
Effect of Sensory Deprivation of Nasal Respiratory on Behavior of C57BL/6J Mice


Abstract
:1. Introduction
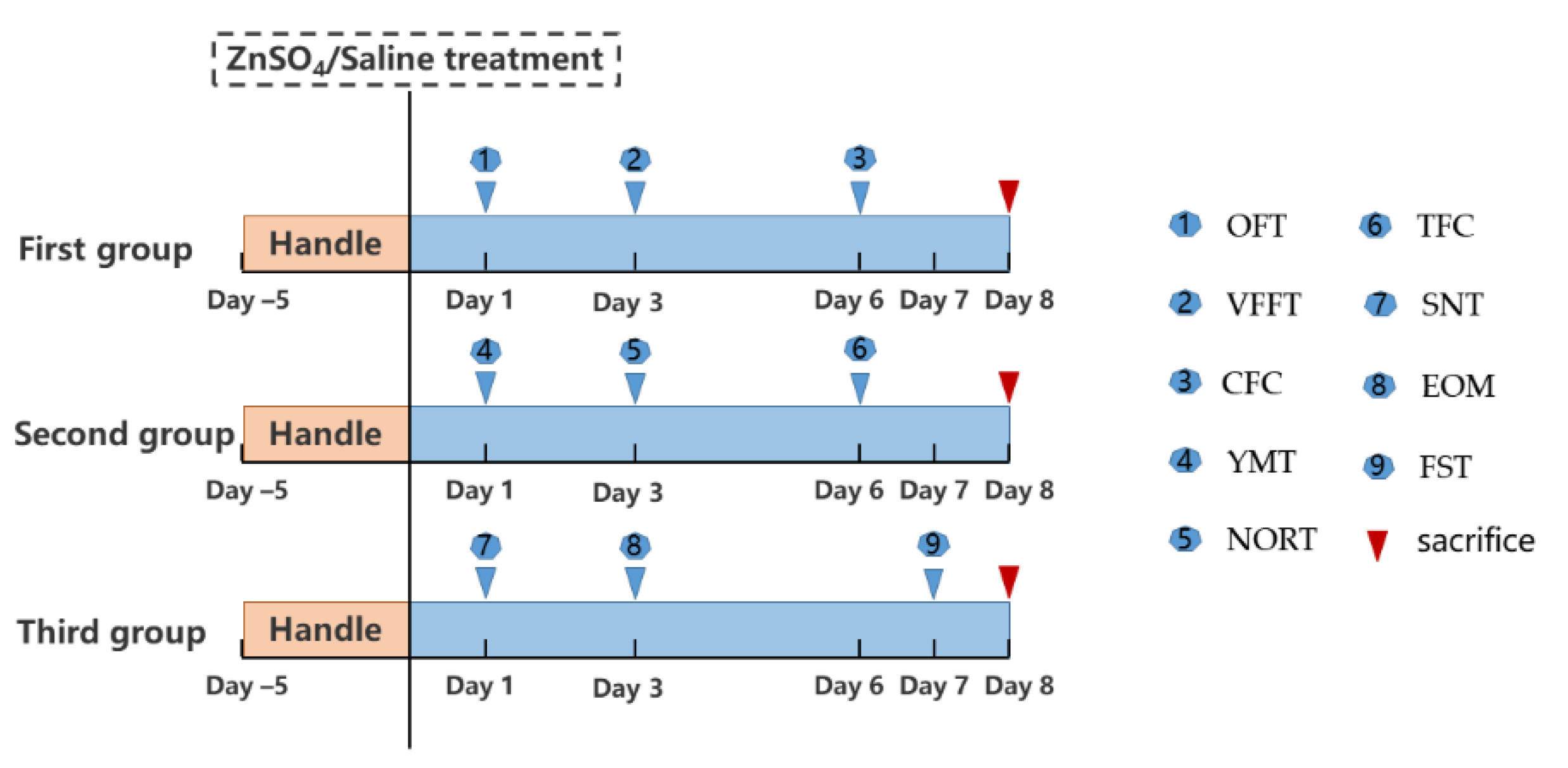
2. Materials and Methods
2.1. Animals
2.2. Nasal Irrigation of Zinc Sulfate
2.3. Behavioral Test Battery
2.3.1. Open Field Test (OFT)
2.3.2. Elevated O Maze (EOM)
2.3.3. Forced Swim Test (FST)
2.3.4. Y-Maze Test (YMT)
2.3.5. Novel Object Recognition Test (NORT)
2.3.6. Contextual Fear Conditioning Test (CFC)
2.3.7. Tone-Cued Fear Conditioning Test (TFC)
2.3.8. Social Novelty Test (SNT)
2.3.9. Von Frey Filaments Test (VFFT)
2.3.10. Food-Seeking Test
2.3.11. Statistical Analysis
3. Results
3.1. Open Field Test
3.2. Elevated O Maze
3.3. Forced Swim Test
3.4. Y-Maze Test
3.5. Novel Object Recognition Test
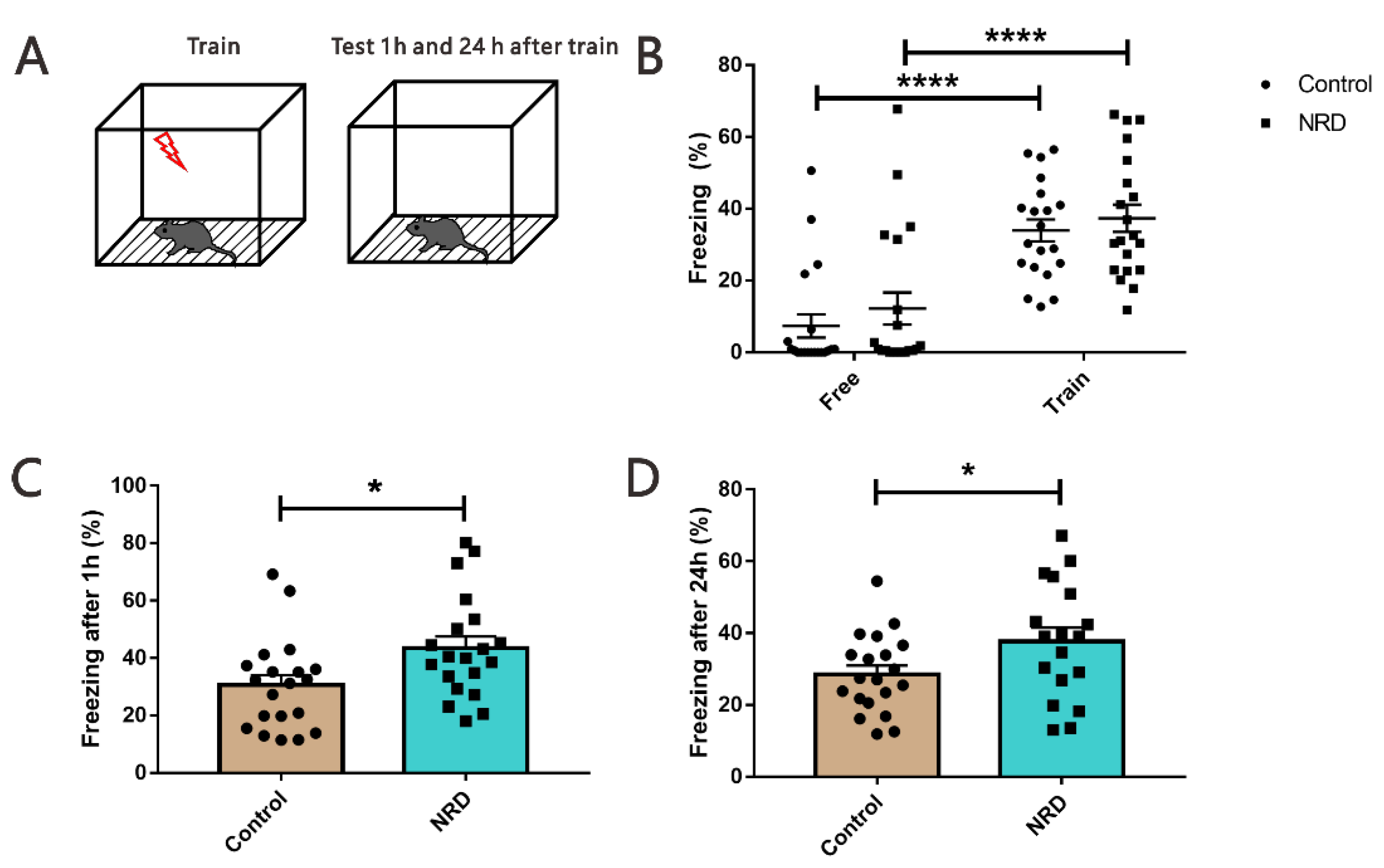
3.6. Fear Conditioning Test
3.7. Social Novelty Test
3.8. Von Frey Filaments Test
4. Discussion
5. Conclusions
Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldman, J.L.; Del Negro, C.A. Looking for inspiration: New perspectives on respiratory rhythm. Nat. Rev. Neurosci. 2006, 7, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Moberly, A.H.; Schreck, M.; Bhattarai, J.P.; Zweifel, L.S.; Luo, W.; Ma, M. Olfactory inputs modulate respiration-related rhythmic activity in the prefrontal cortex and freezing behavior. Nat. Commun. 2018, 9, 1528. [Google Scholar] [CrossRef]
- Zelano, C.; Jiang, H.; Zhou, G.; Arora, N.; Schuele, S.; Rosenow, J.; Gottfried, J.A. Nasal Respiration Entrains Human Limbic Oscillations and Modulates Cognitive Function. J. Neurosci. 2016, 36, 12448–12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, A.V.; Makuch, M.Y.; Setubal, M.S.; Barros, N.F.; Bahamondes, L. A Qualitative Study on the Practice of Yoga for Women with Pain-Associated Endometriosis. J. Altern. Complement. Med. 2016, 22, 977–982. [Google Scholar] [CrossRef]
- Grosmaitre, X.; Santarelli, L.C.; Tan, J.; Luo, M.; Ma, M. Dual functions of mammalian olfactory sensory neurons as odor detectors and mechanical sensors. Nat. Neurosci. 2007, 10, 348–354. [Google Scholar] [CrossRef]
