Ipsi- and Contralateral Oligo- and Polysynaptic Reflexes in Humans Revealed by Low-Frequency Epidural Electrical Stimulation of the Lumbar Spinal Cord

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Subjects
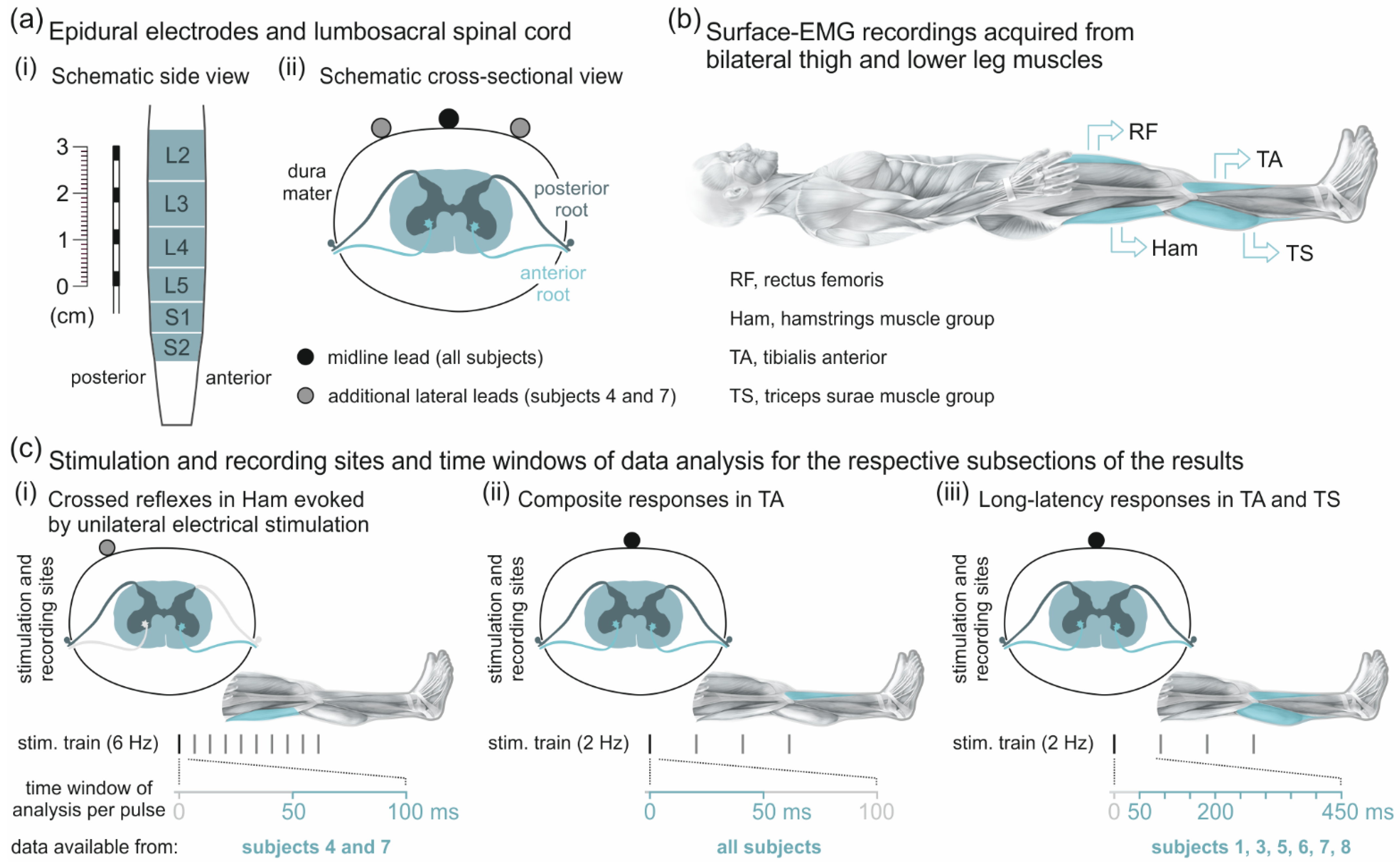
2.2. Epidural Electrical Stimulation of the Spinal Cord and EMG Recordings
2.3. Data Analysis and Statistics
2.3.1. Analysis of Responses to Unilateral Epidural Electrical Stimulation
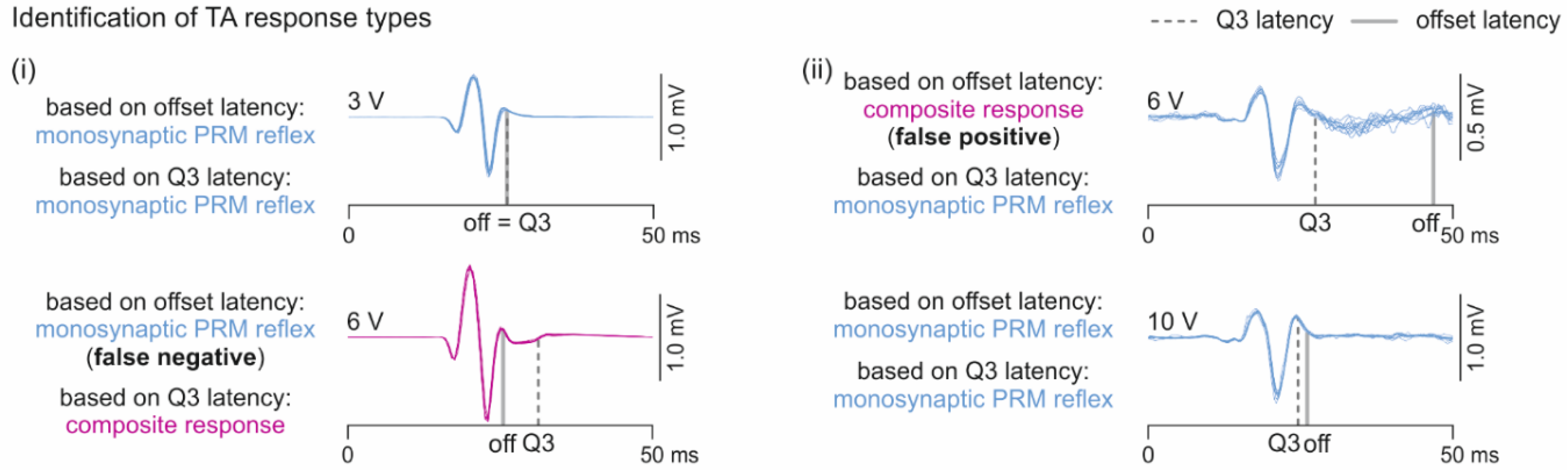
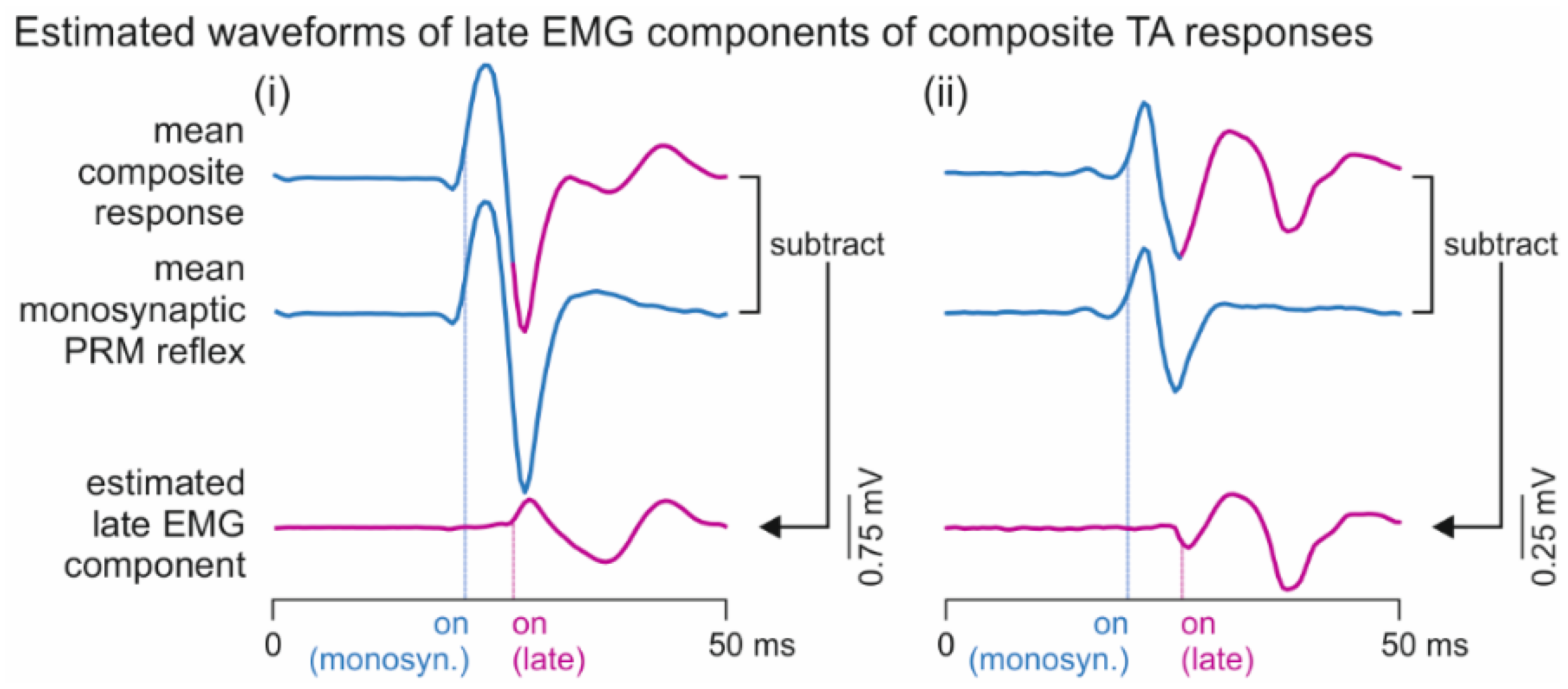
2.3.2. Analysis of Responses in Tibialis Anterior with and without Additional Late EMG Components
2.3.3. Descending Polysynaptic Responses
3. Results
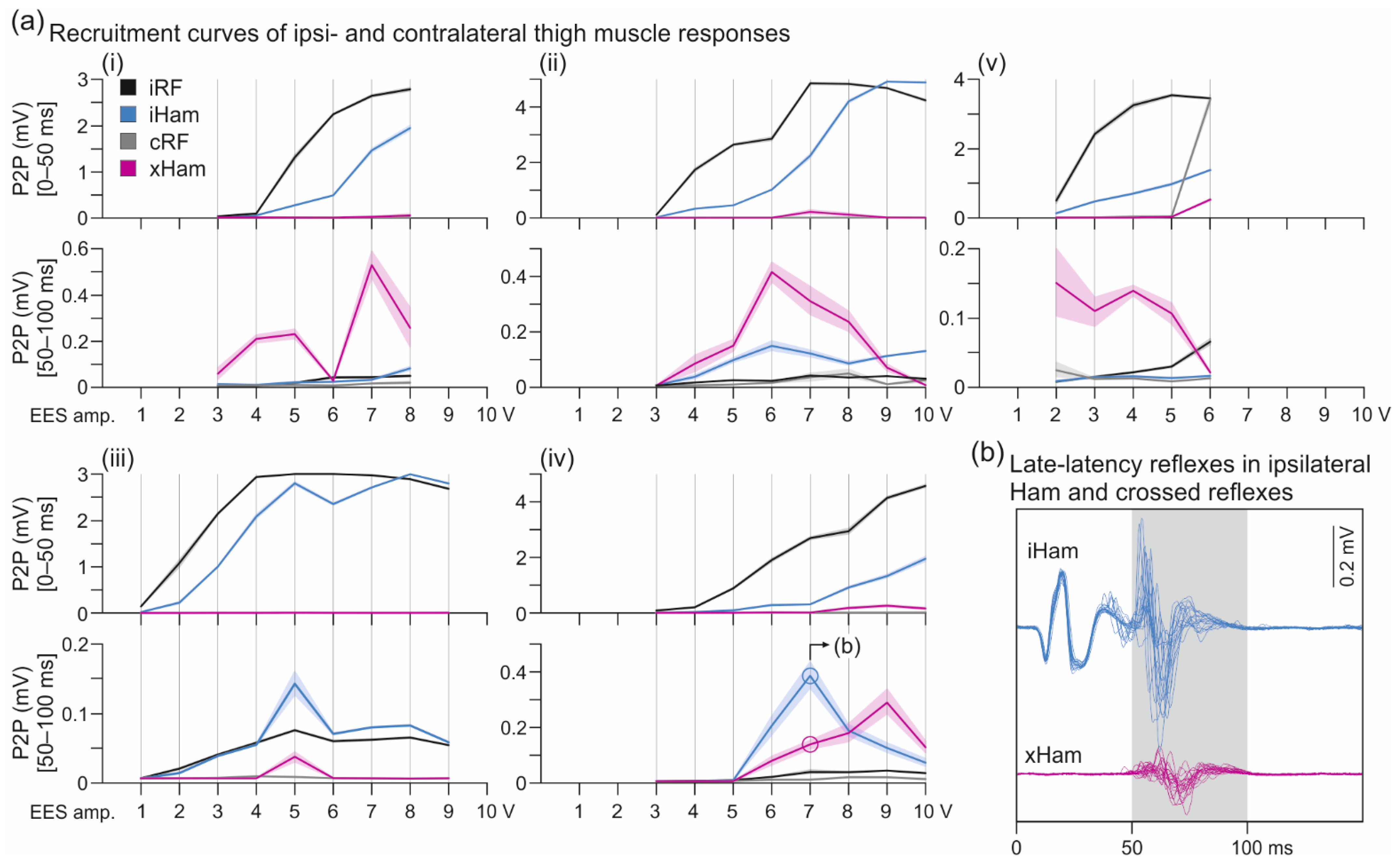
3.1. Crossed Reflexes in the Hamstrings Muscle Group
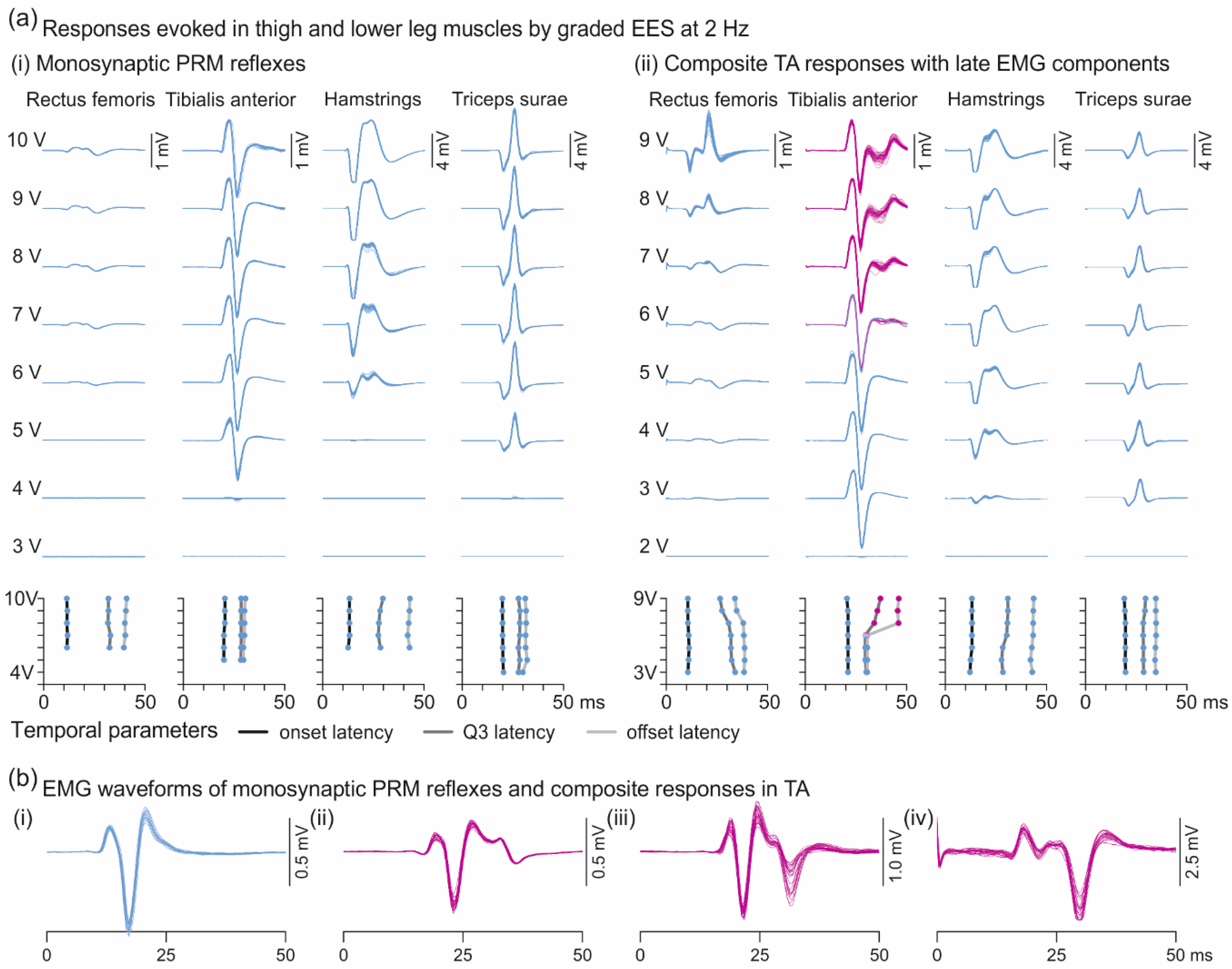
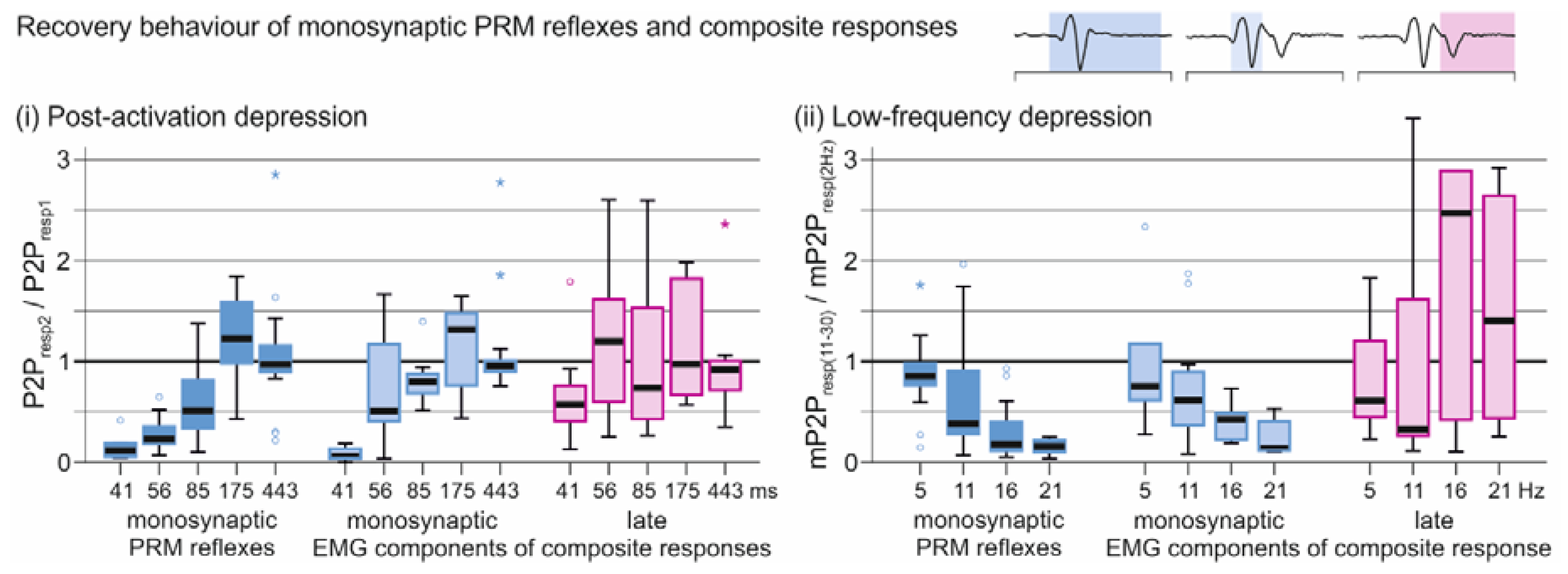
3.2. Oligosynaptic Reflexes in Tibialis Anterior
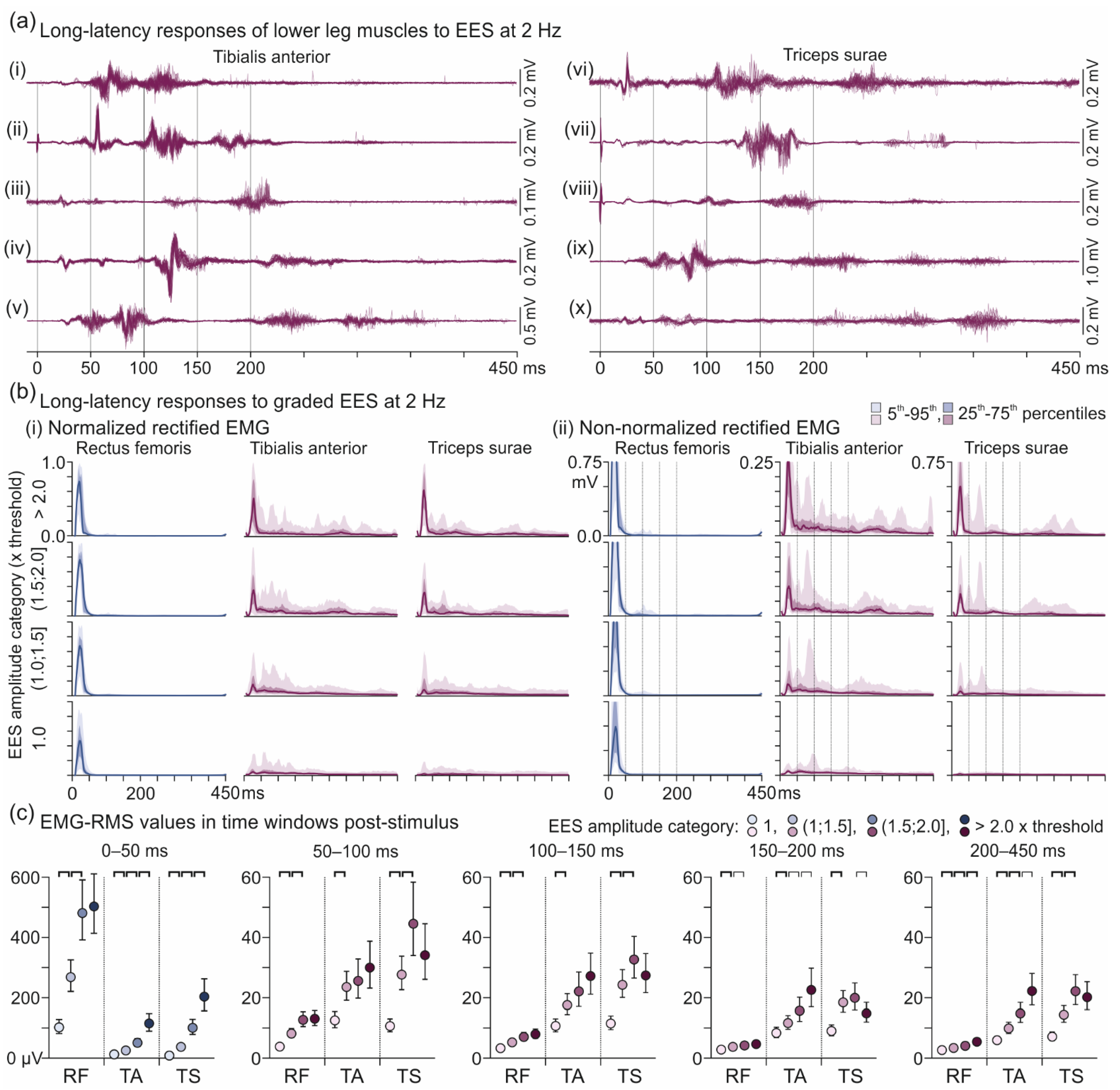
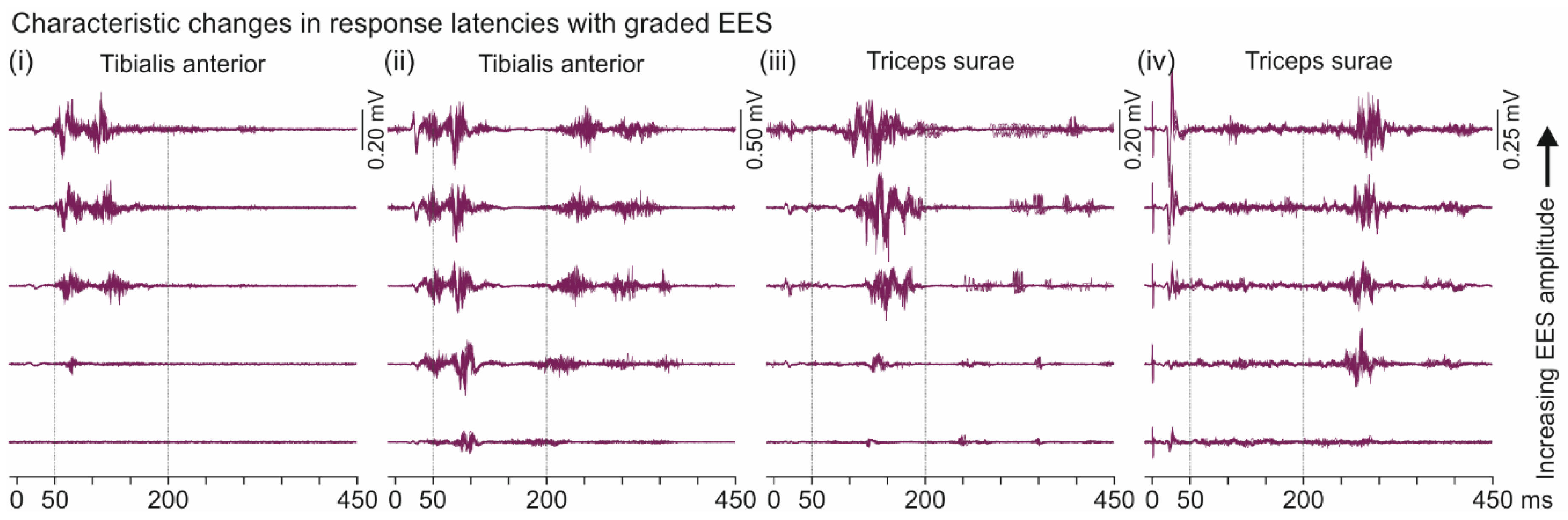
3.3. Descending Polysynaptic Responses in Tibialis Anterior and the Triceps Surae Muscle Group
4. Discussion
4.1. Crossed Reflexes in the Hamstrings Muscle Group
4.2. Oligosynaptic Reflexes in Tibialis Anterior
4.3. Descending Polysynaptic Responses in Tibialis Anterior and the Triceps Surae Muscle Group
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Wagner, F.B.; Mignardot, J.-B.; Le Goff-Mignardot, C.G.; Demesmaeker, R.; Komi, S.; Capogrosso, M.; Rowald, A.; Seáñez, I.; Caban, M.; Pirondini, E.; et al. Targeted neurotechnology restores walking in humans with spinal cord injury. Nature 2018, 563, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Gill, M.L.; Grahn, P.J.; Calvert, J.S.; Linde, M.B.; Lavrov, I.A.; Strommen, J.A.; Beck, L.A.; Sayenko, D.G.; Van Straaten, M.G.; Drubach, D.I.; et al. Neuromodulation of lumbosacral spinal networks enables independent stepping after complete paraplegia. Nat. Med. 2018, 24, 1677–1682. [Google Scholar] [CrossRef] [PubMed]
- Angeli, C.A.; Boakye, M.; Morton, R.A.; Vogt, J.; Benton, K.; Chen, Y.; Ferreira, C.K.; Harkema, S.J. Recovery of Over-Ground Walking after Chronic Motor Complete Spinal Cord Injury. N. Engl. J. Med. 2018, 379, 1244–1250. [Google Scholar] [CrossRef]
- Calvert, J.S.; Grahn, P.J.; Zhao, K.D.; Lee, K.H. Emergence of Epidural Electrical Stimulation to Facilitate Sensorimotor Network Functionality after Spinal Cord Injury. Neuromodul. Technol. Neural Interface 2019, 22, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Minassian, K.; Persy, I.; Rattay, F.; Pinter, M.M.; Kern, H.; Dimitrijevic, M.R. Human lumbar cord circuitries can be activated by extrinsic tonic input to generate locomotor-like activity. Hum. Mov. Sci. 2007, 26, 275–295. [Google Scholar] [CrossRef]
