Effects of Lifelong Musicianship on White Matter Integrity and Cognitive Brain Reserve

 ,
,
Abstract
:1. Introduction
1.1. Cognitive Reserve in Bilinguals
1.2. Cognitive Reserve in Musicians
1.3. Research Question and Hypotheses
2. Materials and Methods
2.1. Participants
2.2. Data
2.2.1. Behavioral Data
2.2.2. Image Acquisition
2.2.3. DTI Processing
2.2.4. Statistical Methods
3. Results
4. Discussion
Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bialystok, E.; Craik, F.I.M.; Freedman, M. Bilingualism as a protection against the onset of symptoms of dementia. Neuropsychologia 2007, 45, 459–464. [Google Scholar] [CrossRef]
- Valenzuela, M.J.; Sachdev, P. Brain reserve and dementia: A systematic review. Psychol. Med. 2006, 36, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Stern, Y. Cognitive reserve in ageing and Alzheimer’s disease. Lancet Neurol. 2012, 11, 1006–1012. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Vélez, E.; Tranel, D. Does bilingualism contribute to cognitive reserve? Cognitive and neural perspectives. Neuropsychology 2015, 29, 139–150. [Google Scholar] [CrossRef]
- Gold, B.T.; Johnson, N.F.; Powell, D.K. Lifelong bilingualism contributes to cognitive reserve against white matter integrity declines in aging. Neuropsychologia 2013, 51, 2841–2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craik, F.I.M.; Bialystok, E.; Freedman, M. Delaying the onset of Alzheimer disease Bilingualism as a form of cognitive reserve. Neurology 2010, 75, 1726–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, E.; Frigau, L.; Voyvodic-Casabo, C.; Voyvodic, J.; Wright, J. Multilingualism and fMRI: Longitudinal Study of Second Language Acquisition. Brain Sci. 2013, 3, 849–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, E. Neuroscience and Multilingualism; Cambridge University Press: Cambridge, UK, 2014; p. 254. [Google Scholar]
- De Bot, K. Multilingualism and aging. In The New Handbook of Second Language Acquisition; Bhatia, T.K., Ritchie, W.C., Eds.; Emerald Group Publishing: Bingley, UK, 2009; pp. 425–442. [Google Scholar]
- Pliatsikas, C.; Moschopoulou, E.; Saddy, J.D. The effects of bilingualism on the white matter structure of the brain. Proc. Natl. Acad. Sci. USA 2015, 112, 1334–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luk, G.; Bialystok, E.; Craik, F.I.M.; Grady, C.L. Lifelong Bilingualism Maintains White Matter Integrity in Older Adults. J. Neurosci. 2011, 31, 16808–16813. [Google Scholar] [CrossRef]
- Anderson, J.A.E.; Grundy, J.G.; De Frutos, J.; Barker, R.M.; Grady, C.; Bialystok, E. Effects of bilingualism on white matter integrity in older adults. Neuroimage 2018, 167, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Coggins, P.E.; Kennedy, T.J.; Armstrong, T.A. Bilingual corpus callosum variability. Brain Lang. 2004, 89, 69–75. [Google Scholar] [CrossRef]