- Yanovsky, Y.; Ciatipis, M.; Draguhn, A.; Tort, A.B.; Brankack, J. Slow oscillations in the mouse hippocampus entrained by nasal respiration. J. Neurosci. 2014, 34, 5949–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W.; Ciatipis, M.; Wolfenstetter, T.; Jessberger, J.; Muller, C.; Ponsel, S.; Yanovsky, Y.; Brankack, J.; Tort, A.B.L.; Draguhn, A. Selective entrainment of gamma subbands by different slow network oscillations. Proc. Natl. Acad. Sci. USA. 2017, 114, 4519–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biskamp, J.; Bartos, M.; Sauer, J.F. Organization of prefrontal network activity by respiration-related oscillations. Sci. Rep. 2017, 7, 45508. [Google Scholar] [CrossRef] [Green Version]
- Ito, J.; Roy, S.; Liu, Y.; Cao, Y.; Fletcher, M.; Lu, L.; Boughter, J.D.; Grun, S.; Heck, D.H. Whisker barrel cortex delta oscillations and gamma power in the awake mouse are linked to respiration. Nat. Commun. 2014, 5, 3572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tort, A.B.L.; Brankack, J.; Draguhn, A. Respiration-Entrained Brain Rhythms Are Global but Often Overlooked. Trends Neurosci. 2018, 41, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Zhang, B.; Qin, Z.; Wang, P. Nasal Respiration is Necessary for the Generation of gamma Oscillation in the Olfactory Bulb. Neuroscience 2018, 398, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; McAfee, S.S.; Heck, D.H. Hippocampal sharp-wave ripples in awake mice are entrained by respiration. Sci. Rep. 2017, 7, 8950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.K.; Gilbey, T.; Barnett, S.C. Immunohistochemical studies of the cellular changes in the peripheral olfactory system after zinc sulfate nasal irrigation. Neurochem. Res. 2004, 29, 891–901. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Morgan, J.A.; Singhal, G.; Corrigan, F.; Jaehne, E.J.; Jawahar, M.C.; Baune, B.T. The effects of aerobic exercise on depression-like, anxiety-like, and cognition-like behaviours over the healthy adult lifespan of C57BL/6 mice. Behav. Brain Res. 2018, 337, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Can, A.; Dao, D.T.; Arad, M.; Terrillion, C.E.; Piantadosi, S.C.; Gould, T.D. The mouse forced swim test. J. Vis. Exp. 2012, 59, e3638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.T.; Olton, D.S.; Gage, F.H., 3rd; Jenko, P.G. Damage to hippocampus and hippocampal connections: Effects on DRL and spontaneous alternation. J. Comp. Physiol. Psychol. 