- Minassian, K.; McKay, W.B.; Binder, H.; Hofstoetter, U.S. Targeting Lumbar Spinal Neural Circuitry by Epidural Stimulation to Restore Motor Function After Spinal Cord Injury. Neurotherapeutics 2016, 13, 284–294. [Google Scholar] [CrossRef]
- Capogrosso, M.; Wenger, N.; Raspopovic, S.; Musienko, P.; Beauparlant, J.; Bassi Luciani, L.; Courtine, G.; Micera, S. A Computational Model for Epidural Electrical Stimulation of Spinal Sensorimotor Circuits. J. Neurosci. 2013, 33, 19326–19340. [Google Scholar] [CrossRef]
- Formento, E.; Minassian, K.; Wagner, F.; Mignardot, J.B.; Le Goff-Mignardot, C.G.; Rowald, A.; Bloch, J.; Micera, S.; Capogrosso, M.; Courtine, G. Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury. Nat. Neurosci. 2018, 21, 1728–1741. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Perret, I.; Bayart, A.; Lackner, P.; Binder, H.; Freundl, B.; Minassian, K. Spinal motor mapping by epidural stimulation of lumbosacral posterior roots in humans. iScience 2020, 101930. [Google Scholar] [CrossRef]
- Lloyd, D. Neuron patterns controlling transmission of ipsilateral hind limb reflexes in cat. J Neurophysiol. 1943, 6, 293–315. [Google Scholar] [CrossRef]
- Lloyd, D.P.C.; McIntyre, A.K. Dorsal column conduction of group I muscle afferent impulses and their relay through Clarke’s column. J. Neurophysiol. 1950, 13, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Davidoff, R.A. The dorsal columns. Neurology 1989, 39, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Waltz, J.M. Spinal cord stimulation: A quarter century of development and investigation. A review of its development and effectiveness in 1,336 cases. Stereotact. Funct. Neurosurg. 1997, 69, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, J.; Lazorthes, Y.; Broggi, G. Electrical spinal cord stimulation for spastic movement disorders. Appl. Neurophysiol. 1981, 44, 77–92. [Google Scholar] [CrossRef]
- Minassian, K.; Hofstoetter, U.; Tansey, K.; Mayr, W. Neuromodulation of lower limb motor control in restorative neurology. Clin. Neurol. Neurosurg. 2012, 114, 489–497. [Google Scholar] [CrossRef]
- Cook, A.W.; Weinstein, S.P. Chronic dorsal column stimulation in multiple sclerosis. Preliminary report. N. Y. State J. Med. 1973, 73, 2868–2872. [Google Scholar]
