Synuclein Deficiency Results in Age-Related Respiratory and Cardiovascular Dysfunctions in Mice


Abstract
:1. Introduction
2. Methods
2.1. Ethical Approval
2.2. Generation of Single-, Double- and Triple-Synuclein-Null Mutant Animals
2.3. Physiological Experiments in Conscious Mice
2.3.1. Non-Invasive Breathing Measurement
2.3.2. Non-Invasive Blood Pressure Measurement
2.4. Anesthetized Preparation for Baroreflex Measurements
2.4.1. Surgical Preparation
2.4.2. Baroreflex Protocol
2.4.3. Morphometries
2.5. Data Analysis
3. Results
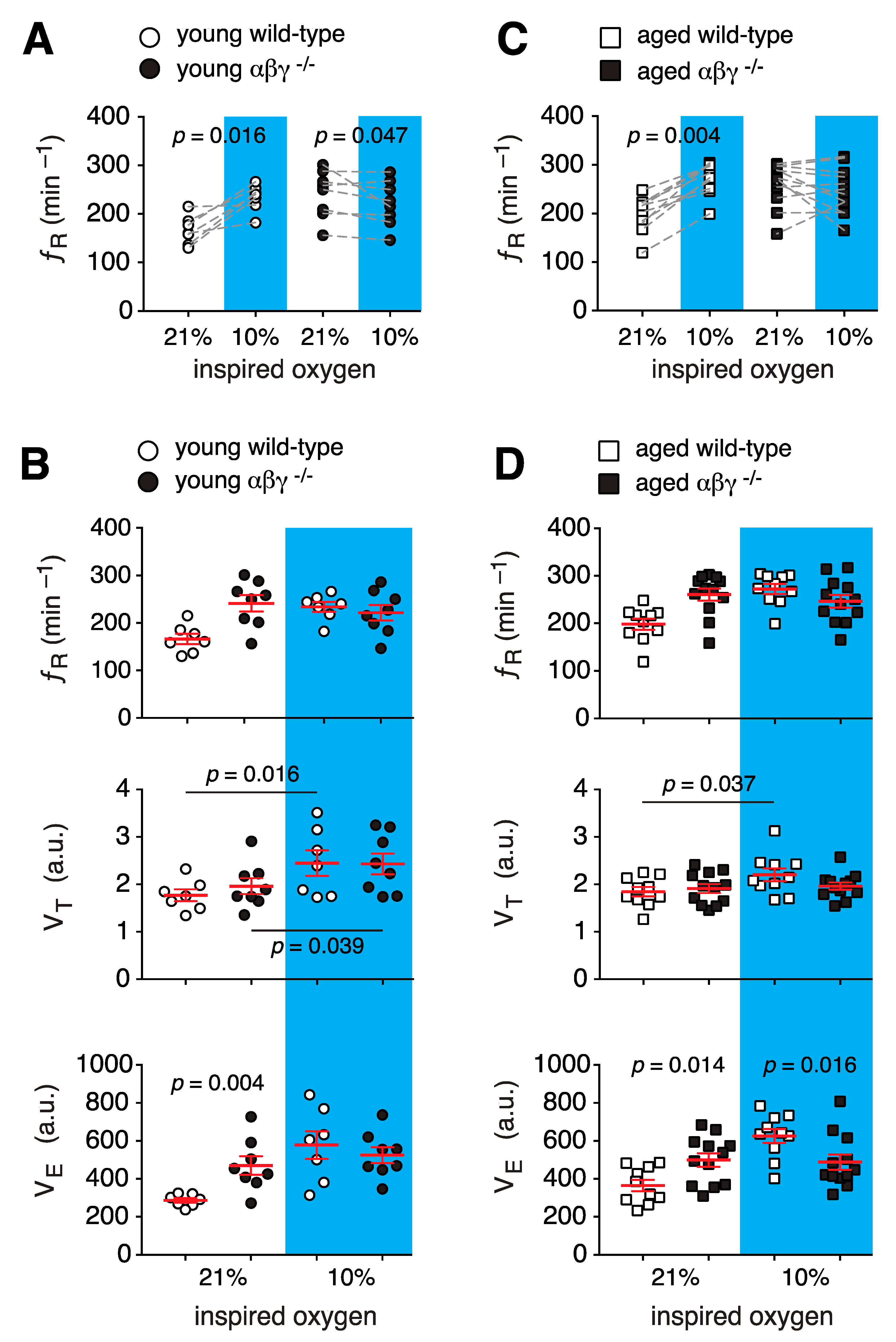
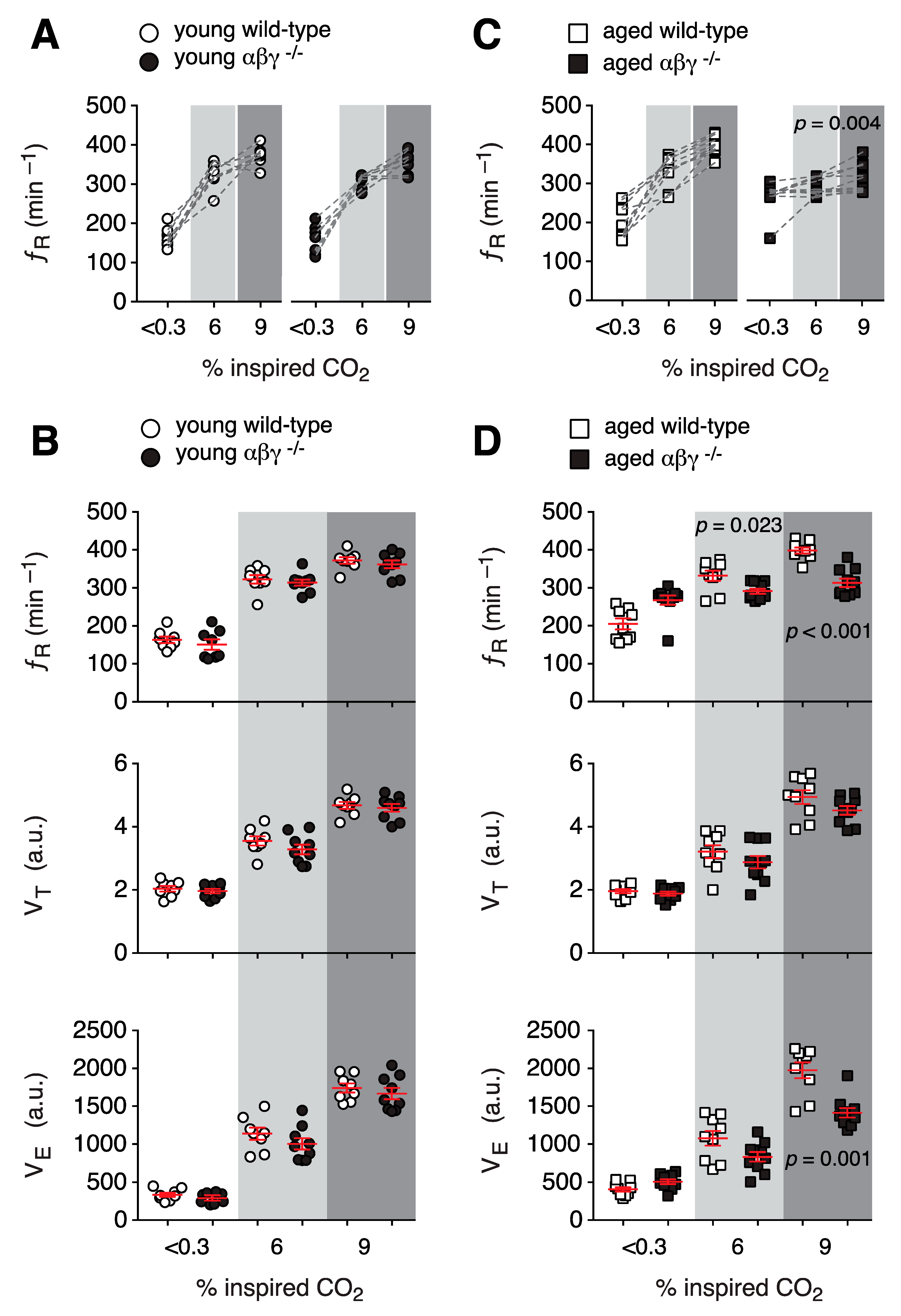
3.1. Respiratory Activity in Conscious Mice
3.1.1. Resting Breathing in Room Air and Hyperoxic Condition
3.1.2. Ventilatory Response to Hypoxia
3.1.3. Ventilatory Response to Hypercapnia
3.2. Heart Morphometry Parameters
3.3. Hemodynamics in Conscious Mice
3.4. Hemodynamics under Anesthesia
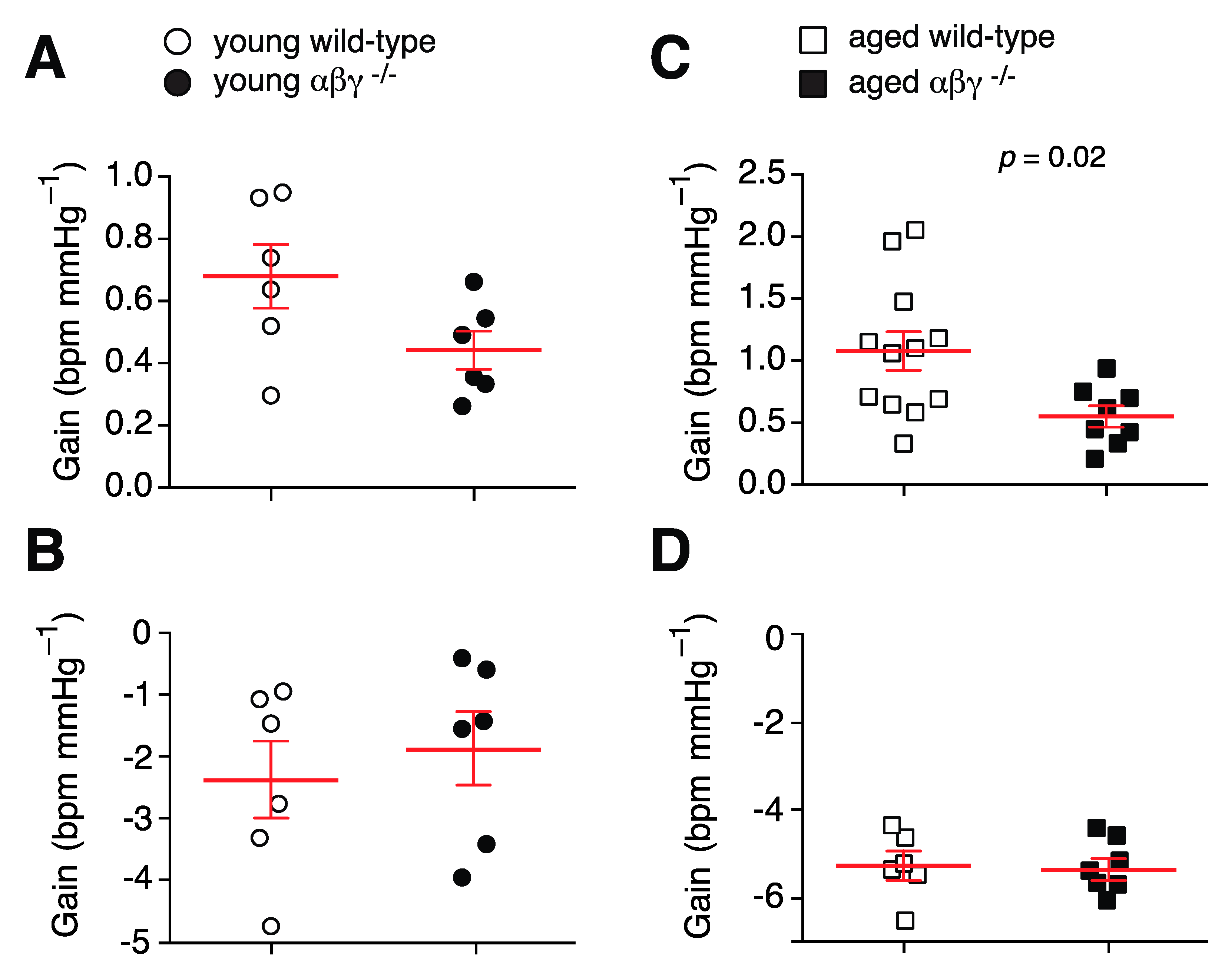
3.5. Baroreflex
3.5.1. Baroreflex Sensitivity
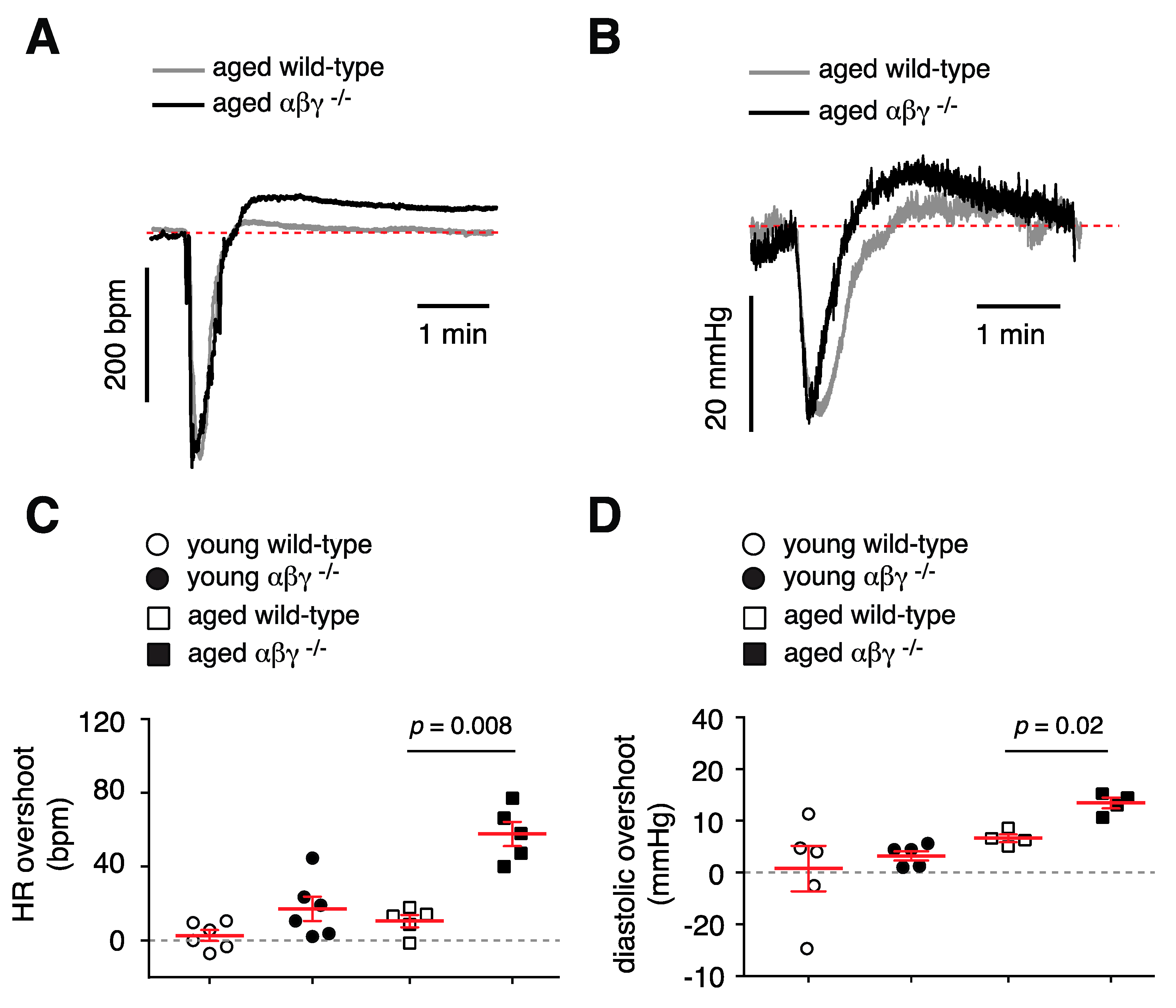
3.5.2. Baroreflex Recovery
4. Discussion
4.1. Respiratory Function
4.2. Cardiovascular Function
5. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clayton, D.F.; George, J.M. Synucleins in synaptic plasticity and neurodegenerative disorders. J. Neurosci. Res. 1999, 58, 120–129. [Google Scholar] [CrossRef]
- Burré, J.; Sharma, M.; Tsetsenis, T.; Buchman, V.; Etherton, M.R.; Südhof, T.C. α-Synuclein promotes SNARE-complex assembly in vivo and in vitro. Science 2010, 329, 1663–1667. [Google Scholar] [CrossRef] [Green Version]
- Burré, J. The Synaptic Function of α-Synuclein. J. Park. Dis. 2015, 5, 699–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polymeropoulos, M.H. Mutation in the α-Synuclein Gene Identified in Families with Parkinson’s Disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visanji, N.P.; Brotchie, J.M.; Kalia, L.V.; Koprich, J.B.; Tandon, A.; Watts, J.C.; Lang, A.E. α-Synuclein-Based Animal Models of Parkinson’s Disease: Challenges and Opportunities in a New Era. Trends Neurosci. 