Aerobic Exercise Prevents Depression via Alleviating Hippocampus Injury in Chronic Stressed Depression Rats



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Protocal of Aerobic Exercise
2.3. Behavioral Tests
2.3.1. Open Field Test
2.3.2. Forced Swimming Test
2.4. Assessment of Hippocampal Neuronal Apoptosis
2.5. ELISA Analysis
2.6. Statistical Analyses
3. Results
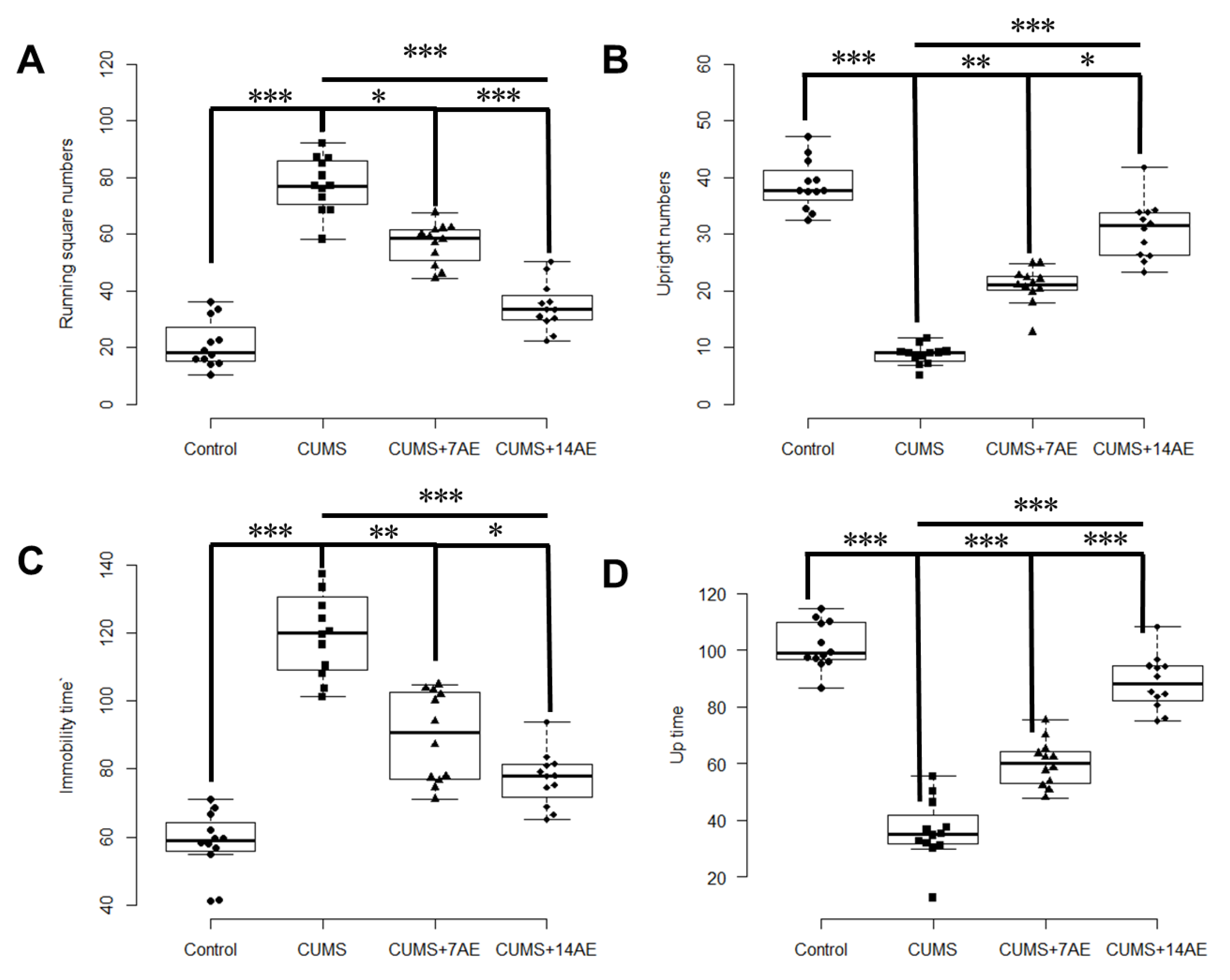
3.1. AE Improved Behavioral Indexes in CUMS-Induced Rats
3.2. AE Reduced CUMS-Induced Hippocampal Neuronal Apoptosis
3.3. AE Beneficially Regulated Levels of Ionotropic Glutamate Receptors in CUMS-Induced Depression Rats
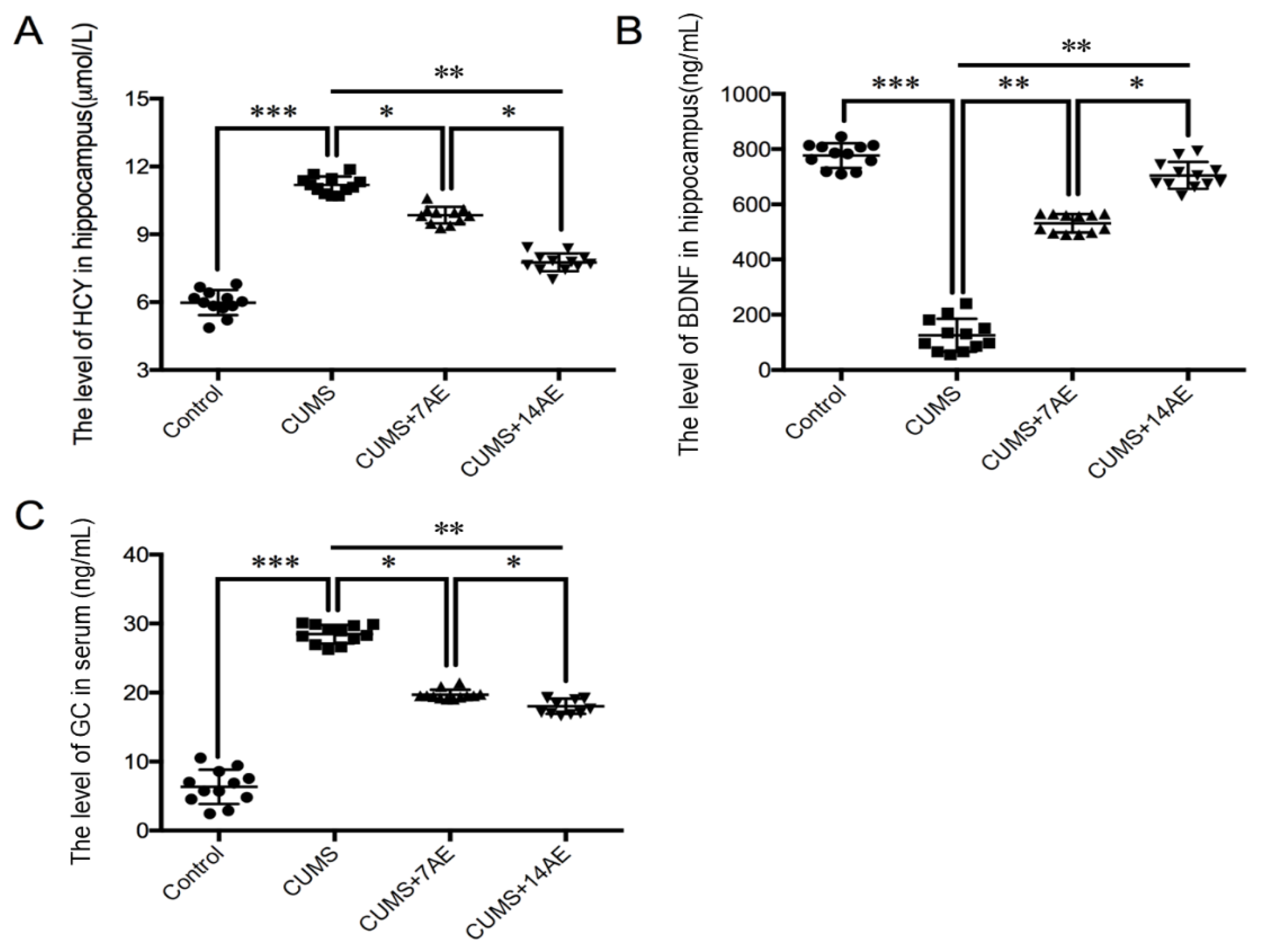
3.4. AE Beneficially Regulated Levels of HCY, BDNF and GC in CUMS-Induced Depression Rats
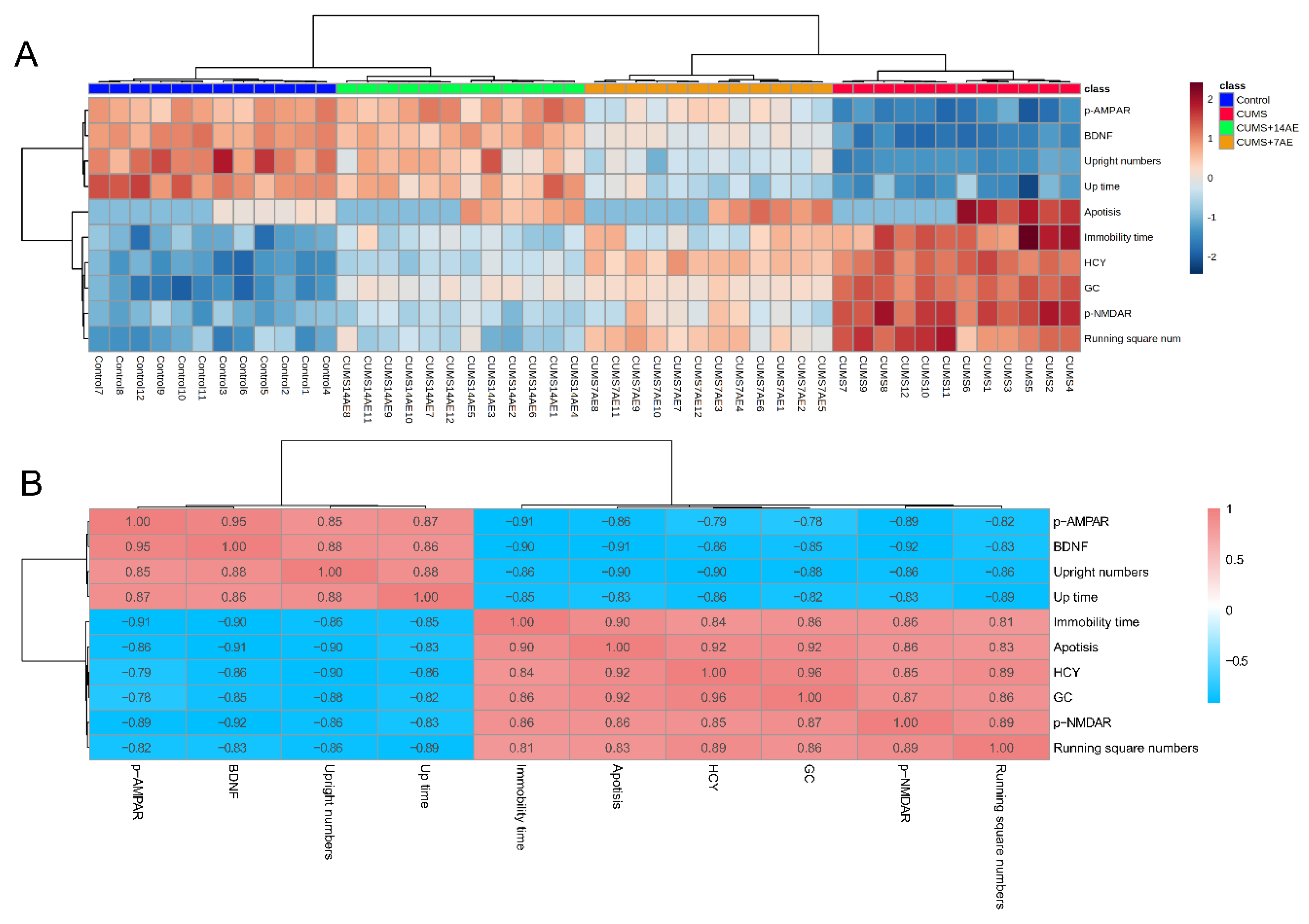
3.5. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ferrari, A.J.; Charlson, F.J.; Norman, R.E.; Patten, S.B.; Freedman, G.D.; Murray, C.J.; Vos, T.; Whiteford, H.A. Burden of Depressive Disorders by Country, Sex, Age, and Year: Findings from the Global Burden of Disease Study 2010. PLoS Med. 2013, 10, e1001547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuckit, M.A.; Irwin, M.R.; Smith, T.L. One-year incidence rate of major depression and other psychiatric disorders in 239 alcoholic men. Addiction 1994, 89, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Lucassen, P.J.; Meerlo, P.; Naylor, A.S.; Van Dam, A.M.; Dayer, A.G.; Fuchs, E.; Oomen, C.A.; Czéh, B. Regulation of adult neurogenesis by stress, sleep disruption, exercise and inflammation: Implications for depression and antidepressant action. Eur. Neuropsychopharmacol. 2010, 20, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Tang, J.; Chao, F.-L.; Zhang, Y.; Chen, L.-M.; Wang, F.-F.; Tan, C.-X.; Luo, Y.-M.; Xiao, Q.; Zhang, L.; et al. Exercise improves depressive symptoms by increasing the number of excitatory synapses in the hippocampus of CUS-Induced depression model rats. Behav. Brain Res. 2019, 374, 112115. [Google Scholar] [CrossRef]