- Jankowska, E. Interneuronal relay in spinal pathways from proprioceptors. Prog. Neurobiol. 1992, 38, 335–378. [Google Scholar] [CrossRef]
- Duysens, J.; De Groote, F.; Jonkers, I. The flexion synergy, mother of all synergies and father of new models of gait. Front. Comput. Neurosci. 2013, 7, 14. [Google Scholar] [CrossRef]
- Kiehn, O. Decoding the organization of spinal circuits that control locomotion. Nat. Rev. Neurosci. 2016, 17, 224–238. [Google Scholar] [CrossRef]
- Hultborn, H.; Conway, B.A.; Gossard, J.P.; Brownstone, R.; Fedirchuk, B.; Schomburg, E.D.; Enríquez-Denton, M.; Perreault, M.C. How do we approach the locomotor network in the mammalian spinal cord? Ann. N. Y. Acad. Sci. 1998, 860, 70–82. [Google Scholar] [CrossRef]
- Minassian, K.; Jilge, B.; Rattay, F.; Pinter, M.M.; Binder, H.; Gerstenbrand, F.; Dimitrijevic, M.R. Stepping-like movements in humans with complete spinal cord injury induced by epidural stimulation of the lumbar cord: Electromyographic study of compound muscle action potentials. Spinal Cord 2004, 42, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Minassian, K.; Freundl, B.; Hofstoetter, U.S. The posterior root-muscle reflex. In Neurophysiology in Neurosurgery; Elsevier: Amsterdam, The Netherlands, 2020; pp. 239–253. [Google Scholar]
- Hofstoetter, U.S.; Freundl, B.; Binder, H.; Minassian, K. Common neural structures activated by epidural and transcutaneous lumbar spinal cord stimulation: Elicitation of posterior root-muscle reflexes. PLoS ONE 2018, 13, e0192013. [Google Scholar] [CrossRef] [PubMed]
- Hofstoetter, U.S.; Danner, S.M.; Freundl, B.; Binder, H.; Mayr, W.; Rattay, F.; Minassian, K. Periodic modulation of repetitively elicited monosynaptic reflexes of the human lumbosacral spinal cord. J. Neurophysiol. 2015, 114, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijevic, M.R.; Gerasimenko, Y.; Pinter, M.M. Evidence for a spinal central pattern generator in humans. Ann. N. Y. Acad. Sci. 1998, 860, 360–376. [Google Scholar] [CrossRef]
- Minassian, K.; Hofstoetter, U.S.; Dzeladini, F.; Guertin, P.A.; Ijspeert, A. The Human Central Pattern Generator for Locomotion: Does It Exist and Contribute to Walking? Neuroscience 2017, 23, 649–663. [Google Scholar] [CrossRef]
- Danner, S.M.; Hofstoetter, U.S.; Freundl, B.; Binder, H.; Mayr, W.; Rattay, F.; Minassian, K. Human spinal locomotor control is based on flexibly organized burst generators. Brain 2015, 138, 577–588. [Google Scholar] [CrossRef]