2016, 39, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Venda, L.L.; Cragg, S.J.; Buchman, V.L.; Wade-Martins, R. α-Synuclein and dopamine at the crossroads of Parkinson’s disease. Trends Neurosci. 2010, 33, 559–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, S.; Fornai, F.; Kwon, H.-B.; Yazdani, U.; Atasoy, D.; Liu, X.; Hammer, R.E.; Battaglia, G.; German, D.C.; Castillo, P.E.; et al. Double-knockout mice for alpha- and beta-synucleins: Effect on synaptic functions. Proc. Natl. Acad. Sci. USA 2004, 101, 14966–14971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.C.; Schmidt, O.; Ninkina, N.; Jones, P.A.; Sharkey, J.; Buchman, V.L. Developmental loss and resistance to MPTP toxicity of dopaminergic neurones in substantia nigra pars compacta of γ-synuclein, α-synuclein and double α/γ-synuclein null mutant mice. J. Neurochem. 2004, 89, 1126–1136. [Google Scholar] [CrossRef] [PubMed]
- Maroteaux, L.; Campanelli, J.T.; Scheller, R.H. Synuclein: A neuron-specific protein localized to the nucleus and presynaptic nerve terminal. J. Neurosci. 1988, 8, 2804–2815. [Google Scholar] [CrossRef] [Green Version]
- Al-Wandi, A.; Ninkina, N.; Millership, S.J.; Williamson, S.J.; Jones, P.A.; Buchman, V.L. Absence of α-synuclein affects dopamine metabolism and synaptic markers in the striatum of aging mice. Neurobiol. Aging 2010, 31, 796–804. [Google Scholar] [CrossRef] [Green Version]
- Connor-Robson, N.; Peters, O.; Millership, S.; Ninkina, N.; Buchman, V.L. Combinational losses of synucleins reveal their differential requirements for compensating age-dependent alterations in motor behavior and dopamine metabolism. Neurobiol. Aging 2016, 46, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninkina, N.; Tarasova, T.V.; Chaprov, K.D.; Roman, A.Y.; Kukharsky, M.S.; Kolik, L.G.; Ovchinnikov, R.; Ustyugov, A.A.; Durnev, A.D.; Buchman, V.L. Alterations in the nigrostriatal system following conditional inactivation of α-synuclein in neurons of adult and aging mice. Neurobiol. Aging 2020, 91, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Greten-Harrison, B.; Polydoro, M.; Morimoto-Tomita, M.; Diao, L.; Williams, A.M.; Nie, E.H.; Makani, S.; Tian, N.; Castillo, P.E.; Buchman, V.L.; et al. αβγ-Synuclein triple knockout mice reveal age-dependent neuronal dysfunction. Proc. Natl. Acad. Sci. USA 2010, 107, 19573–19578. [Google Scholar] [CrossRef] [Green Version]