- Felton, A.; Vazquez, D.; Ramos-Nunez, A.I.; Greene, M.R.; Macbeth, A.; Hernandez, A.E.; Chiarello, C. Bilingualism influences structural indices of interhemispheric organization. J. Neurolinguist. 2017, 42, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, S.; Oishi, K.; Jiang, H.; Jiang, L.; Li, X.; Akhter, K.; Hua, K.; Faria, A.V.; Mahmood, A.; Woods, R.; et al. Stereotaxic white matter atlas based on diffusion tensor imaging in an ICBM template. Neuroimage 2008, 40, 570–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, K.; Zilles, K.; Amunts, K.; Faria, A.; Jiang, H.; Li, X.; Akhter, K.; Hua, K.; Woods, R.; Toga, A.W.; et al. Human brain white matter atlas: Identification and assignment of common anatomical structures in superficial white matter. Neuroimage 2008, 43, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, J.G.; Anderson, J.A.; Bialystok, E. Neural correlates of cognitive processing in monolinguals and bilinguals. Ann. N. Y. Acad. Sci. 2017, 1396, 183–201. [Google Scholar] [CrossRef] [Green Version]
- Abutalebi, J.; Canini, M.; Della Rosa, P.A.; Green, D.W.; Weekes, B.S. The neuroprotective effects of bilingualism upon the inferior parietal lobule: A Structural Neuroimaging Study in Aging Chinese Bilinguals. J. Neurolinguist. 2015, 33, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Abutalebi, J.; Guidi, L.; Borsa, V.; Canini, M.; Della Rosa, P.A.; Parris, B.A.; Weekes, B.S. Bilingualism provides a neural reserve for aging populations. Neuropsychologia 2015, 69, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Elmer, S.; Jancke, L. Relationships between music training, speech processing, and word learning: A network perspective. Ann. N. Y. Acad. Sci. 2018, 1423, 10–18. [Google Scholar] [CrossRef]
- Elmer, S.; Hanggi, J.; Vaquero, L.; Cadena, G.O.; FranCois, C.; Rodriguez-Fornells, A. Tracking the microstructural properties of the main white matter pathways underlying speech processing in simultaneous interpreters. Neuroimage 2019, 191, 518–528. [Google Scholar] [CrossRef]
- Jäncke, L. The plastic human brain. Restor. Neurol. Neurosci. 2009, 27, 521–538. [Google Scholar] [CrossRef]
- Munte, T.F.; Altenmuller, E.; Jancke, L. The musician’s brain as a model of neuroplasticity. Nat. Rev. Neurosci. 2002, 3, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, P.; Lerch, J.P.; Evans, A.C.; Zatorre, R.J. Neuroanatomical Correlates of Musicianship as Revealed by Cortical Thickness and Voxel-Based Morphometry. Cereb. Cortex 2009, 19, 1583–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhnis, J.; Elmer, S.; Meyer, M.; Jancke, L. The encoding of vowels and temporal speech cues in the auditory cortex of professional musicians: An EEG study. Neuropsychologia 2013, 51, 1608–1618. [Google Scholar] [CrossRef]