- Wenger, N.; Moraud, E.M.; Gandar, J.; Musienko, P.; Capogrosso, M.; Baud, L.; Le Goff, C.G.; Barraud, Q.; Pavlova, N.; Dominici, N.; et al. Spatiotemporal neuromodulation therapies engaging muscle synergies improve motor control after spinal cord injury. Nat. Med. 2016, 22, 138–145. [Google Scholar] [CrossRef]
- Capogrosso, M.; Milekovic, T.; Borton, D.; Wagner, F.; Moraud, E.M.; Mignardot, J.-B.; Buse, N.; Gandar, J.; Barraud, Q.; Xing, D.; et al. A brain–spine interface alleviating gait deficits after spinal cord injury in primates. Nature 2016, 539, 284–288. [Google Scholar] [CrossRef]
- Gerasimenko, Y.P.; Lavrov, I.A.; Courtine, G.; Ichiyama, R.M.; Dy, C.J.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Spinal cord reflexes induced by epidural spinal cord stimulation in normal awake rats. J. Neurosci. Methods 2006, 157, 253–263. [Google Scholar] [CrossRef]
- Lavrov, I.; Gerasimenko, Y.P.; Ichiyama, R.M.; Courtine, G.; Zhong, H.; Roy, R.R.; Edgerton, V.R. Plasticity of spinal cord reflexes after a complete transection in adult rats: Relationship to stepping ability. J. Neurophysiol. 2006, 96, 1699–1710. [Google Scholar] [CrossRef]
- Sayenko, D.G.; Angeli, C.; Harkema, S.J.; Edgerton, V.R.; Gerasimenko, Y.P. Neuromodulation of evoked muscle potentials induced by epidural spinal-cord stimulation in paralyzed individuals. J. Neurophysiol. 2014, 111, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Pinter, M.M.; Gerstenbrand, F.; Dimitrijevic, M.R. Epidural electrical stimulation of posterior structures of the human lumbosacral cord: 3. Control Of spasticity. Spinal Cord 2000, 38, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378. [Google Scholar] [CrossRef]
- Thage, O. The myotomes L2-S2 in man. Acta Neurol. Scand. 2009, 41, 241–244. [Google Scholar] [CrossRef]
- Schirmer, C.M.; Shils, J.L.; Arle, J.E.; Cosgrove, G.R.; Dempsey, P.K.; Tarlov, E.; Kim, S.; Martin, C.J.; Feltz, C.; Moul, M.; et al. Heuristic map of myotomal innervation in humans using direct intraoperative nerve root stimulation. J. Neurosurg. Spine 2011, 15, 64–70. [Google Scholar] [CrossRef]
- Jankowska, E. Spinal interneuronal networks in the cat: Elementary components. Brain Res. Rev. 2008, 57, 46–55. [Google Scholar] [CrossRef]
- Schindler-Ivens, S.; Shields, R.K. Low frequency depression of H-reflexes in humans with acute and chronic spinal-cord injury. Exp. Brain Res. 2000, 133, 233–241. [Google Scholar] [CrossRef]
- Elbasiouny, S.M.; Moroz, D.; Bakr, M.M.; Mushahwar, V.K. Management of spasticity after spinal cord injury: Current techniques and future directions. Neurorehabil. Neural Repair 2010, 24, 23–33. [Google Scholar] [CrossRef]