- Anwar, S.; Peters, O.; Millership, S.J.; Ninkina, N.; Doig, N.; Connor-Robson, N.; Threlfell, S.; Kooner, G.; Deacon, R.M.; Bannerman, D.M.; et al. Functional alterations to the nigrostriatal system in mice lacking all three members of the synuclein family. J. Neurosci. 2011, 31, 7264–7274. [Google Scholar] [CrossRef] [PubMed]
- Tomić, S.; Rajkovaca, I.; Pekic, V.; Salha, T.; Misevic, S. Impact of autonomic dysfunctions on the quality of life in Parkinson’s disease patients. Acta Neurol. Belg. 2016, 117, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Merola, A.; Romagnolo, A.; Rosso, M.; Suri, R.; Berndt, Z.; Maule, S.; Lopiano, L.; Espay, A.J. Autonomic dysfunction in Parkinson’s disease: A prospective cohort study. Mov. Disord. 2018, 33, 391–397. [Google Scholar] [CrossRef]
- Micieli, G.; Tosi, P.; Marcheselli, S.; Cavallini, A. Autonomic dysfunction in Parkinson’s disease. Neurol. Sci. 2003, 24, s32–s34. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; De Vos, R.A.; Steur, E.N.J.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Braak, H.; Sastre, M.; Bohl, J.R.E.; De Vos, R.A.I.; Del Tredici, K. Parkinson’s disease: Lesions in dorsal horn layer I, involvement of parasympathetic and sympathetic pre- and postganglionic neurons. Acta Neuropathol. 2007, 113, 421–429. [Google Scholar] [CrossRef]
- Kaufmann, H.; Nahm, K.; Purohit, D.; Wolfe, D. Autonomic failure as the initial presentation of Parkinson disease and dementia with Lewy bodies. Neurology 2004, 63, 1093–1095. [Google Scholar] [CrossRef]
- Metzger, J.M.; Emborg, M. Autonomic dysfunction in Parkinson disease and animal models. Clin. Auton. Res. 2019, 29, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Onodera, M.; Kuwaki, T.; Kumada, M.; Masuda, Y. Determination of Ventilatory Volume in Mice by Whole Body Plethysmography. Jpn. J. Physiol. 