and Research; Tracy, J.L., Robins, R.W., Tangney, J.P., Eds.; Guilford Press: New York, NY, USA, 2007; pp. 68–87. [Google Scholar]
- Skoluda, N.; Strahler, J.; Schlotz, W.; Niederberger, L.; Marques, S.; Fischer, S.; Thoma, M.V.; Spoerri, C.; Ehlert, U.; Nater, U.M. Intra-individual psychological and physiological responses to acute laboratory stressors of different intensity. Psychoneuroendocrinology 2015, 51, 227–236. [Google Scholar] [CrossRef]
- Becker, L.; Schade, U.; Rohleder, N. Activation of the hypothalamic-pituitary adrenal axis in response to a verbal fluency task and associations with task performance. PLoS ONE 2020, 15, e0227721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musen, G.; Treisman, A. Implicit and explicit memory for visual patterns. J. Exp. Psychol. Learn. Mem. Cogn. 1990, 16, 127. [Google Scholar] [CrossRef] [PubMed]
- Unsworth, N.; Robison, M.K. The importance of arousal for variation in working memory capacity and attention control: A latent variable pupillometry study. J. Exp. Psychol. Learn. Mem. Cogn. 2017, 43, 1962–1987. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W.; Doppelmayr, M.; Russegger, H.; Pachinger, T.; Schwaiger, J. Induced alpha band power changes in the human EEG and attention. Neurosci. Lett. 1998, 244, 73–76. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall (N = 81) | WWT (N = 12) | SECPT (N = 69) | Pre-Stress (N = 37) | Post-Stress (N = 44) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | % | N | % | N | % | N | % | N | % | |
| Female | 56 | 69.1 | 6 | 50 | 50 | 72.5 | 27 | 73 | 29 | 65.9 |
| Smokers | 2 | 2.5 | 1 | 8.3 | 1 | 1.4 | 2 | 5.4 | 0 | 0 |
| M | SD | M | SD | M | SD | M | SD | M | SD | |
| Age (years) | 21.8 | 3.9 | 22.8 | 3.6 | 21.6 | 4.0 | 21.9 | 4.0 | 21.6 | 4.0 |
| BMI 1 (kg/m2) | 22.1 | 2.8 | 23.3 | 3.4 | 21.8 | 2.6 | 22.3 | 2.9 | 21.8 | 2.7 |
| BL stress rating | 2.8 | 1.3 | 2.8 | 1.7 | 2.8 | 1.3 | 2.6 | 1.5 | 2.9 | 1.2 |
| BL sAA 2 (U/mL) | 109.7 | 71.0 | 100.4 | 49.8 | 111.4 | 74.2 | 108.6 | 68.0 | 110.7 | 74.1 |
| BL cortisol (nmol/L) | 2.9 | 2.0 | 3.6 | 1.9 | 2.7 | 2.0 | 3.0 | 2.3 | 2.8 | 1.7 |
| BL Stroop reaction time (ms) | 677.3 | 142.7 | 711.1 | 152.9 | 671.4 | 141.2 | 676.2 | 149.8 | 678.2 | 138.2 |
| BL Stroop errors | 0.8 | 1.0 | 0.8 | 1.1 | 0.8 | 1.0 | 0.8 | 1.1 | 0.7 | 0.9 |
| SECPT 1 | WWT 2 | N | |
|---|---|---|---|
| Non-responders | 9 | 3 | 12 |
| Only sAA responders | 17 | 8 | 25 |
| Only cortisol responders | 16 | 1 | 17 |
| sAA-and-cortisol responders | 27 | 0 | 27 |
| Overall | 69 | 12 | 81 |
| Overall | Non-Responders | sAA-and-Cortisol Responders | Only Cortisol Responders | Only sAA Responders | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (N = 81) | (N = 12) | (N = 27) | (N = 17) | (N = 25) | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Rating s1 | 2.8 | 1.3 | 2.7 | 1.5 | 2.7 | 1.4 | 2.9 | 1.3 | 2.8 | 1.3 |