- Lamy, J.C.; Wargon, I.; Baret, M.; Ben Smail, D.; Milani, P.; Raoul, S.; Pénicaud, A.; Katz, R. Post-activation depression in various group I spinal pathways in humans. Exp. Brain Res. 2005, 166, 248–262. [Google Scholar] [CrossRef]
- Hammar, I.; Slawinska, U.; Jankowska, E. A comparison of postactivation depression of synaptic actions evoked by different afferents and at different locations in the feline spinal cord. Exp. Brain Res. 2002, 145, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, A.J.T.; Kamavuako, E.N.; Geertsen, S.S.; Farina, D.; Mrachacz-Kersting, N. Short-latency crossed responses in the human biceps femoris muscle. J. Physiol. 2015, 593, 3657–3671. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, A.J.T.; Geertsen, S.S.; Andersen, J.B.; Sinkjaer, T.; Nielsen, J.B.; Mrachacz-Kersting, N. Interlimb communication to the knee flexors during walking in humans. J. Physiol. 2013, 591, 4921–4935. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E.; Noga, B.R. Contralaterally projecting lamina VIII interneurones in middle lumbar segments in the cat. Brain Res. 1990, 535, 327–330. [Google Scholar] [CrossRef]
- Jankowska, E.; Bannatyne, B.A.; Stecina, K.; Hammar, I.; Cabaj, A.; Maxwell, D.J. Commissural interneurons with input from group I and II muscle afferents in feline lumbar segments: Neurotransmitters, projections and target cells. J. Physiol. 2009, 587, 401–418. [Google Scholar] [CrossRef]
- Zhang, Y.; Narayan, S.; Geiman, E.; Lanuza, G.M.; Velasquez, T.; Shanks, B.; Akay, T.; Dyck, J.; Pearson, K.; Gosgnach, S.; et al. V3 Spinal Neurons Establish a Robust and Balanced Locomotor Rhythm during Walking. Neuron 2008, 60, 84–96. [Google Scholar] [CrossRef]
- Danner, S.M.; Zhang, H.; Shevtsova, N.A.; Borowska-Fielding, J.; Deska-Gauthier, D.; Rybak, I.A.; Zhang, Y. Spinal V3 Interneurons and Left–Right Coordination in Mammalian Locomotion. Front. Cell. Neurosci. 2019, 13, 516. [Google Scholar] [CrossRef]
- Pierrot-Deseilligny, E.; Burke, D. Group II Pathways. In The Circuitry of the Human Spinal Cord; Cambridge University Press: Cambridge, UK, 2012; pp. 249–291. [Google Scholar]
- Nardone, A.; Schieppati, M. Medium-latency response to muscle stretch in human lower limb: Estimation of conduction velocity of group II fibres and central delay. Neurosci. Lett. 1998, 249, 29–32. [Google Scholar] [CrossRef]
- Marque, P.; Nicolas, G.; Simonetta-Moreau, M.; Pierrot-Deseilligny, E.; Marchand-Pauvert, V. Group II excitations from plantar foot muscles to human leg and thigh motoneurones. Exp. Brain Res. 2005, 161, 486–501. [Google Scholar] [CrossRef]