1997, 47, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Sheikhbahaei, S.; Gourine, A.V.; Smith, J.C. Respiratory rhythm irregularity after carotid body denervation in rats. Respir. Physiol. Neurobiol. 2017, 246, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Sheikhbahaei, S.; Turovsky, E.A.; Hosford, P.S.; Hadjihambi, A.; Theparambil, S.M.; Liu, B.; Marina, N.; Teschemacher, A.G.; Kasparov, S.; Smith, J.C.; et al. Astrocytes modulate brainstem respiratory rhythm-generating circuits and determine exercise capacity. Nat. Commun. 2018, 9, 370. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Whitesall, S.; Zhang, Y.; Beibel, M.; Alecy, L.D.; DiPetrillo, K. Validation of Volume-Pressure Recording Tail-Cuff Blood Pressure Measurements. Am. J. Hypertens. 2008, 21, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- David, J.M.; Chatziioannou, A.F.; Taschereau, R.; Wang, H.; Stout, D.B. The Hidden Cost of Housing Practices: Using Noninvasive Imaging to Quantify the Metabolic Demands of Chronic Cold Stress of Laboratory Mice. Comp. Med. 2013, 63, 386–391. [Google Scholar]
- Devine, M.J.; Gwinn-Hardy, K.; Singleton, A.; Hardy, J. Parkinson’s disease and α-synuclein expression. Mov. Disord. 2011, 26, 2160–2168. [Google Scholar] [CrossRef] [Green Version]
- Martin, I.; Dawson, V.L.; Dawson, V.L. Recent advances in the genetics of Parkinson’s disease. Annu. Rev. Genom. Hum. Genet. 2011, 12, 301–325. [Google Scholar] [CrossRef] [Green Version]
- Eschbach, J.; Danzer, K.M. α-Synuclein in Parkinson’s disease: Pathogenic function and translation into animal models. Neurodegener. Dis. 2014, 14, 1–17. [Google Scholar] [CrossRef]
- Lee, Y.; Dawson, V.L.; Dawson, T.M. Animal Models of Parkinson’s Disease: Vertebrate Genetics. Cold Spring Harb. Perspect. Med. 2012, 2, a009324. [Google Scholar] [CrossRef]
- Crabtree, D.M.; Zhang, J. Genetically engineered mouse models of Parkinson’s disease. Brain Res. Bull. 2012, 88, 13–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesselet, M.-F.; Richter, F. Modelling of Parkinson’s disease in mice. Lancet Neurol. 2011, 10, 1108–1118. [Google Scholar] [CrossRef]