1977, 91, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.J.; Stackman, R.W., Jr. Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav Brain Res. 2015, 285, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Saxe, M.D.; Battaglia, F.; Wang, J.W.; Malleret, G.; David, D.J.; Monckton, J.E.; Garcia, A.D.; Sofroniew, M.V.; Kandel, E.R.; Santarelli, L.; et al. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proc. Natl. Acad. Sci. USA. 2006, 103, 17501–17506. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Li, J.; Zhou, G.; Yang, J.; McKenzie, S.; Li, Y.; Li, W.; Yu, J.; Wang, Y.; Qu, J.; et al. Gating of hippocampal rhythms and memory by synaptic plasticity in inhibitory interneurons. Neuron 2021, 109, 1013–1028.e9. [Google Scholar] [CrossRef] [PubMed]
- Faizi, M.; Bader, P.L.; Saw, N.; Nguyen, T.V.; Beraki, S.; Wyss-Coray, T.; Longo, F.M.; Shamloo, M. Thy1-hAPP(Lond/Swe+) mouse model of Alzheimer’s disease displays broad behavioral deficits in sensorimotor, cognitive and social function. Brain Behav. 2012, 2, 142–154. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Li, S.; Li, W.; Wu, X.; Li, J.; Yang, J.; Tu, C.; Ye, X.; Ling, S. Olfactory deficit is associated with mitral cell dysfunction in the olfactory bulb of P301S tau transgenic mice. Brain Res. Bull. 2019, 148, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, L.; Alessandri, M.G.; Tola, J.; Putignano, E.; Migliore, M.; Amendola, E.; Gross, C.; Leuzzi, V.; Cioni, G.; Pizzorusso, T. A novel mouse model of creatine transporter deficiency. F1000Res. 2014, 3, 228. [Google Scholar] [CrossRef] [Green Version]
- Kalawski, J.P. The Alba Method and the Science of Emotions. Integr. Psychol. Behav. Sci. 2020, 54, 903–919. [Google Scholar] [CrossRef]
- Homma, I.; Masaoka, Y. Breathing rhythms and emotions. Exp. Physiol. 2008, 93, 1011–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaccaro, A.; Piarulli, A.; Laurino, M.; Garbella, E.; Menicucci, D.; Neri, B.; Gemignani, A. How Breath-Control Can Change Your Life: A Systematic Review on Psycho-Physiological Correlates of Slow Breathing. Front. Hum. Neurosci. 2018, 12, 353. [Google Scholar] [CrossRef] [Green Version]
- Hare, B.D.; Duman, R.S. Prefrontal cortex circuits in depression and anxiety: Contribution of discrete neuronal populations and target regions. Mol. Psychiatry 2020, 25, 2742–2758. [Google Scholar] [CrossRef] [PubMed]