- Corna, S.; Grasso, M.; Nardone, A.; Schieppati, M. Selective depression of medium-latency leg and foot muscle responses to stretch by an alpha 2-agonist in humans. J. Physiol. 1995, 484, 803–809. [Google Scholar] [CrossRef]
- Bras, H.; Cavallari, P.; Jankowska, E.; McCrea, D. Comparison of effects of monoamines on transmission in spinal pathways from group I and II muscle afferents in the cat. Exp. Brain Res. 1989, 76, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Crone, C.; Nielsen, J. Methodological implications of the post activation depression of the soleus H-reflex in man. Exp. Brain Res. 1989, 78, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Hultborn, H.; Illert, M.; Nielsen, J.; Paul, A.; Ballegaard, M.; Wiese, H. On the mechanism of the post-activation depression of the H-reflex in human subjects. Exp. Brain Res. 1996, 108, 450–462. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, E.; Edgley, S.A. Functional subdivision of feline spinal interneurons in reflex pathways from group Ib and II muscle afferents; an update. Eur. J. Neurosci. 2010, 32, 881–893. [Google Scholar] [CrossRef]
- Jankowska, E.; Slawinska, U.; Hammar, I. On organization of a neuronal network in pathways from group II muscle afferents in feline lumbar spinal segments. J. Physiol. 2002, 542, 301–314. [Google Scholar] [CrossRef]
- Chaix, Y.; Marque, P.; Meunier, S.; Pierrot-Deseilligny, E.; Simonetta-Moreau, M. Further evidence for non-monoynaptic group I excitation of motoneurones in the human lower limb. Exp. Brain Res. 1997, 115, 35–46. [Google Scholar] [CrossRef]
- Pierrot-Deseilligny, E.; Burke, D. Propriospinal Transmission of Descending Motor Commands. In The Circuitry of the Human Spinal Cord; Cambridge University Press: Cambridge, UK, 2012; pp. 395–445. [Google Scholar]
- Shefchyk, S.; McCrea, D.; Kriellaars, D.; Fortier, P.; Jordan, L. Activity of interneurons within the L4 spinal segment of the cat during brainstem-evoked fictive locomotion. Exp. Brain Res. 1990, 80, 290–295. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Freundl, B.; Binder, H.; Minassian, K. Recovery cycles of posterior root-muscle reflexes evoked by transcutaneous spinal cord stimulation and of the H reflex in individuals with intact and injured spinal cord. PLoS ONE 2019, 14, e0227057. [Google Scholar] [CrossRef]
- Kagamihara, Y.; Hayashi, A.; Okuma, Y.; Nagaoka, M.; Nakajima, Y.; Tanaka, R. Reassessment of H-reflex recovery curve using the double stimulation procedure. Muscle Nerve 1998, 21, 352–360. [Google Scholar] [CrossRef]
