Allocentric Spatial Memory Performance in a Virtual Reality-Based Task is Conditioned by Visuospatial Working Memory Capacity

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
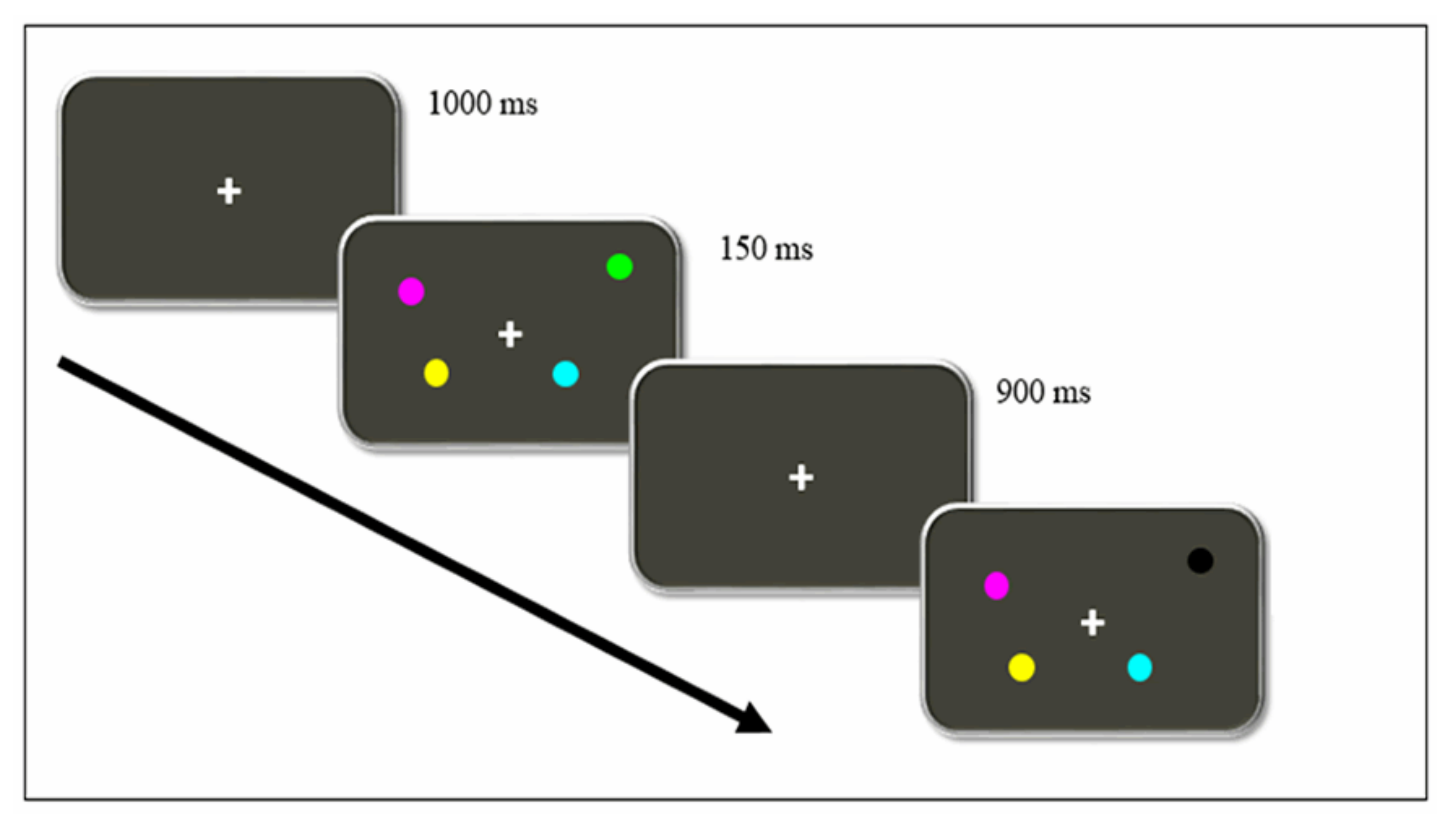
2.2. Procedure
- Working memory capacity
- Spatial memory
2.3. Statistical Procedure
3. Results
3.1. Comparisons Regarding Gender
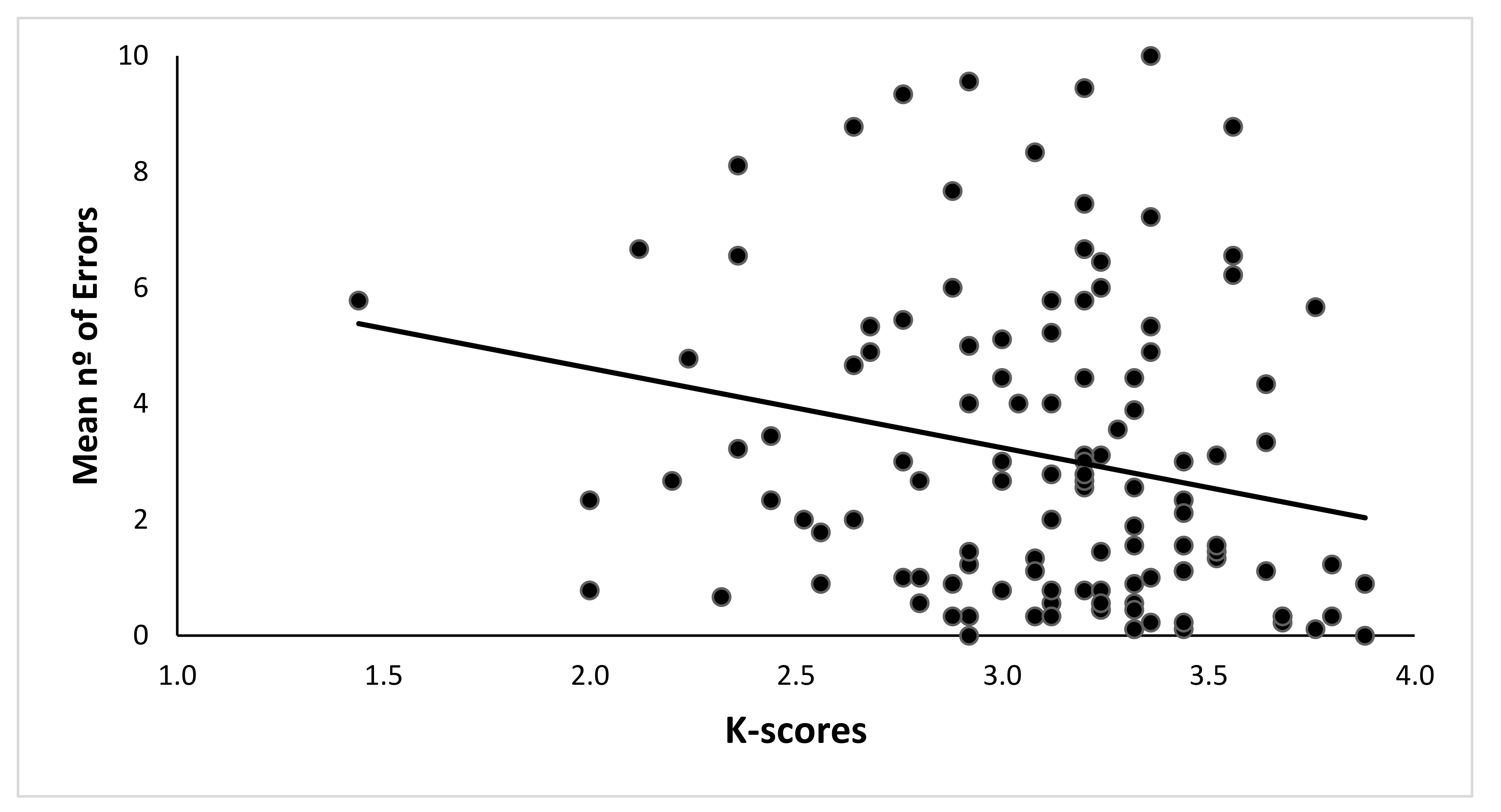
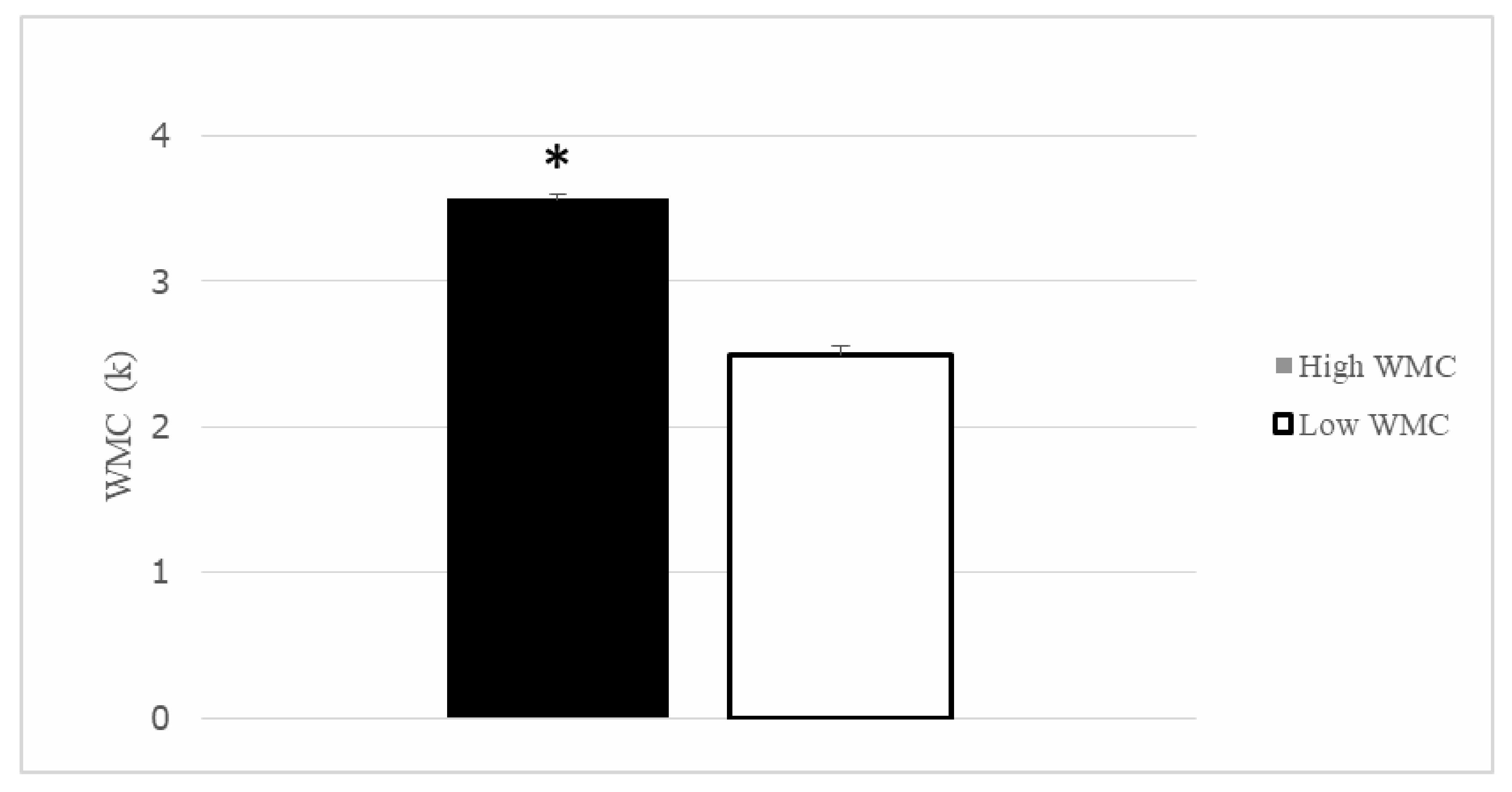
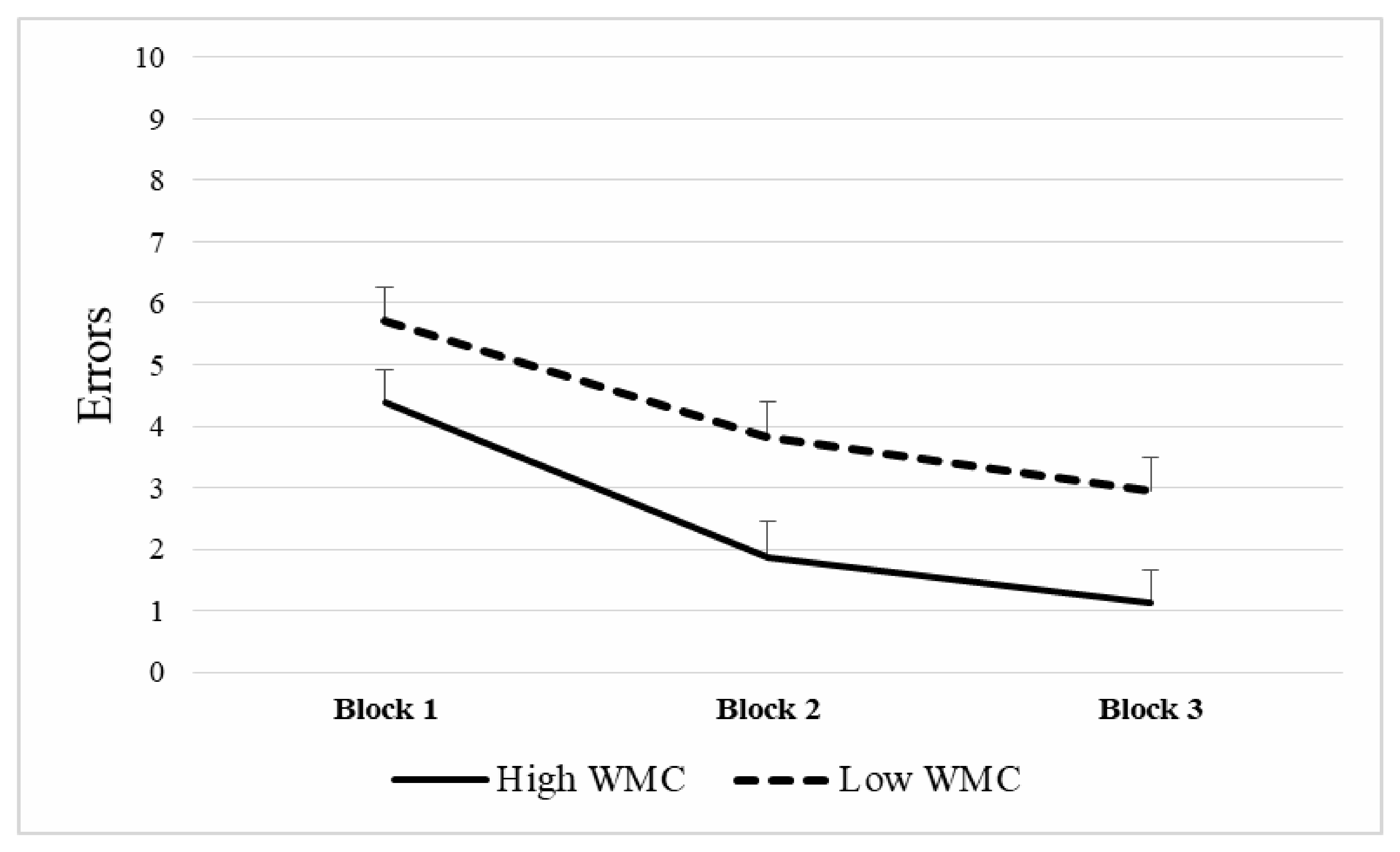
3.2. WMC and Spatial Memory Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nemmi, F.; Boccia, M.; Piccardi, L.; Galati, G.; Guariglia, C. Segregation of neural circuits involved in spatial learning in reaching and navigational space. Neuropsychologia 2013, 51, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.; Becker, S.; Burgess, N. Remembering the past and imagining the future: A neural model of spatial memory and imagery. Psychol. Rev. 2007, 114, 340–375. [Google Scholar] [CrossRef]
- Morris, R.G.M.; Garrud, P.; Rawlins, J.N.P.; O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Nadel, L. The Hippocampus as a Cognitive Map; Clarendon Press: Oxford, UK, 1978. [Google Scholar]
- Gallistel, C.R. The Organization of Learning; MIT Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Hartley, T.; Lever, C.; Burgess, N.; O’Keefe, J. Space in the brain: How the hippocampal formation supports spatial cognition. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20120510. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.A. Oscillations and hippocampal–prefrontal synchrony. Curr. Opin. Neurobiol. 2011, 21, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Sapiurka, M.; Squire, L.R.; Clark, R.E. Distinct roles of hippocampus and medial prefrontal cortex in spatial and nonspatial memory. Hippocampus 2016, 26, 1515–1524. [Google Scholar] [CrossRef] [Green Version]