- Arshamian, A.; Iravani, B.; Majid, A.; Lundstrom, J.N. Respiration Modulates Olfactory Memory Consolidation in Humans. J. Neurosci. 2018, 38, 10286–10294. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.G.; LeDoux, J.E. Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav. Neurosci. 1992, 106, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Bagur, S.; Lefort, J.M.; Lacroix, M.M.; de Lavilleon, G.; Herry, C.; Chouvaeff, M.; Billand, C.; Geoffroy, H.; Benchenane, K. Breathing-driven prefrontal oscillations regulate maintenance of conditioned-fear evoked freezing independently of initiation. Nat. Commun. 2021, 12, 2605. [Google Scholar] [CrossRef]
- Marek, R.; Strobel, C.; Bredy, T.W.; Sah, P. The amygdala and medial prefrontal cortex: Partners in the fear circuit. J. Physiol. 2013, 591, 2381–2391. [Google Scholar] [CrossRef] [PubMed]
- Goosens, K.A.; Maren, S. NMDA receptors are essential for the acquisition, but not expression, of conditional fear and associative spike firing in the lateral amygdala. Eur. J. Neurosci. 2004, 20, 537–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corcoran, K.A.; Quirk, G.J. Activity in prelimbic cortex is necessary for the expression of learned, but not innate, fears. J. Neurosci. 2007, 27, 840–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anglada-Figueroa, D.; Quirk, G.J. Lesions of the basal amygdala block expression of conditioned fear but not extinction. J. Neurosci. 2005, 25, 9680–9685. [Google Scholar] [CrossRef] [Green Version]
- Knight, M.J.; Baune, B.T. Cognitive dysfunction in major depressive disorder. Curr. Opin. Psychiatry 2018, 31, 26–31. [Google Scholar] [CrossRef]
- Kupferberg, A.; Bicks, L.; Hasler, G. Social functioning in major depressive disorder. Neurosci. Biobehav. Rev. 2016, 69, 313–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacker, D.W.; Ludwig, M. Vasopressin, oxytocin, and social odor recognition. Horm. Behav. 2012, 61, 259–265. [Google Scholar] [CrossRef]
- Ossipov, M.H.; Dussor, G.O.; Porreca, F. Central modulation of pain. J. Clin. Invest. 2010, 120, 3779–3787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, W.Y.; Stohler, C.S.; Herr, D.R. Role of the Prefrontal Cortex in Pain Processing. Mol. Neurobiol. 2019, 56, 1137–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fields, H.L. Pain and the primary somatosensory cortex. Pain 2012, 153, 742–743. [Google Scholar] [CrossRef] [PubMed]