- Faist, M.; Mazevet, D.; Dietz, V.; Pierrot-Deseilligny, E. A quantitative assessment of presynaptic inhibition of Ia afferents in spastics. Differences in hemiplegics and paraplegics. Brain 1994, 117, 1449–1455. [Google Scholar] [CrossRef]
- Bergmans, J.; Delwaide, P.J.; Gadea-Ciria, M. Short-latency effects of low-threshold muscular afferent fibers on different motoneuronal pools of the lower limb in man. Exp. Neurol. 1978, 60, 380–385. [Google Scholar] [CrossRef]
- Meunier, S.; Pierrot-Deseilligny, E.; Simonetta, M. Pattern of monosynaptic heteronymous Ia connections in the human lower limb. Exp. Brain Res. 1993, 96, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Pierrot-Deseilligny, E.; Burke, D. Monosynaptic Ia Excitation, Homosynaptic Depression and Transcortical Ia Excitation. In The Circuitry of the Human Spinal Cord; Cambridge University Press: Cambridge, UK, 2012; pp. 65–109. [Google Scholar]
- Roby-Brami, A.; Bussel, B. Long-latency spinal reflex in man after flexor reflex afferent stimulation. Brain 1987, 110, 707–725. [Google Scholar] [CrossRef] [PubMed]
- Pierrot-Deseilligny, E.; Burke, D. Cutaneomuscular, Withdrawal and Flexor Reflex Afferent (‘FRA’) Responses. In The Circuitry of the Human Spinal Cord; Cambridge University Press: Cambridge, UK, 2012; pp. 334–394. [Google Scholar]
- Andén, N.-E.; Jukes, M.G.M.; Lundberg, A.; Vyklicky, L. The Effect of DOPA on the Spinal Cord. 1. Influence on Transmission from Primary Aflerents. Acta Physiol. Scand. 1966, 67, 373–386. [Google Scholar] [CrossRef]
- Bussel, B.; Roby-Brami, A.; Rémy Néris, O.; Yakovleff, A. Evidence for a spinal stepping generator in man. Spinal Cord 1996, 34, 91–92. [Google Scholar] [CrossRef]
- Jankowska, E.; Jukes, M.G.; Lund, S.; Lundberg, A. The effect of DOPA on the spinal cord. 6. Half-centre organization of interneurones transmitting effects from the flexor reflex afferents. Acta Physiol. Scand. 1967, 70, 389–402. [Google Scholar] [CrossRef]
- Lundberg, A. Multisensory control of spinal reflex pathways. Prog. Brain Res. 1979, 50, 11–28. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject No. | Gender | Age (y) 1 | AIS Grade | Neurol. Level of SCI | SCI Chronicity (y) 1 | No. of Recordings Analyzed | ||
|---|---|---|---|---|---|---|---|---|
| Lateral EES | Medial EES | Frequency Study | ||||||
| 1 | m | 18.0 | A | C4 | 2.9 | 0 | 3 | 0 |
| 2 | m | 25.6 | A | C4 | 3.8 | 0 | 3 | 0 |
| 3 | m | 21.9 | A | C7 | 5.1 | 0 | 3 | 1 |