| Rating s2 | 3.4 | 1.7 | 2.8 | 1.4 | 3.0 | 1.6 | 3.7 | 1.7 | 4.0 | 1.9 |
| Rating s3 | 3.8 | 2.0 | 3.3 | 1.9 | 3.5 | 1.7 | 4.8 | 2.2 | 3.6 | 2.2 |
| Rating s4 | 3.1 | 1.5 | 2.8 | 1.1 | 3.0 | 1.4 | 3.8 | 2.0 | 2.9 | 1.3 |
| Rating s5 | 2.6 | 1.5 | 2.3 | 1.1 | 2.4 | 1.4 | 3.3 | 2.0 | 2.4 | 1.3 |
| Rating s6 | 2.4 | 1.4 | 2.2 | 0.8 | 2.1 | 1.1 | 2.9 | 1.7 | 2.3 | 1.5 |
| Rating s7 | 2.2 | 1.2 | 1.9 | 1.0 | 2.0 | 1.1 | 2.5 | 1.4 | 2.2 | 1.1 |
| Rating s8 | 2.0 | 1.1 | 1.9 | 1.2 | 1.9 | 0.9 | 2.2 | 1.2 | 2.1 | 1.2 |
| Rating s9 | 1.9 | 0.9 | 2.0 | 1.0 | 1.7 | 0.7 | 2.2 | 1.1 | 1.8 | 1.0 |
| Rating s10 | 1.9 | 1.0 | 2.1 | 1.4 | 1.7 | 0.8 | 1.9 | 1.1 | 1.8 | 1.0 |
| Rating s11 | 1.7 | 0.8 | 1.8 | 1.0 | 1.7 | 0.8 | 1.6 | 0.7 | 1.7 | 0.9 |
| sAA s1 | 109.7 | 71.0 | 81.4 | 47.5 | 108.3 | 57.4 | 136.8 | 96.2 | 106.5 | 71.3 |
| sAA s2 | 94.2 | 71.2 | 107.0 | 68.5 | 79.2 | 46.5 | 137.7 | 101.9 | 74.7 | 58.5 |
| sAA s3 | 124.0 | 79.6 | 90.7 | 63.5 | 138.7 | 64.7 | 112.9 | 94.5 | 131.6 | 88.6 |
| sAA s4 | 119.7 | 81.6 | 82.0 | 36.7 | 125.2 | 70.7 | 149.0 | 115.9 | 112.0 | 76.0 |
| sAA s5 | 106.2 | 71.7 | 68.0 | 31.4 | 109.6 | 64.9 | 138.8 | 93.9 | 98.6 | 68.3 |
| sAA s6 | 89.7 | 61.4 | 56.6 | 34.9 | 96.6 | 56.6 | 111.1 | 76.2 | 83.5 | 60.7 |
| sAA s7 | 102.6 | 67.4 | 79.2 | 46.0 | 105.4 | 52.9 | 136.6 | 101.7 | 87.6 | 53.8 |
| sAA s8 | 104.3 | 69.0 | 81.2 | 41.6 | 106.0 | 55.7 | 132.9 | 100.2 | 94.1 | 63.6 |
| sAA s9 | 106.2 | 74.6 | 83.2 | 52.8 | 105.2 | 53.8 | 145.2 | 116.6 | 91.8 | 59.4 |
| sAA s10 | 93.2 | 70.0 | 91.3 | 57.7 | 88.4 | 54.2 | 129.1 | 104.8 | 74.7 | 54.8 |
| sAA s11 | 107.7 | 72.9 | 91.9 | 76.0 | 107.0 | 55.6 | 145.8 | 103.6 | 90.0 | 55.7 |
| Cortisol s1 | 2.9 | 2.0 | 3.3 | 2.5 | 2.4 | 1.4 | 3.8 | 2.3 | 2.5 | 1.7 |
| Cortisol s2 | 2.7 | 1.9 | 2.9 | 1.8 | 2.1 | 1.2 | 4.0 | 2.8 | 2.2 | 1.4 |
| Cortisol s3 | 3.0 | 2.6 | 3.1 | 2.4 | 2.4 | 1.4 | 5.0 | 4.1 | 2.3 | 1.5 |
| Cortisol s4 | 3.2 | 3.3 | 2.6 | 1.6 | 2.6 | 1.6 | 5.8 | 6.0 | 2.2 | 1.3 |
| Cortisol s5 | 4.2 | 4.4 | 2.5 | 1.6 | 4.5 | 2.9 | 7.8 | 7.6 | 2.3 | 1.4 |
| Cortisol s6 | 5.1 | 5.3 | 2.3 | 1.1 | 6.7 | 4.1 | 8.8 | 8.4 | 2.2 | 1.4 |
| Cortisol s7 | 6.0 | 6.7 | 2.5 | 1.4 | 7.9 | 4.9 | 11.2 | 10.8 | 2.1 | 1.3 |
| Cortisol s8 | 5.3 | 5.5 | 2.4 | 1.2 | 6.9 | 4.9 | 9.8 | 7.8 | 1.9 | 1.1 |
| Cortisol s9 | 5.2 | 5.5 | 2.9 | 2.4 | 6.7 | 4.9 | 9.3 | 8.2 | 1.9 | 1.1 |
| Cortisol s10 | 4.6 | 4.8 | 2.4 | 1.7 | 5.6 | 3.7 | 8.6 | 7.4 | 1.7 | 0.9 |
| Cortisol s11 | 4.1 | 3.9 | 2.2 | 1.3 | 4.8 | 3.2 | 7.5 | 5.5 | 1.7 | 0.8 |