- Pantev, C.; Roberts, L.E.; Schulz, M.; Engelien, A.; Ross, B. Timbre-specific enhancement of auditory cortical representations in musicians. Neuroreport 2001, 12, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangert, M.; Peschel, T.; Schlaug, G.; Rotte, M.; Drescher, D.; Hinrichs, H.; Heinze, H.J.; Altenmuller, E. Shared networks for auditory and motor processing in professional pianists: Evidence from fMRI conjunction. Neuroimage 2006, 30, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.W.; Bidelman, G.M. Listening to the Brainstem: Musicianship Enhances Intelligibility of Subcortical Representations for Speech. J. Neurosci. 2015, 35, 1687–1691. [Google Scholar] [CrossRef] [Green Version]
- Halwani, G.F.; Loui, P.; Ruber, T.; Schlaug, G. Effects of practice and experience on the arcuate fasciculus: Comparing singers, instrumentalists, and non-musicians. Front. Psychol. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Oechslin, M.S.; Imfeld, A.; Loenneker, T.; Meyer, M.; Jancke, L. The plasticity of the superior longitudinal fasciculus as a function of musical expertise: A diffusion tensor imaging study. Front. Hum. Neurosci. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Yuskaitis, C.J.; Parviz, M.; Loui, P.; Wan, C.Y.; Pearl, P.L. Neural Mechanisms Underlying Musical Pitch Perception and Clinical Applications Including Developmental Dyslexia. Curr. Neurol. Neurosci. Rep. 2015, 15, 51. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.Y.; Schlaug, G. Neural pathways for language in autism: The potential for music-based treatments. Future Neurol. 2010, 5, 797–805. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Nyengaard, J.R.; Pakkenberg, B.; Gundersen, H.J.G. Age-induced white matter changes in the human brain: A stereological investigation. Neurobiol. Aging 1997, 18, 609–615. [Google Scholar] [CrossRef]
- Giorgio, A.; Santelli, L.; Tomassini, V.; Bosnell, R.; Smith, S.; De Stefano, N.; Johansen-Berg, H. Age-related changes in grey and white matter structure throughout adulthood. Neuroimage 2010, 51, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westlye, L.T.; Walhovd, K.B.; Dale, A.M.; Bjornerud, A.; Due-Tonnessen, P.; Engvig, A.; Grydeland, H.; Tamnes, C.K.; Ostby, Y.; Fjell, A.M. Life-Span Changes of the Human Brain White Matter: Diffusion Tensor Imaging (DTI) and Volumetry. Cereb. Cortex 2010, 20, 