- Duty, S.; Jenner, P. Animal models of Parkinson’s disease: A source of novel treatments and clues to the cause of the disease. Br. J. Pharmacol. 2011, 164, 1357–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinta, S.J.; Mallajosyula, J.K.; Rane, A.; Andersen, J.K. Mitochondrial alpha-synuclein accumulation impairs complex I function in dopaminergic neurons and results in increased mitophagy in vivo. Neurosci. Lett. 2010, 486, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Mullin, S.; Schapira, A.H.V. α-Synuclein and mitochondrial dysfunction in Parkinson’s disease. Mol. Neurobiol. 2013, 47, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, R.K.; Beal, M.F. Mitochondrial Diseases of the Brain. Free Radic. Boil. Med. 2013, 63, 1–29. [Google Scholar] [CrossRef]
- Di Maio, R.; Barrett, P.J.; Hoffman, E.K.; Barrett, C.W.; Zharikov, A.; Borah, A.; Hu, X.; McCoy, J.; Chu, C.T.; Burton, E.A.; et al. α-Synuclein binds to TOM20 and inhibits mitochondrial protein import in Parkinson’s disease. Sci. Transl. Med. 2016, 8, 342ra78. [Google Scholar] [CrossRef] [Green Version]
- Angelova, P.R.; Kasymov, V.; Christie, I.; Sheikhbahaei, S.; Turovsky, E.; Marina, N.; Korsak, A.; Zwicker, J.; Teschemacher, A.G.; Ackland, G.L.; et al. Functional Oxygen Sensitivity of Astrocytes. J. Neurosci. 2015, 35, 10460–10473. [Google Scholar] [CrossRef] [Green Version]
- Gourine, A.V.; Funk, G.D. On the existence of a central respiratory oxygen sensor. J. Appl. Physiol. 2017, 123, 1344–1349. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.C.; Ellenberger, H.H.; Ballanyi, K.; Richter, D.W.; Feldman, J.L. Pre-Botzinger complex: A brainstem region that may generate respiratory rhythm in mammals. Science 1991, 254, 726–729. [Google Scholar] [CrossRef]
- Del Negro, C.A.; Funk, G.D.; Feldman, J.L. Breathing matters. Nat. Rev. Neurosci. 2018, 19, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Rajani, V.; Zhang, Y.; Jalubula, V.; Rancic, V.; Sheikhbahaei, S.; Zwicker, J.D.; Pagliardini, S.; Dickson, C.; Ballanyi, K.; Kasparov, S.; et al. Release of ATP by pre-Bötzinger complex astrocytes contributes to the hypoxic ventilatory response via a Ca2+ -dependent P2Y1 receptor mechanism. J. Physiol. 