- Sheline, Y.I. Depression and the Hippocampus: Cause or Effect? Biol. Psychiatry 2011, 70, 308–309. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.-W.; Shiue, Y.-L.; Liao, J.-C.; Wee, H.-Y.; Wang, C.-C.; Chio, C.-C.; Chang, C.-H.; Hu, C.-Y.; Kuo, J.-R. Simvastatin Therapy in the Acute Stage of Traumatic Brain Injury Attenuates Brain Trauma-Induced Depression-Like Behavior in Rats by Reducing Neuroinflammation in the Hippocampus. Neurocritical Care 2016, 26, 122–132. [Google Scholar] [CrossRef]
- Salmon, P. Effects of physical exercise on anxiety, depression, and sensitivity to stress: A unifying theory. Clin. Psychol. Rev. 2001, 21, 33–61. [Google Scholar] [CrossRef]
- Farris, S.G.; Abrantes, A.M.; Uebelacker, L.A.; Weinstock, L.M.; Battle, C.L. Exercise as a Nonpharmacological Treatment for Depression. Psychiatr. Ann. 2019, 49, 6–10. [Google Scholar] [CrossRef]
- Busch, A.M.; Ciccolo, J.T.; Puspitasari, A.J.; Nosrat, S.; Whitworth, J.W.; Stults-Kolehmainen, M.A. Preferences for exercise as a treatment for depression. Ment. Health Phys. Act. 2016, 10, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Harvey, S.; Øverland, S.; Hatch, S.L.; Wessely, S.; Mykletun, A.; Hotopf, M. Exercise and the Prevention of Depression: Results of the HUNT Cohort Study. Am. J. Psychiatry 2018, 175, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Tang, C.; Chen, X.; Ren, Z.; Qu, H.; Chen, R.; Tong, Z. Impacts of Aerobic Exercise on Depression-Like Behaviors in Chronic Unpredictable Mild Stress Mice and Related Factors in the AMPK/PGC-1α Pathway. Int. J. Environ. Res. Public Health 2020, 17, 2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.-H.; Park, H.-S.; Park, S.-S.; Kim, C.-J.; Kim, D.-H.; Kim, T.-W. Physical exercise ameliorates psychiatric disorders and cognitive dysfunctions by hippocampal mitochondrial function and neuroplasticity in post-traumatic stress disorder. Exp. Neurol. 2019, 322, 113043. [Google Scholar] [CrossRef]