- Zielinski, M.C.; Shin, J.D.; Jadhav, S.P. Coherent Coding of Spatial Position Mediated by Theta Oscillations in the Hippocampus and Prefrontal Cortex. J. Neurosci. 2019, 39, 4550–4565. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, O.; Hildebrandt, A.; Oberauer, K. What is working memory capacity, and how can we measure it? Front. Psychol. 2013, 4, 433. [Google Scholar] [CrossRef] [Green Version]
- Kane, M.J.; Engle, R.W. Working-memory capacity and the control of attention: The contributions of goal neglect, response competition, and task set to Stroop interference. J. Exp. Psychol. Gen. 2003, 132, 47–70. [Google Scholar] [CrossRef]
- Vertes, R.P. Interactions among the medial prefrontal cortex, hippocampus and midline thalamus in emotional and cognitive processing in the rat. Neuroscience 2006, 142, 1–20. [Google Scholar] [CrossRef]
- Bergmann, H.C.; Daselaar, S.M.; Fernández, G.; Kessels, R.P.C. Neural substrates of successful working memory and long-term memory formation in a relational spatial memory task. Cogn. Process. 2016, 17, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belder, M.D.; Santens, P.; Sieben, A.; Fias, W. Impaired Processing of Serial Order Determines Working Memory Impairments in Alzheimer’s Disease. J. Alzheimers Dis. 2017, 59, 1171–1186. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, L.K.; Girard, T.A.; Herdman, K.A.; Christensen, B.K.; King, J.; Kiang, M.; Bohbot, V.D. Hippocampal activation and memory performance in schizophrenia depend on strategy use in a virtual maze. Psychiatry Res. Neuroimaging 2017, 268, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, S.D.; Minshew, N.J.; Luna, B.; Sweeney, J.A. Spatial Working Memory Deficits in Autism. J. Autism. Dev. Disord. 