2055–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billiet, T.; Vandenbulcke, M.; Madler, B.; Peeters, R.; Dhollander, T.; Zhang, H.; Deprez, S.; Van den Bergh, B.R.H.; Sunaert, S.; Emsell, L. Age-related microstructural differences quantified using myelin water imaging and advanced diffusion MRI. Neurobiol. Aging 2015, 36, 2107–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathee, R.; Rallabandi, V.P.S.; Roy, P.K. Age-Related Differences in White Matter Integrity in Healthy Human Brain: Evidence from Structural MRI and Diffusion Tensor Imaging. Magn. Reson. Insights 2016, 9, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierpaoli, C.; Walker, L.; Irfanoglu, M.O.; Barnett, A.; Basser, P.; Chang, L.-C.; Koay, C.; Pajevic, S.; Rohde, G.; Sarlls, J.; et al. TORTOISE: An integrated software package for processing of diffusion MRI data. In Proceedings of the ISMRM 18th Annual Meeting, Stockholm, Sweden, 5 May 2010. abstract #1597. [Google Scholar]
- Irfanoglu, M.O.; Nayak, A.; Jenkins, J.; Pierpaoli, P. TORTOISE v3: Improvements and New Features of the NIH Diffusion MRI Processing Pipeline. In Proceedings of the ISMRM 25th Annual Meeting, Honolulu, HI, USA, 25 April 2017. abstract #3540. [Google Scholar]
- Woolrich, M.W.; Jbabdi, S.; Patenaude, B.; Chappell, M.; Makni, S.; Behrens, T.; Beckmann, C.; Jenkinson, M.; Smith, S.M. Bayesian analysis of neuroimaging data in FSL. Neuroimage 2009, 45, S173–S186. [Google Scholar] [CrossRef]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; De Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. Neuroimage 2004, 23 (Suppl. 1), S208–S219. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.; Woolrich, M.W.; Smith, S.M. Fsl. Neuroimage 2012, 62, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Fisher, R.A. Statistical Methods for Research Workers; Oliver and Boyd: Edinburgh, UK, 1925. [Google Scholar]
- Friederici, A.D. Pathways to language: Fiber tracts in the human brain. Trends Cogn. Sci 2009, 13, 175–181. [Google Scholar] [CrossRef]
- Madhavan, K.M.; McQueeny, T.; Howe, S.R.; Shear, P.; Szaflarski, J. Superior longitudinal fasciculus and language functioning in healthy aging. Brain Res. 2014, 1562, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Riley, J.D.; Franklin, D.L.; Choi, V.; Kim, R.C.; Binder, D.K.; Cramer, S.C.; Lin, J.J. Altered white matter integrity in temporal lobe epilepsy: Association with cognitive and clinical profiles. Epilepsia 2010, 51, 536–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, B.; Busch, R.M.; Duncan, J.S.; Piao, Z.; Tkach, J.; Luders, H.O. Abnormalities in diffusion tensor imaging of the uncinate fasciculus relate to reduced memory in temporal lobe