2017, 596, 3245–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludtmann, M.H.R.; Angelova, P.R.; Ninkina, N.; Gandhi, S.; Buchman, V.L.; Abramov, A.Y. Monomeric Alpha-Synuclein Exerts a Physiological Role on Brain ATP Synthase. J. Neurosci. 2016, 36, 10510–10521. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K. α-Synuclein and Mitochondria: Partners in Crime? Neurotherapeutics 2013, 10, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Tuppy, M.; Barna, B.; Alves-Dos-Santos, L.; Britto, L.; Chiavegatto, S.; Moreira, T.; Takakura, A.C. Respiratory deficits in a rat model of Parkinson’s disease. Neuroscience 2015, 297, 194–204. [Google Scholar] [CrossRef]
- Andrzejewski, K.; Budzińska, K.; Kaczyńska, K. Phrenic and hypoglossal nerve activity during respiratory response to hypoxia in 6-OHDA unilateral model of Parkinson’s disease. Life Sci. 2017, 180, 143–150. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Tuppy, M.; Moreira, T.S.; Takakura, A.C. Role of the locus coeruleus catecholaminergic neurons in the chemosensory control of breathing in a Parkinson’s disease model. Exp. Neurol. 2017, 293, 172–180. [Google Scholar] [CrossRef]
- Lima, J.C.; Oliveira, L.M.; Botelho, M.T.; Moreira, T.S.; Takakura, A.C. The involvement of the pathway connecting the substantia nigra, the periaqueductal gray matter and the retrotrapezoid nucleus in breathing control in a rat model of Parkinson’s disease. Exp. Neurol. 2018, 302, 46–56. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Oliveira, M.A.; Moriya, H.T.; Moreira, T.S.; Takakura, A.C. Respiratory disturbances in a mouse model of Parkinson’s disease. Exp. Physiol. 2019, 104, 729–739. [Google Scholar] [CrossRef]
- Machhada, A.; Ang, R.; Ackland, G.L.; Ninkina, N.; Buchman, V.L.; Lythgoe, M.F.; Trapp, S.; Tinker, A.; Marina, N.; Gourine, A.V. Control of ventricular excitability by neurons of the dorsal motor nucleus of the vagus nerve. Hear. Rhythm. 2015, 12, 2285–2293. [Google Scholar] [CrossRef] [Green Version]