2006, 37, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Kessels, R.P.; Haan, E.H.D.; Kappelle, L.; Postma, A. Varieties of human spatial memory: A meta-analysis on the effects of hippocampal lesions. Brain Res. Rev. 2001, 35, 295–303. [Google Scholar] [CrossRef]
- Schurgin, M.W.; Flombaum, J.I. How undistorted spatial memories can produce distorted responses. Atten. Percept. Psychophys. 2014, 76, 1371–1380. [Google Scholar] [CrossRef]
- Johnson, M.K.; Mcmahon, R.P.; Robinson, B.M.; Harvey, A.N.; Hahn, B.; Leonard, C.J.; Luck, S.J.; Gold, J.M. The relationship between working memory capacity and broad measures of cognitive ability in healthy adults and people with schizophrenia. Neuropsychology 2013, 27, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Noguera, C.; Fernández, S.; Álvarez, D.; Carmona, E.; Marí-Beffa, P.; Ortells, J.J. The implementation of expectancy-based strategic processes is delayed in normal aging. PLoS ONE 2019, 14, e0214322. [Google Scholar] [CrossRef]
- Ortells, J.J.; Fockert, J.W.D.; Romera, N.; Fernández, S. Expectancy-Based Strategic Processes Are Influenced by Spatial Working Memory Load and Individual Differences in Working Memory Capacity. Front. Psychol. 2018, 9, 1239. [Google Scholar] [CrossRef]
- Astur, R.S.; Tropp, J.; Sava, S.; Constable, R.; Markus, E.J. Sex differences and correlations in a virtual Morris water task, a virtual radial arm maze, and mental rotation. Behav. Brain Res. 2004, 151, 103–115. [Google Scholar] [CrossRef]