- Luers, J.C.; Rokohl, A.C.; Loreck, N.; Wawer Matos, P.A.; Augustin, M.; Dewald, F.; Klein, F.; Lehmann, C.; Heindl, L.M. Olfactory and Gustatory Dysfunction in Coronavirus Disease 2019 (COVID-19). Clin. Infect. Dis. 2020, 71, 2262–2264. [Google Scholar] [CrossRef] [PubMed]
- Dan, X.; Wechter, N.; Gray, S.; Mohanty, J.G.; Croteau, D.L.; Bohr, V.A. Olfactory dysfunction in aging and neurodegenerative diseases. Ageing Res. Rev. 2021, 70, 101416. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Choi, M.; Kim, H.; Yang, E.J.; Mahmood, U.; Kang, S.I.; Shin, H.W.; Kim, D.W.; Kim, H.S. Transient Anosmia Induces Depressive-like and Anxiolytic-like Behavior and Reduces Amygdalar Corticotropin-Releasing Hormone in a ZnSO4-Induced Mouse Model. Chem. Senses 2018, 43, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Kass, M.D.; Czarnecki, L.A.; Moberly, A.H.; McGann, J.P. Differences in peripheral sensory input to the olfactory bulb between male and female mice. Sci. Rep. 2017, 7, 45851. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | NRD | p-Value | df | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SEM | n | Mean ± SEM | n | ||||
| OFT | Distance in open field | 3499 ± 149.5 | 14 | 3171 ± 224 | 14 | 0.2336 | 26 |
| Moving speed in open field | 465 ± 28.35 | 14 | 412.5 ± 46.51 | 14 | 0.3437 | 26 | |
| Distance in center | 514.9 ± 33.11 | 14 | 399.6 ± 41.42 | 14 | 0.0390 | 26 | |
| Number of center squares entered | 42.86 ± 1.83 | 14 | 33 ± 3.063 | 14 | 0.0104 | 26 | |
| EOM | Distance in open arms | 443.6 ± 55.05 | 13 | 295.7 ± 38.91 | 13 | 0.0381 | 24 |
| Time in open arms | 170.6 ± 17.67 | 13 | 121.8 ± 14.18 | 13 | 0.0414 | 24 | |
| Number of open arms entered | 91.38 ± 7.238 | 13 | 73.23 ± 4.512 | 13 | 0.0438 | 24 | |
| FST | Struggle times | 69.96 ± 3.945 | 17 | 51.28 ± 3.468 | 17 | 0.0012 | 32 |
| YMT | Number of arms | 38.6 ± 4.498 | 15 | 41.14 ± 4.555 | 14 | 0.6946 | 27 |
| Spontaneous alternation | 66.88 ± 3.128 | 15 | 54.88 ± 4.157 | 14 | 0.0276 | 27 | |
| NORT | ORT index (%) | 24.56 ± 10.08 | 13 | −18.98 ± 10.95 | 13 | 0.0074 | 24 |