- Morgan, J.A.; Singhal, G.; Corrigan, F.; Jaehne, E.J.; Jawahar, M.C.; Baune, B.T. Exercise related anxiety-like behaviours are mediated by TNF receptor signaling, but not depression-like behaviours. Brain Res. 2018, 1695, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Zhan, Q.; Xiao, W.; Tang, X.; Li, J.; Dong, H.; Sha, W.; Zhang, X. Altered serum levels of vascular endothelial growth factor in first-episode drug-naïve and chronic medicated schizophrenia. Psychiatry Res. 2018, 264, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Wang, Y.; Wang, D. Endurance and resistance training mitigate the negative consequences of depression on synaptic plasticity through different molecular mechanisms. Int. J. Neurosci. 2019, 130, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; Dileone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of Depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Zhao, Y.-Q.; Ding, X.-F.; Wu, Y.; Qu, W. Quinacrine pretreatment reduces microwave-induced neuronal damage by stabilizing the cell membrane. Neural Regen. Res. 2018, 13, 449–455. [Google Scholar] [CrossRef]
- Mitchell, D.M.; Lovel, A.G.; Stenkamp, D.L. Dynamic changes in microglial and macrophage characteristics during degeneration and regeneration of the zebrafish retina. J. Neuroinflammation 2018, 15, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Wagner, W.; Lippmann, K.; Heisler, F.F.; Gromova, K.V.; Lombino, F.L.; Roesler, M.K.; Pechmann, Y.; Hornig, S.; Schweizer, M.; Polo, S.; et al. Myosin VI Drives Clathrin-Mediated AMPA Receptor Endocytosis to Facilitate Cerebellar Long-Term Depression. Cell Rep. 2019, 28, 11–20.e9. [Google Scholar] [CrossRef] [Green Version]
- Treccani, G.; Du Jardin, K.G.; Wegener, G.; Müller, H.K. Differential expression of postsynaptic NMDA and AMPA receptor subunits in the hippocampus and prefrontal cortex of the flinders sensitive line rat model of depression. Synapse 2016, 70, 471–474. [Google Scholar] [CrossRef]
- Sucher, N.J.; Awobuluyi, M.; Choi, Y.B.; Lipton, S.A. NMDA receptors: From genes to channels. Trends Pharmacol. Sci. 1996, 17, 348–355. [Google Scholar] [CrossRef]