- Tascón, L.; Castillo, J.; León, I.; Cimadevilla, J.M. Walking and non-walking space in an equivalent virtual reality task: Sexual dimorphism and aging decline of spatial abilities. Behav. Brain Res. 2018, 347, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Moodley, K.; Minati, L.; Contarino, V.; Prioni, S.; Wood, R.; Cooper, R.; D’Incerti, L.; Tagliavini, F.; Chan, D. Diagnostic differentiation of mild cognitive impairment due to Alzheimers disease using a hippocampus-dependent test of spatial memory. Hippocampus 2015, 25, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Rosas, K.; Parrón, I.; Serrano, P.; Cimadevilla, J.M. Spatial recognition memory in a virtual reality task is altered in refractory temporal lobe epilepsy. Epilepsy Behav. 2013, 28, 227–231. [Google Scholar] [CrossRef]
- Lind, S.E.; Williams, D.M.; Raber, J.; Peel, A.; Bowler, D.M. Spatial navigation impairments among intellectually high-functioning adults with autism spectrum disorder: Exploring relations with theory of mind, episodic memory, and episodic future thinking. J. Abnorm. Psychol. 2013, 122, 1189–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, R.; Leon, I.; Roldan, M.D.; Astur, R.; Cimadevilla, J.M. Virtual reality tasks disclose spatial memory alterations in fibromyalgia. Rheumatology 2009, 48, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Cánovas, R.; Espínola, M.; Iribarne, L.; Cimadevilla, J.M. A new virtual task to evaluate human place learning. Behav. Brain Res. 2008, 190, 112–118. [Google Scholar] [CrossRef]
- León, I.; Tascón, L.; Ortells-Pareja, J.J.; Cimadevilla, J.M. Virtual reality assessment of walking and non-walking space in men and women with virtual reality-based tasks. PLoS ONE 2018, 13, e0204995. [Google Scholar] [CrossRef] [Green Version]
- Marchette, S.A.; Sever, M.W.; Flombaum, J.I.; Shelton, A.L. Individual Differences in Representational Precision Predict Spatial Working Memory Span. Spat. Cogn. Comput. 2015, 15, 308–328. [Google Scholar] [CrossRef]