- Wirth, A.; Wang, S.; Takefuji, M.; Tang, C.; Althoff, T.F.; Schweda, F.; Wettschureck, N.; Offermanns, S. Age-dependent blood pressure elevation is due to increased vascular smooth muscle tone mediated by G-protein signaling. Cardiovasc. Res. 2015, 109, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, T.S.; Antunes, V.R.; Falquetto, B.; Marina, N. Long-term stimulation of cardiac vagal preganglionic neurons reduces blood pressure in the spontaneously hypertensive rat. J. Hypertens. 2018, 36, 2444–2452. [Google Scholar] [CrossRef] [PubMed]
- Hotta, H.; Uchida, S. Aging of the autonomic nervous system and possible improvements in autonomic activity using somatic afferent stimulation. Geriatr. Gerontol. Int. 2010, 10, S127–S136. [Google Scholar] [CrossRef] [PubMed]
- Mancia, G.; Grassi, G. The Autonomic Nervous System and Hypertension. Circ. Res. 2014, 114, 1804–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, J.-A.; Kaufmann, H. Treatment of autonomic dysfunction in Parkinson disease and other synucleinopathies. Mov. Disord. 2018, 33, 372–390. [Google Scholar] [CrossRef]
- Isonaka, R.; Rosenberg, A.Z.; Sullivan, P.; Corrales, A.; Holmes, C.; Sharabi, Y.; Goldstein, D.S. Alpha-Synuclein Deposition Within Sympathetic Noradrenergic Neurons Is Associated With Myocardial Noradrenergic Deficiency in Neurogenic Orthostatic Hypotension. Hypertension 2019, 73, 910–918. [Google Scholar] [CrossRef]
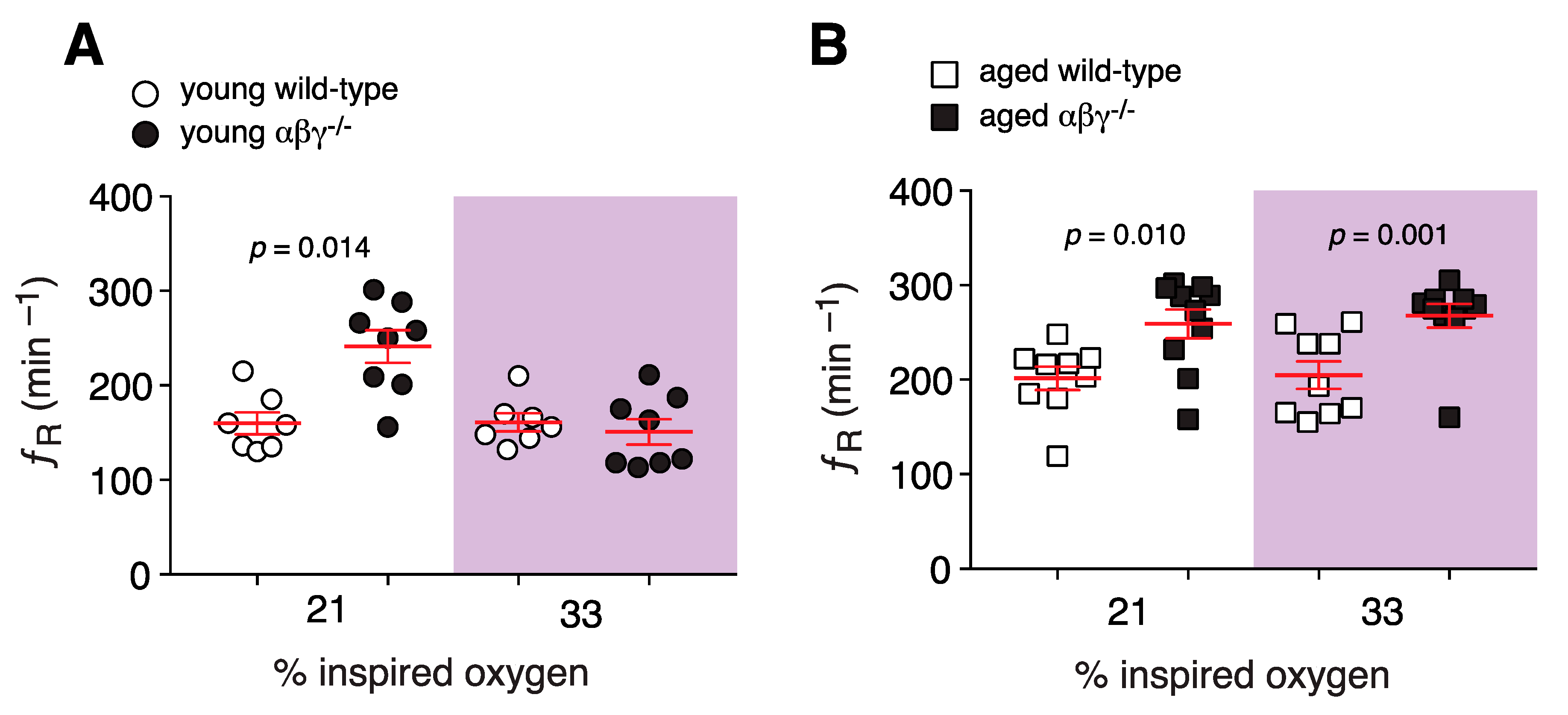

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young Wild Type | Young αβγ−/− | p Value | Aged Wild Type | Aged αβγ−/− | p Value | |
|---|---|---|---|---|---|---|
| n | 6 | 6 | 6 | 6 | ||
| Body Weight (g) | 26.8 ± 0.5 | 28.2 ± 0.6 | 0.13 | 27.2 ± 0.7 | 25.6 ± 0.8 | 0.08 |
| Heart Weight (g) | 0.156 ± 0.005 | 0.157 ± 0.006 | 0.85 | 0.183 ± 0.2 | 0.157 ± 0.01 | 0.18 |
| LV Weight (g) | 0.088 ± 0.006 | 0.091 ± 0.002 | 0.79 | 0.113 ± 0.1 | 0.092 ± 0.005 | 0.14 |
| HW/BW Ratio | 5.78 ± 0.04 | 5.59 ± 0.20 | 0.23 | 6.71 ± 0.5 | 6.39 ± 0.4 | 0.53 |
| Young Wild Type | Young αβγ−/− | p Value | Aged Wild Type | Aged αβγ−/− | p Value | |
|---|---|---|---|---|---|---|
| n | 8 | 9 | 8 | 8 | ||
| Diastolic BP (mmHg) | 86.7 ± 4.2 | 87.8 ± 3.4 | 0.94 | 121 ± 5.9 | 147 ± 6.9 | 0.02 |