- Forti, P.; Rietti, E.; Pisacane, N.; Olivelli, V.; Dalmonte, E.; Mecocci, P.; Ravaglia, G. Blood homocysteine and risk of depression in the elderly. Arch. Gerontol. Geriatr. 2010, 51, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Pascoe, M.; Crewther, S.G.; Carey, L.M.; Noonan, K.; Crewther, D.P.; Linden, T. Homocysteine as a potential biochemical marker for depression in elderly stroke survivors. Food Nutr. Res. 2012, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-S.; Chou, M.-C.; Yeh, Y.-C.; Yang, Y.-H.; Lai, C.-L.; Yen, C.-F.; Liu, C.-K.; Liao, Y.-C. Plasma Homocysteine Levels and Major Depressive Disorders in Alzheimer Disease. Am. J. Geriatr. Psychiatry 2010, 18, 1045–1048. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Wang, J.; Yan, H.; Zhao, Y.X.; Liu, X.Y. A proteomics study of hyperhomocysteinemia injury of the hippocampal neurons using iTRAQ. Mol. Med. Rep. 2014, 10, 2511–2516. [Google Scholar] [CrossRef] [Green Version]
- Choe, Y.M.; Sohn, B.K.; Choi, H.J.; Byun, M.S.; Seo, E.H.; Han, J.Y.; Kim, Y.K.; Yoon, E.J.; Lee, J.-M.; Park, J.; et al. Association of homocysteine with hippocampal volume independent of cerebral amyloid and vascular burden. Neurobiol. Aging 2014, 35, 1519–1525. [Google Scholar] [CrossRef]
- Poddar, R.; Chen, A.; Winter, L.; Rajagopal, S.; Paul, S. Role of AMPA receptors in homocysteine-NMDA receptor-induced crosstalk between ERK and p38 MAPK. J. Neurochem. 2017, 142, 560–573. [Google Scholar] [CrossRef] [Green Version]
- Suri, D.; Vaidya, V. Glucocorticoid regulation of brain-derived neurotrophic factor: Relevance to hippocampal structural and functional plasticity. Neuroscience 2013, 239, 196–213. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, J.; Deng, M.; Liu, Y.; Hu, Y.; Zhang, L. The Antidepressant Effect of Angelica sinensis Extracts on Chronic Unpredictable Mild Stress-Induced Depression Is Mediated via the Upregulation of the BDNF Signaling Pathway in Rats. Evid. Based Complement. Altern. Med. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fahey, B.; Barlow, S.; Day, J.S.; O’Mara, S.M. Interferon-α-induced deficits in novel object recognition are rescued by chronic exercise. Physiol. Behav. 2008, 95, 125–129. [Google Scholar] [CrossRef]