- Cowan, N.; Elliott, E.M.; Saults, J.S.; Morey, C.C.; Mattox, S.; Hismjatullina, A.; Conway, A.R. On the capacity of attention: Its estimation and its role in working memory and cognitive aptitudes. Cogn. Psychol. 2005, 51, 42–100. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, K.A. Attentional Control and the relatedness proportion effects on semantic priming. J. Exp. Psychol. Learn. Mem. Cogn. 2007, 33, 645–662. [Google Scholar] [CrossRef] [Green Version]
- Richmond, L.L.; Redick, T.S.; Braver, T.S. Remembering to prepare: The benefits (and costs) of high working memory capacity. J. Exp. Psychol. Learn. Mem. Cogn. 2015, 41, 1764–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engle, R. Role of Working-Memory Capacity in Cognitive Control. Curr. Anthr. 2010, 51, S17–S26. [Google Scholar] [CrossRef] [Green Version]
- Conway, A.; Kane, M.; Bunting, M.; Hambrick, Z.; Wilhem, O.; Engle, R. Working memory span task: A methodological review and user’s guide. Psychol. Bull. Rev. 2005, 12, 769–786. [Google Scholar] [CrossRef] [PubMed]
- Shipstead, Z.; Lindsey, D.R.; Marshall, R.L.; Engle, R.W. The mechanisms of working memory capacity: Primary memory, secondary memory, and attention control. J. Mem. Lang. 2014, 72, 116–141. [Google Scholar] [CrossRef]
- Hockey, R. The Psychology of Fatigue: Work, Effort and Control; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Wurm, L.H.; Legge, G.E.; Isenberg, L.M.; Luebker, A. Color improves object recognition in normal and low vision. J. Exp. Psychol. Hum. Percept. Perform. 1993, 19, 899–911. [Google Scholar] [CrossRef]
- Cánovas, M.R.; Cimadevilla, J.M. Sexual orientation and spatial memory. Psicothema 2011, 23, 752–758. [Google Scholar]