epilepsy. Epilepsia 2008, 49, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- McDonald, C.R.; Ahmadi, M.E.; Hagler, D.J.; Tecoma, E.S.; Iragui, V.J.; Gharapetian, L.; Dale, A.M.; Halgren, E. Diffusion tensor imaging correlates of memory and language impairments in temporal lobe epilepsy. Neurology 2008, 71, 1869–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.H.; Crosson, B.; Nadeau, S.E.; Heilman, K.M.; Gonzalez-Rothi, L.J.; Raymer, A.; Gilmore, R.L.; Bauer, R.M.; Roper, S.N. Category-specific naming deficits for objects and actions: Semantic attribute and grammatical role hypotheses. Neuropsychologia 2002, 40, 1608–1621. [Google Scholar] [CrossRef]
- Hamberger, M.J.; Drake, E.B. Cognitive functioning following epilepsy surgery. Curr. Neurol. Neurosci. Rep. 2006, 6, 319–326. [Google Scholar] [CrossRef]
- Grabowski, T.J.; Damasio, H.; Tranel, D.; Ponto, L.L.; Hichwa, R.D.; Damasio, A.R. A role for left temporal pole in the retrieval of words for unique entities. Hum. Brain Mapp. 2001, 13, 199–212. [Google Scholar] [CrossRef]
- Craig, M.C.; Catani, M.; Deeley, Q.; Latham, R.; Daly, E.; Kanaan, R.; Picchioni, M.; McGuire, P.K.; Fahy, T.; Murphy, D.G. Altered connections on the road to psychopathy. Mol. Psychiatry 2009, 14, 907, 946–953. [Google Scholar] [CrossRef]
- Duffau, H.; Gatignol, P.; Moritz-Gasser, S.; Mandonnet, E. Is the left uncinate fasciculus essential for language? A cerebral stimulation study. J. Neurol. 2009, 256, 382–389. [Google Scholar] [CrossRef]
- Krzyżak, A.T.; Olejniczak, Z. Improving the accuracy of PGSE DTI experiments using the spatial distribution of b matrix. Magn. Reson. Imaging 2015, 33, 286–295. [Google Scholar] [CrossRef]
- Borkowski, K.; Krzyżak, A.T. Analysis and correction of errors in DTI-based tractography due to diffusion gradient inhomogeneity. J. Magn. Reson. 2018, 296, 5–11. [Google Scholar] [CrossRef]
- Helmer, K.G.; Chou, M.C.; Preciado, R.I.; Gimi, B.; Rollins, N.K.; Song, A.; Turner, J.; Mori., S. Multi-site study of diffusion metric variability: Characterizing the effects of site, vendor, field strength, and echo time using the histogram distance. In Proceedings of the SPIE, San Diego, CA, USA, 29 March 2016; p. 9788. [Google Scholar] [CrossRef] [Green Version]
- Kochunov, P.; Thompson, P.M.; Lancaster, J.L.; Bartzokis, G.; Smith, S.; Coyle, T.l.; Royall, D.R.; Laird, A.; Fox, P.T. Relationship between white matter fractional anisotropy and other indices of cerebral health in normal aging: Tract-based spatial statistics study of aging. NeuroImage 2007, 35, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Kochunov, P.; Glahn, D.C.; Lancaster, J.; Thompson, P.M.; Kochunov, V.; Rogers, B.; Fox, P.; Blangero, J.; Williamson, D.E. Fractional