- Arteni, N.; Pereira, L.; Rodrigues, A.; Lavinsky, D.; Achaval, M.; Netto, C.A. Lateralized and sex-dependent behavioral and morphological effects of unilateral neonatal cerebral hypoxia-ischemia in the rat. Behav. Brain Res. 2010, 210, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Slattery, D.A.; Cryan, J.F. Using the rat forced swim test to assess antidepressant-like activity in rodents. Nat. Protoc. 2012, 7, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Seibenhener, M.L.; Wooten, M.W. Isolation and Culture of Hippocampal Neurons from Prenatal Mice. J. Vis. Exp. 2012, 2012, e3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Wang, L.; Wang, S.; Li, S.; Li, Y.; Zhang, L. Effects of calcium-sensing receptors on apoptosis in rat hippocampus during hypoxia/reoxygenation through the ERK1/2 pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 10808–10815. [Google Scholar]
- Sun, W.; Chen, L.; Zheng, W.; Wei, X.; Wu, W.; Duysen, E.G.; Jiang, W. Study of acetylcholinesterase activity and apoptosis in SH-SY5Y cells and mice exposed to ethanol. Toxicology 2017, 384, 33–39. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, Z.; Liu, Y.; Zhang, R.; Mo, H.; Gao, L.; Shi, Y. Physical exercise rectifies CUMS-induced aberrant regional homogeneity in mice accompanied by the adjustment of skeletal muscle PGC-1a/IDO1 signals and hippocampal function. Behav. Brain Res. 2020, 383, 112516. [Google Scholar] [CrossRef]
- Olson, R.L.; Brush, C.J.; Ehmann, P.J.; Alderman, B.L. A randomized trial of aerobic exercise on cognitive control in major depression. Clin. Neurophysiol. 2017, 128, 903–913. [Google Scholar] [CrossRef]
- Gulyaeva, N.V. Functional Neurochemistry of the Ventral and Dorsal Hippocampus: Stress, Depression, Dementia and Remote Hippocampal Damage. Neurochem. Res. 2019, 44, 1306–1322. [Google Scholar] [CrossRef]