- Sánchez-Horcajo, R.; Llamas-Alonso, J.; Cimadevilla, J.M. Practice of Aerobic Sports is Associated with Better Spatial Memory in Adults and Older Men. Exp. Aging Res. 2015, 41, 193–203. [Google Scholar] [CrossRef]
- Cimadevilla, J.M.; Lizana, J.R.; Roldán, M.D.; Cánovas, R.; Rodríguez, E. Spatial memory alterations in children with epilepsy of genetic origin or unknown cause. Epileptic Disord. 2014, 16, 203–207. [Google Scholar] [CrossRef] [Green Version]
- León, I.; Cimadevilla, J.M.; Tascón, L. Developmental gender differences in children in a virtual spatial memory task. Neuropsychology 2014, 28, 485–495. [Google Scholar] [CrossRef]
- Chamizo, V.D. Spatial learning: Conditions and basic effects. Psicologica 2002, 23, 33–57. [Google Scholar]
- Wolbers, T.; Hegarty, M. What determines our navigational abilities? Trends Cogn. Sci. 2010, 14, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuis, I.L.; Takashima, A. The role of the ventromedial prefrontal cortex in memory consolidation. Behav. Brain Res. 2011, 218, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyman, J.M.; Zilli, E.A.; Paley, A.M.; Hasselmo, M.E. Working Memory Performance Correlates with Prefrontal-Hippocampal Theta Interactions but not with Prefrontal Neuron Firing Rates. Front. Integr. Neurosci. 2010, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.W.; Wilson, M.A. Theta Rhythms Coordinate Hippocampal–Prefrontal Interactions in a Spatial Memory Task. PLoS Biol. 2005, 3, e402. [Google Scholar] [CrossRef] [Green Version]
- Kyd, R.J.; Bilkey, D.K. Prefrontal cortex lesions modify the spatial properties of hippocampal place cells. Cereb. Cortex. 2003, 13, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, A.P. The role of the prefrontal cortex in dynamic filtering. Psychobiology 2000, 28, 207–218. [Google Scholar] [CrossRef]