| 4 | m | 21.3 | A | T7 | 2.6 | 3 | 2 | 0 |
| 5 | f | 24.7 | A | T7 | 4.2 | 0 | 2 | 0 |
| 6 | m | 33.2 | A | T8 | 13.5 | 0 | 3 | 1 |
| 7 | m | 29.0 | B | C6 | 2.5 | 1 | 3 | 1 |
| 8 | m | 25.3 | B | C8 | 1.6 | 0 | 4 | 1 |
| Subject | 4 | 7 | |||
|---|---|---|---|---|---|
| Recording | 1 | 2 | 3 | 1 | |
| EES side | right | left | left | right | right |
| Electrode setup | 2+3− | 2+3− | 1+3− | 1+3− | 1−3+ |
| Max. EES amp. (V) | 8 | 10 | 9 | 10 | 6 |
| Thresholds of Monosynaptic PRM Reflexes in RF and TS | |||||
| Thr iRF (0–50 ms) | 4 | 3 | 1 | 3 | 2 |
| Thr iTS (0–50 ms) | NaN | 8 | 3 | 7 | 5 |
| Thr cRF (0–50 ms) | NaN | NaN | NaN | NaN | 6 |
| Thr cTS (0–50 ms) | NaN | NaN | NaN | NaN | NaN |
| Thresholds of Reflexes in Ham | |||||
| Thr iHam (0–50 ms) | 4 | 5 | 2 | 4 | 2 |
| Thr cHam (0–50 ms) | NaN | 8 | 6 | NaN | 7 |
| Thr xHam (50–100 ms) | 3 | 6 | 5 | 4 | 2 |
| Response Group | Onset Latency | Offset Latency | Q3 Latency | No. Peaks |
|---|---|---|---|---|
| 1 | 19.61 ± 0.05 ms | 31.56 ± 0.19 ms | 28.55 ± 0.07 ms | 2.52 ± 0.02 |
| 2 | 19.13 ± 0.09 ms | 29.84 ± 0.30 ms | 28.14 ± 0.15 ms | 2.57 ± 0.04 |
| 3 | 18.91 ± 0.09 ms | 35.31 ± 0.53 ms | 33.57 ± 0.22 ms | 3.47 ± 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofstoetter, U.S.; Danner, S.M.; Freundl, B.; Binder, H.; Lackner, P.; Minassian, K. Ipsi- and Contralateral Oligo- and Polysynaptic Reflexes in Humans Revealed by Low-Frequency Epidural Electrical Stimulation of the Lumbar Spinal Cord. Brain Sci. 2021, 11, 112. https://doi.org/10.3390/brainsci11010112
Hofstoetter US, Danner SM, Freundl B, Binder H, Lackner P, Minassian K. Ipsi- and Contralateral Oligo- and Polysynaptic Reflexes in Humans Revealed by Low-Frequency Epidural Electrical Stimulation of the Lumbar Spinal Cord. Brain Sciences. 2021; 11(1):112. https://doi.org/10.3390/brainsci11010112
Chicago/Turabian StyleHofstoetter, Ursula S., Simon M. Danner, Brigitta Freundl, Heinrich Binder, Peter Lackner, and Karen Minassian. 2021. "Ipsi- and Contralateral Oligo- and Polysynaptic Reflexes in Humans Revealed by Low-Frequency Epidural Electrical Stimulation of the Lumbar Spinal Cord" Brain Sciences 11, no. 1: 112. https://doi.org/10.3390/brainsci11010112
APA StyleHofstoetter, U. S., Danner, S. M., Freundl, B., Binder, H., Lackner, P., & Minassian, K. (2021). Ipsi- and Contralateral Oligo- and Polysynaptic Reflexes in Humans Revealed by Low-Frequency Epidural Electrical Stimulation of the Lumbar Spinal Cord. Brain Sciences, 11(1), 112. https://doi.org/10.3390/brainsci11010112