- Mu, J.; Xie, P.; Yang, Z.-S.; Yang, D.-L.; Lv, F.-J.; Luo, T.-Y.; Li, Y. Neurogenesis and major depression: Implications from proteomic analyses of hippocampal proteins in a rat depression model. Neurosci. Lett. 2007, 416, 252–256. [Google Scholar] [CrossRef]
- Vythilingam, M.; Heim, C.; Newport, J.; Miller, A.H.; Anderson, E.; Bronen, R.; Brummer, M.; Staib, L.; Vermetten, E.; Charney, D.S.; et al. Childhood Trauma Associated With Smaller Hippocampal Volume in Women With Major Depression. Am. J. Psychiatry 2002, 159, 2072–2080. [Google Scholar] [CrossRef]
- Liu, Z.; Han, Y.S.; Zhao, H.Q.; Luo, W.X.; Jia, L.; Wang, Y.H. Glu-mGluR2/3-ERK Signaling Regulates Apoptosis of Hippocampal Neurons in Diabetic-Depression Model Rats. Evid. Based Complement Altern. Med. 2019, 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.W.; Seo, J.-H.; Baek, S.-B.; Kim, C.-J.; Kim, T.-W. Treadmill exercise inhibits hippocampal apoptosis through enhancing N-methyl-D-aspartate receptor expression in the MK-801-induced schizophrenic mice. J. Exerc. Rehabil. 2014, 10, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, M.O.; Mantese, C.E.; Porciúncula, L.O.; Ghisleni, G.; Vinade, L.; Souza, D.O.; Portela, L.V. Exercise affects glutamate receptors in postsynaptic densities from cortical mice brain. Brain Res. 2005, 1065, 20–25. [Google Scholar] [CrossRef]
- VanLeeuwen, J.-E.; Petzinger, G.M.; Walsh, J.P.; Akopian, G.K.; Vuckovic, M.; Jakowec, M.W. Altered AMPA receptor expression with treadmill exercise in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse model of basal ganglia injury. J. Neurosci. Res. 2009, 88, 650–668. [Google Scholar] [CrossRef] [PubMed]
- Beneyto, M.; Kristiansen, L.V.; Oni-Orisan, A.; McCullumsmith, R.E.; Meador-Woodruff, J.H. Abnormal Glutamate Receptor Expression in the Medial Temporal Lobe in Schizophrenia and Mood Disorders. Neuropsychopharmacology 2007, 32, 1888–1902. [Google Scholar] [CrossRef]
- Pawlak, R.; Rao, B.S.S.; Melchor, J.P.; Chattarji, S.; McEwen, B.S.; Strickland, S. Tissue plasminogen activator and plasminogen mediate stress-induced decline of neuronal and cognitive functions in the mouse hippocampus. Proc. Natl. Acad. Sci. USA 2005, 102, 18201–18206. [Google Scholar] [CrossRef] [Green Version]