- Martinet, L.-E.; Sheynikhovich, D.; Benchenane, K.; Arleo, A. Spatial Learning and Action Planning in a Prefrontal Cortical Network Model. PLoS Comput. Biol. 2011, 7, e1002045. [Google Scholar] [CrossRef]
- Barker, G.R.I.; Bird, F.; Alexander, V.; Warburton, E.C. Recognition Memory for Objects, Place, and Temporal Order: A Disconnection Analysis of the Role of the Medial Prefrontal Cortex and Perirhinal Cortex. J. Neurosci. 2007, 27, 2948–2957. [Google Scholar] [CrossRef] [Green Version]
- Barker, G.R.I.; Banks, P.J.; Scott, H.; Ralph, G.S.; Mitrophanous, K.A.; Wong, L.-F.; Bashir, Z.; Uney, J.B.; Warburton, E.C. Separate elements of episodic memory subserved by distinct hippocampal–prefrontal connections. Nat. Neurosci. 2017, 20, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Fallon, S.J.; Zokaei, N.; Husain, M. Causes and consequences of limitations in visual working memory. Ann. N. Y. Acad. Sci. 2016, 1369, 40–54. [Google Scholar] [CrossRef] [Green Version]
- León, I.; Tascón, L.; Cimadevilla, J.M. Age and gender-related differences in a spatial memory task in humans. Behav. Brain Res. 2016, 306, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, R.; García, R.F.; Cimadevilla, J.M. Effect of reference frames and number of cues available on the spatial orientation of males and females in a virtual memory task. Behav. Brain Res. 2011, 216, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Chai, X.J.; Jacobs, L.F. Effects of cue types on sex differences in human spatial memory. Behav. Brain Res. 2010, 208, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Nori, R.; Piccardi, L.; Maialetti, A.; Goro, M.; Rossetti, A.; Argento, O.; Guariglia, C. No Gender Differences in Egocentric and Allocentric Environmental Transformation after Compensating for Male Advantage by Manipulating Familiarity. Front. Neurosci. 