| FCT (freezing %) | Free | 7.347 ± 3.237 | 20 | 12.22 ± 4.46 | 20 | 0.3826 | 38 |
| Train | 33.96 ± 3.05 | 20 | 37.38 ± 3.791 | 20 | 0.4864 | 38 | |
| After 1 h | 30.5 ± 3.567 | 20 | 43.55 ± 4.01 | 20 | 0.0199 | 38 | |
| After 24 h | 28.52 ± 2.431 | 20 | 37.78 ± 3.809 | 18 | 0.0436 | 36 | |
| TCF (freezing %) | Free | 4.883 ± 2.757 | 16 | 4.258 ± 1.788 | 16 | 0.8504 | 30 |
| Train | 32.93 ± 4.959 | 16 | 34.26 ± 5.783 | 16 | 0.8619 | 30 | |
| After 1 h | 61.14 ± 2.89 | 15 | 48.71 ± 4.538 | 15 | 0.0285 | 28 | |
| After 24 h | 69.89 ± 4.398 | 15 | 54.38 ± 4.723 | 16 | 0.0233 | 29 | |
| SNT | Sociability; empty | 49.52 ± 4.164 | 13 | 66.78 ± 7.364 | 13 | 0.0525 | 24 |
| Sociability; stanger1 | 130.6 ± 7.801 | 13 | 111.3 ± 8.946 | 13 | 0.1157 | 24 | |
| Sociability; assessment | 2.64 ± 0.2201 | 13 | 1.762 ± 0.1841 | 13 | 0.0054 | 24 | |
| Social novelty; stranger1 | 71.12 ± 10.32 | 13 | 62.69 ± 6.437 | 13 | 0.4951 | 24 | |
| Social novelty; stranger2 | 103.7 ± 8.037 | 13 | 93.97 ± 10.85 | 13 | 0.4763 | 24 | |
| Social novelty; assessment | 2.046 ± 0.1855 | 13 | 1.469 ± 0.1854 | 12 | 0.0386 | 23 | |
| VFFT | 50% mechanical threshold | 0.2593 ± 0.02058 | 14 | 0.5686 ± 0.04063 | 14 | <0.0001 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Ye, Y.; Zhou, C.; Sun, S.; Zhang, J.; Zhao, Z.; Sun, T.; Li, J.; Yang, J.; Li, W.; et al. Effect of Sensory Deprivation of Nasal Respiratory on Behavior of C57BL/6J Mice. Brain Sci. 2021, 11, 1626. https://doi.org/10.3390/brainsci11121626
Zhu Y, Ye Y, Zhou C, Sun S, Zhang J, Zhao Z, Sun T, Li J, Yang J, Li W, et al. Effect of Sensory Deprivation of Nasal Respiratory on Behavior of C57BL/6J Mice. Brain Sciences. 2021; 11(12):1626. https://doi.org/10.3390/brainsci11121626
Chicago/Turabian StyleZhu, Yongji, Yujing Ye, Chenyang Zhou, Siqi Sun, Jingjing Zhang, Zixuan Zhao, Tingting Sun, Jing Li, Jing Yang, Weiyun Li, and et al. 2021. "Effect of Sensory Deprivation of Nasal Respiratory on Behavior of C57BL/6J Mice" Brain Sciences 11, no. 12: 1626. https://doi.org/10.3390/brainsci11121626
APA StyleZhu, Y., Ye, Y., Zhou, C., Sun, S., Zhang, J., Zhao, Z., Sun, T., Li, J., Yang, J., Li, W., & Li, S. (2021). Effect of Sensory Deprivation of Nasal Respiratory on Behavior of C57BL/6J Mice. Brain Sciences, 11(12), 1626. https://doi.org/10.3390/brainsci11121626