- Park, M.-J.; Seo, B.A.; Lee, B.; Shin, H.-S.; Kang, M.-G. Stress-induced changes in social dominance are scaled by AMPA-type glutamate receptor phosphorylation in the medial prefrontal cortex. Sci. Rep. 2018, 8, 15008. [Google Scholar] [CrossRef]
- Benz, B.; Grima, G.; Do, K.Q. Glutamate-induced homocysteic acid release from astrocytes: Possible implication in glia-neuron signaling. Neuroscience 2004, 124, 377–386. [Google Scholar] [CrossRef]
- Rogers, M.A.; Yamasue, H.; Kasai, K.; Information, R. Antidepressant Medication May Moderate the Effect of Depression Duration on Hippocampus Volume. Psychophysiology 2016, 30, 1–8. [Google Scholar] [CrossRef]
- Bleich, S.; Degner, D.; Sperling, W.; Bönsch, D.; Thürauf, N.; Kornhuber, J. Homocysteine as a neurotoxin in chronic alcoholism. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2004, 28, 453–464. [Google Scholar] [CrossRef]
- Abushik, P.A.; Karelina, T.V.; Sibarov, D.A.; Stepanenko, Y.D.; Giniatullin, R.A.; Antonov, S.M. Homocysteine-induced membrane currents, calcium responses and changes in mitochondrial potential in rat cortical neurons. J. Evol. Biochem. Physiol. 2015, 51, 296–304. [Google Scholar] [CrossRef]
- Almeida, O.P.; Flicker, L.; Lautenschlager, N.T.; Leedman, P.; Vasikaran, S.; Van Bockxmeer, F.M. Contribution of the MTHFR gene to the causal pathway for depression, anxiety and cognitive impairment in later life. Neurobiol. Aging 2005, 26, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, P.; Singh, N. Homocysteine excess: Delineating the possible mechanism of neurotoxicity and depression. Fundam. Clin. Pharmacol. 2015, 29, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Kuhn, H.G.; Gage, F.H.F. More hippocampal neurons in adult mice living in an enriched environment. Nat. Cell Biol. 1997, 386, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Liu, Y.; Li, W.; Yang, B.; Chen, D.; Wang, X.; Jiang, Z.; Wang, H.; Wang, Z.; Cornelisson, G.; et