| Overall | Non-Responders | sAA-and-Cortisol Responders | Only Cortisol Responders | Only sAA Responders | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (N = 81) | (N = 12) | (N = 27) | (N = 17) | (N = 25) | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| RT 1 (ms) | 677.3 | 142.7 | 706.8 | 154.3 | 652.7 | 108.6 | 729.9 | 212.9 | 653.9 | 101.5 |
| RT 2 (ms) | 654.7 | 128.0 | 682.3 | 134.4 | 637.6 | 87.9 | 708.7 | 198.9 | 623.2 | 88.1 |
| RT 3 (ms) | 624.8 | 112.9 | 656.5 | 130.9 | 598.3 | 71.7 | 673.6 | 169.9 | 604.8 | 80.4 |
| RT 4 (ms) | 606.7 | 93.2 | 631.9 | 97.0 | 585.4 | 62.0 | 650.7 | 147.9 | 587.8 | 57.8 |
| RT 5 (ms) | 602.8 | 89.2 | 606.8 | 80.6 | 585.4 | 68.5 | 648.2 | 138.8 | 588.7 | 60.0 |
| RT 6 (ms) | 604.3 | 100.9 | 630.8 | 108.8 | 576.7 | 70.0 | 642.6 | 147.6 | 595.2 | 79.8 |
| RT 7 (ms) | 594.2 | 91.8 | 618.9 | 87.5 | 565.4 | 53.5 | 637.2 | 152.9 | 584.4 | 56.9 |
| RT 8 (ms) | 590.7 | 88.6 | 614.9 | 100.5 | 561.9 | 61.1 | 626.6 | 135.4 | 585.9 | 55.8 |
| RT 9 (ms) | 588.9 | 84.8 | 594.6 | 64.6 | 577.5 | 60.9 | 629.0 | 135.2 | 571.4 | 65.2 |
| Errors 1 | 0.8 | 1.0 | 0.8 | 1.0 | 0.7 | 1.0 | 1.0 | 1.1 | 0.6 | 1.0 |
| Errors 2 | 0.8 | 1.0 | 0.8 | 1.2 | 0.6 | 0.8 | 1.1 | 1.5 | 0.8 | 0.8 |
| Errors 3 | 0.8 | 0.9 | 0.4 | 0.5 | 0.9 | 1.0 | 1.1 | 1.0 | 0.8 | 0.9 |
| Errors 4 | 0.8 | 1.0 | 0.4 | 0.9 | 0.7 | 1.1 | 1.3 | 1.2 | 0.6 | 0.7 |
| Errors 5 | 1.0 | 1.2 | 0.8 | 1.2 | 1.0 | 1.1 | 1.6 | 1.6 | 0.6 | 0.8 |
| Errors 6 | 1.0 | 1.1 | 0.7 | 1.2 | 1.0 | 1.0 | 1.1 | 1.4 | 1.0 | 1.0 |
| Errors 7 | 1.0 | 1.1 | 1.3 | 1.6 | 0.9 | 1.0 | 1.2 | 1.0 | 0.8 | 0.9 |
| Errors 8 | 1.0 | 1.1 | 1.1 | 1.2 | 0.9 | 1.2 | 0.9 | 1.1 | 1.0 | 1.2 |
| Errors 9 | 0.9 | 1.0 | 0.7 | 0.8 | 1.0 | 1.2 | 1.2 | 1.3 | 0.8 | 0.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, L.; Rohleder, N. Associations between Attention and Implicit Associative Learning in Healthy Adults: The Role of Cortisol and Salivary Alpha-Amylase Responses to an Acute Stressor. Brain Sci. 2020, 10, 544. https://doi.org/10.3390/brainsci10080544
Becker L, Rohleder N. Associations between Attention and Implicit Associative Learning in Healthy Adults: The Role of Cortisol and Salivary Alpha-Amylase Responses to an Acute Stressor. Brain Sciences. 2020; 10(8):544. https://doi.org/10.3390/brainsci10080544
Chicago/Turabian StyleBecker, Linda, and Nicolas Rohleder. 2020. "Associations between Attention and Implicit Associative Learning in Healthy Adults: The Role of Cortisol and Salivary Alpha-Amylase Responses to an Acute Stressor" Brain Sciences 10, no. 8: 544. https://doi.org/10.3390/brainsci10080544
APA StyleBecker, L., & Rohleder, N. (2020). Associations between Attention and Implicit Associative Learning in Healthy Adults: The Role of Cortisol and Salivary Alpha-Amylase Responses to an Acute Stressor. Brain Sciences, 10(8), 544. https://doi.org/10.3390/brainsci10080544