2018, 12, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tascón, L.; León, I.; Cimadevilla, J.M. Viewpoint-related gender differences in a spatial recognition task. Learn. Individ. Differ. 2016, 50, 270–274. [Google Scholar] [CrossRef]
- Moffat, S.D.; Resnick, S.M. Effects of age on virtual environment place navigation and allocentric cognitive mapping. Behav. Neurosci. 2002, 116, 851–859. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Block 1 (Trials 2–4) | Block 2 (Trials 5–7) | Block 3 (Trials 8–10) | |
|---|---|---|---|
| K-Index | −0.176 | −0.237 | −0.180 |
| Significance | 0.051 | 0.008 | 0.046 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo Escamilla, J.; Fernández Castro, J.J.; Baliyan, S.; Ortells-Pareja, J.J.; Ortells Rodríguez, J.J.; Cimadevilla, J.M. Allocentric Spatial Memory Performance in a Virtual Reality-Based Task is Conditioned by Visuospatial Working Memory Capacity. Brain Sci. 2020, 10, 552. https://doi.org/10.3390/brainsci10080552
Castillo Escamilla J, Fernández Castro JJ, Baliyan S, Ortells-Pareja JJ, Ortells Rodríguez JJ, Cimadevilla JM. Allocentric Spatial Memory Performance in a Virtual Reality-Based Task is Conditioned by Visuospatial Working Memory Capacity. Brain Sciences. 2020; 10(8):552. https://doi.org/10.3390/brainsci10080552
Chicago/Turabian StyleCastillo Escamilla, Joaquín, José Javier Fernández Castro, Shishir Baliyan, Juan José Ortells-Pareja, Juan José Ortells Rodríguez, and José Manuel Cimadevilla. 2020. "Allocentric Spatial Memory Performance in a Virtual Reality-Based Task is Conditioned by Visuospatial Working Memory Capacity" Brain Sciences 10, no. 8: 552. https://doi.org/10.3390/brainsci10080552