| Systolic BP (mmHg) | 121.9 ± 5.1 | 125.0 ± 3.6 | 0.72 | 161 ± 7.0 | 185 ± 7.0 | 0.02 |
| Mean BP (mmHg) | 98.1 ± 4.4 | 99.8 ± 3.4 | 0.87 | 133.9 ± 6.2 | 159.5 ± 6.9 | 0.02 |
| Heart rate (bpm) | 568 ± 33 | 572.0 ±1 9 | 0.73 | 599 ± 37 | 632 ± 9 | 0.92 |
| Aged Wild Type | Aged α−/− | p Value | Aged β−/− | p Value | Aged γ−/− | p Value | Aged αγ−/− | p Value | |
|---|---|---|---|---|---|---|---|---|---|
| n | 8 | 3 | 5 | 3 | 3 | ||||
| Diastolic BP (mmHg) | 119 ± 7 | 130 ± 6 | 0.38 | 137 ± 6 | 0.093 | 118 ± 8 | 0.667 | 120 ± 5 | 0.67 |
| Systolic BP (mmHg) | 159 ± 6 | 163 ± 5 | 0.92 | 171 ± 6 | 0.617 | 152 ± 7 | 0.170 | 155 ± 6 | 0.17 |
| Mean BP (mmHg) | 132 ± 6 | 141 ± 5 | 0.63 | 148 ± 6 | 0.127 | 129 ± 8 | 0.667 | 131 ± 5 | 0.55 |
| Heart rate (bpm) | 572 ± 45 | 636 ± 38 | >0.99 | 651 ± 11 | 0.343 | 625 ± 49 | 0.667 | 601 ± 19 | 0.33 |
| Young Wild Type | Young αβγ−/− | p Value | Aged Wild Type | Aged αβγ−/− | p Value | |
|---|---|---|---|---|---|---|
| n | 5 | 5 | 8 | 8 | ||
| Diastolic BP (mmHg) | 78 ± 4 | 80 ± 5 | 0.67 | 72 ± 3 | 79 ± 4 | 0.71 |
| Systolic BP (mmHg) | 107 ± 3 | 109 ± 5 | 0.74 | 96 ± 4.0 | 94 ± 5 | 0.77 |
| Mean BP (mmHg) | 65 ± 5 | 67 ± 5 | 0.65 | 83 ± 4 | 85 ± 4 | 0.92 |
| Heart rate (bpm) | 658 ± 7 | 623 ± 22 | 0.22 | 575 ± 33 | 626 ± 25 | 0.36 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosford, P.S.; Ninkina, N.; Buchman, V.L.; Smith, J.C.; Marina, N.; SheikhBahaei, S. Synuclein Deficiency Results in Age-Related Respiratory and Cardiovascular Dysfunctions in Mice. Brain Sci. 2020, 10, 583. https://doi.org/10.3390/brainsci10090583
Hosford PS, Ninkina N, Buchman VL, Smith JC, Marina N, SheikhBahaei S. Synuclein Deficiency Results in Age-Related Respiratory and Cardiovascular Dysfunctions in Mice. Brain Sciences. 2020; 10(9):583. https://doi.org/10.3390/brainsci10090583
Chicago/Turabian StyleHosford, Patrick S., Natalia Ninkina, Vladimir L. Buchman, Jeffrey C. Smith, Nephtali Marina, and Shahriar SheikhBahaei. 2020. "Synuclein Deficiency Results in Age-Related Respiratory and Cardiovascular Dysfunctions in Mice" Brain Sciences 10, no. 9: 583. https://doi.org/10.3390/brainsci10090583