al. Beneficial effects of exercise and its molecular mechanisms on depression in rats. Behav. Brain Res. 2006, 168, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Han, L.; Ambrogini, E.; Weinstein, R.S.; Manolagas, S.C. Glucocorticoids and Tumor Necrosis Factor α Increase Oxidative Stress and Suppress Wnt Protein Signaling in Osteoblasts. J. Biol. Chem. 2011, 286, 44326–44335. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.-L.; Tang, X. Effect of glucocorticoid-induced oxidative stress on the expression of Cbfa1. Chem. Interact. 2014, 207, 26–31. [Google Scholar] [CrossRef]
- Numakawa, T.; Adachi, N.; Richards, M.; Chiba, S.; Kunugi, H. Brain-derived neurotrophic factor and glucocorticoids: Reciprocal influence on the central nervous system. Neuroscience 2013, 239, 157–172. [Google Scholar] [CrossRef]
- Dunn, A.L.; Reigle, T.G.; Youngstedt, S.D.; Armstrong, R.B.; Dishman, R.K. Brain norepinephrine and metabolites after treadmill training and wheel running in rats. Med. Sci. Sports Exerc. 1996, 28, 204–209. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.; Wang, D.; Duan, Y.; Zhai, L.; Shi, L.; Guo, F. Aerobic Exercise Prevents Depression via Alleviating Hippocampus Injury in Chronic Stressed Depression Rats. Brain Sci. 2021, 11, 9. https://doi.org/10.3390/brainsci11010009
Kang J, Wang D, Duan Y, Zhai L, Shi L, Guo F. Aerobic Exercise Prevents Depression via Alleviating Hippocampus Injury in Chronic Stressed Depression Rats. Brain Sciences. 2021; 11(1):9. https://doi.org/10.3390/brainsci11010009
Chicago/Turabian StyleKang, Jie, Di Wang, Yongchang Duan, Lin Zhai, Lin Shi, and Fei Guo. 2021. "Aerobic Exercise Prevents Depression via Alleviating Hippocampus Injury in Chronic Stressed Depression Rats" Brain Sciences 11, no. 1: 9. https://doi.org/10.3390/brainsci11010009
APA StyleKang, J., Wang, D., Duan, Y., Zhai, L., Shi, L., & Guo, F. (2021). Aerobic Exercise Prevents Depression via Alleviating Hippocampus Injury in Chronic Stressed Depression Rats. Brain Sciences, 11(1), 9. https://doi.org/10.3390/brainsci11010009