anisotropy of cerebral white matter and thickness of cortical gray matter across the lifespan. Neuroimage 2011, 58, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, I.J.; Motes, M.A.; Rao, N.K.; Rypma, B. White matter tract integrity predicts visual search performance in young and older adults. Neurobiol. Aging 2012, 33, 433–e421. [Google Scholar] [CrossRef] [Green Version]
- Li, J.F.; Luo, C.; Peng, Y.H.; Xie, Q.K.; Gong, J.N.; Dong, L.; Lai, Y.X.; Li, H.; Yao, D.Z. Probabilistic Diffusion Tractography Reveals Improvement of Structural Network in Musicians. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]



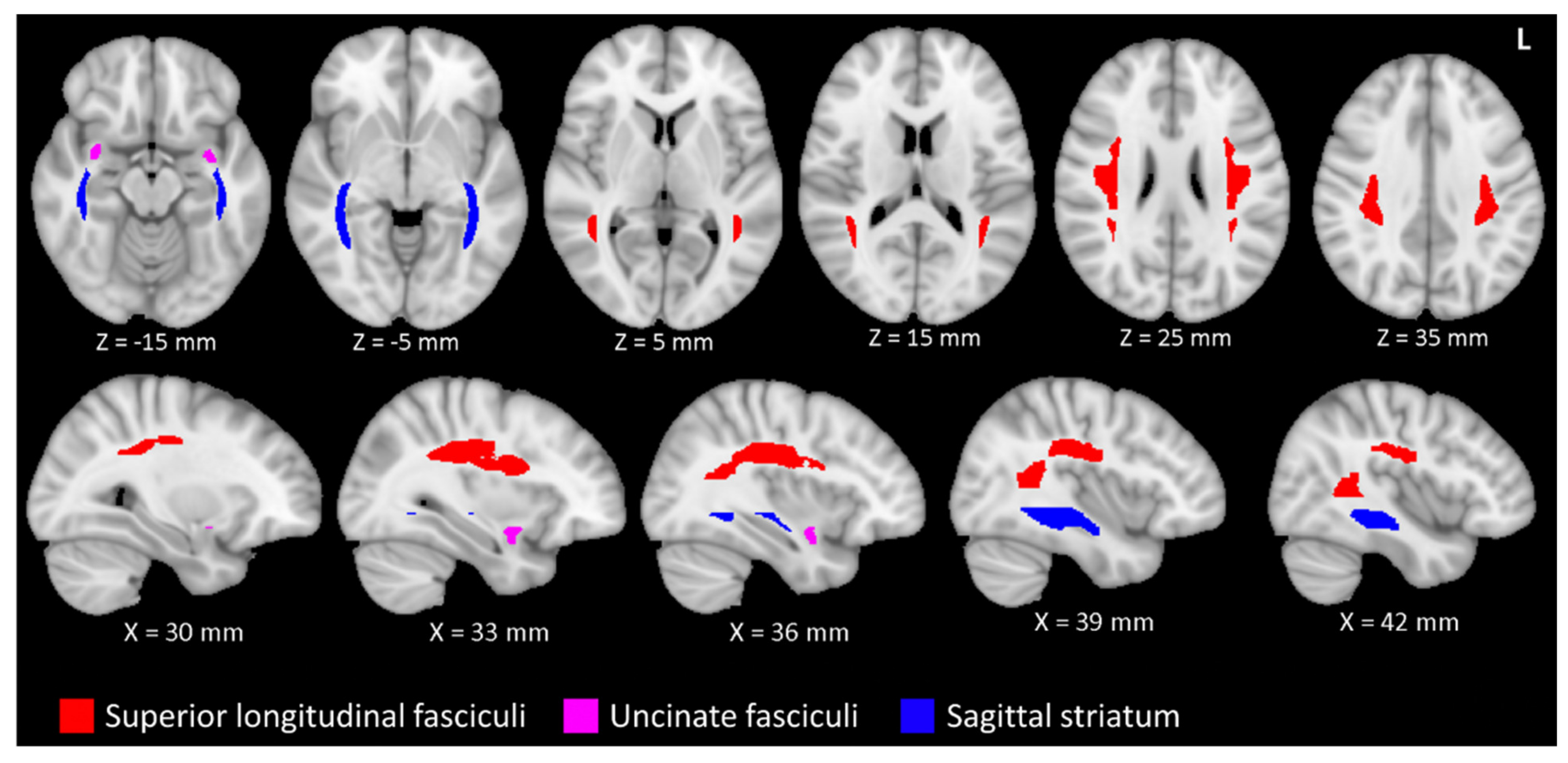{kind=link}
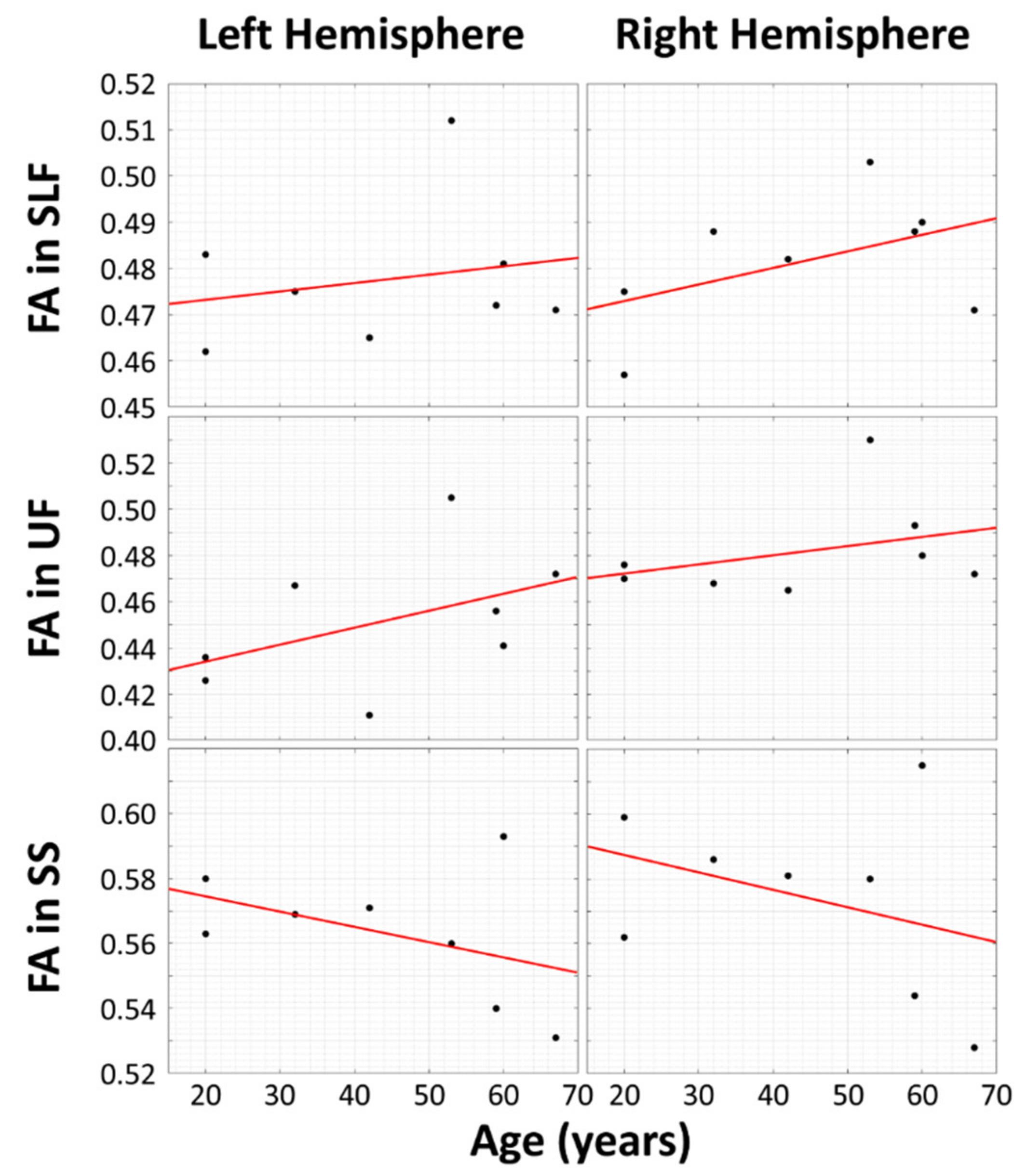{kind=link}
| Subject | LSLF | RSLF | LUF | RUF | LSS | RSS |
|---|---|---|---|---|---|---|
| 1 | 0.471 (0.14) | 0.471 (0.13) | 0.472 (0.12) | 0.472 (0.10) | 0.531 (0.12) | 0.528 (0.10) |
| 2 | 0.512 (0.14) | 0.503 (0.13) | 0.505 (0.15) | 0.530 (0.14) | 0.560 (0.10) | 0.580 (0.09) |
| 3 | 0.472 (0.14) | 0.488 (0.13) | 0.456 (0.13) | 0.493 (0.11) | 0.540 (0.11) | 0.544 (0.09) |
| 4 | 0.462 (0.14) | 0.457 (0.13) | 0.426 (0.13) | 0.470 (0.12) | 0.580 (0.12) | 0.562 (0.10) |
| 5 | 0.465 (0.12) | 0.482 (0.11) | 0.411 (0.12) | 0.465 (0.10) | 0.571 (0.10) | 0.581 (0.10) |
| 6 | 0.481 (0.12) | 0.490 (0.12) | 0.441 (0.15) | 0.480 (0.14) | 0.593 (0.11) | 0.615 (0.10) |
| 7 | 0.475 (0.13) | 0.488 (0.13) | 0.467 (0.13) | 0.468 (0.12) | 0.569 (0.09) | 0.586 (0.09) |
| 8 | 0.483 (0.12) | 0.475 (0.12) | 0.436 (0.11) | 0.476 (0.12) | 0.563 (0.10) | 0.599 (0.10) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrews, E.; Eierud, C.; Banks, D.; Harshbarger, T.; Michael, A.; Rammell, C. Effects of Lifelong Musicianship on White Matter Integrity and Cognitive Brain Reserve. Brain Sci. 2021, 11, 67. https://doi.org/10.3390/brainsci11010067
Andrews E, Eierud C, Banks D, Harshbarger T, Michael A, Rammell C. Effects of Lifelong Musicianship on White Matter Integrity and Cognitive Brain Reserve. Brain Sciences. 2021; 11(1):67. https://doi.org/10.3390/brainsci11010067
Chicago/Turabian StyleAndrews, Edna, Cyrus Eierud, David Banks, Todd Harshbarger, Andrew Michael, and Charlotte Rammell. 2021. "Effects of Lifelong Musicianship on White Matter Integrity and Cognitive Brain Reserve" Brain Sciences 11, no. 1: 67. https://doi.org/10.3390/brainsci11010067
APA StyleAndrews, E., Eierud, C., Banks, D., Harshbarger, T., Michael, A., & Rammell, C. (2021). Effects of Lifelong Musicianship on White Matter Integrity and Cognitive Brain Reserve. Brain Sciences, 11(1), 67. https://doi.org/10.3390/brainsci11010067