
Sex Differences in Spatial Memory: Comparison of Three Tasks Using the Same Virtual Context


 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Apparatus
2.3. Procedure
2.3.1. The Walking Space Boxes Room Task (WSBRT)
2.3.2. The Non-Walking Space Boxes Room Task (NWSBRT)
2.3.3. The Almeria Spatial Memory Recognition Test (ASMRT)
2.4. Statistical Analyses
3. Results
3.1. WSBRT
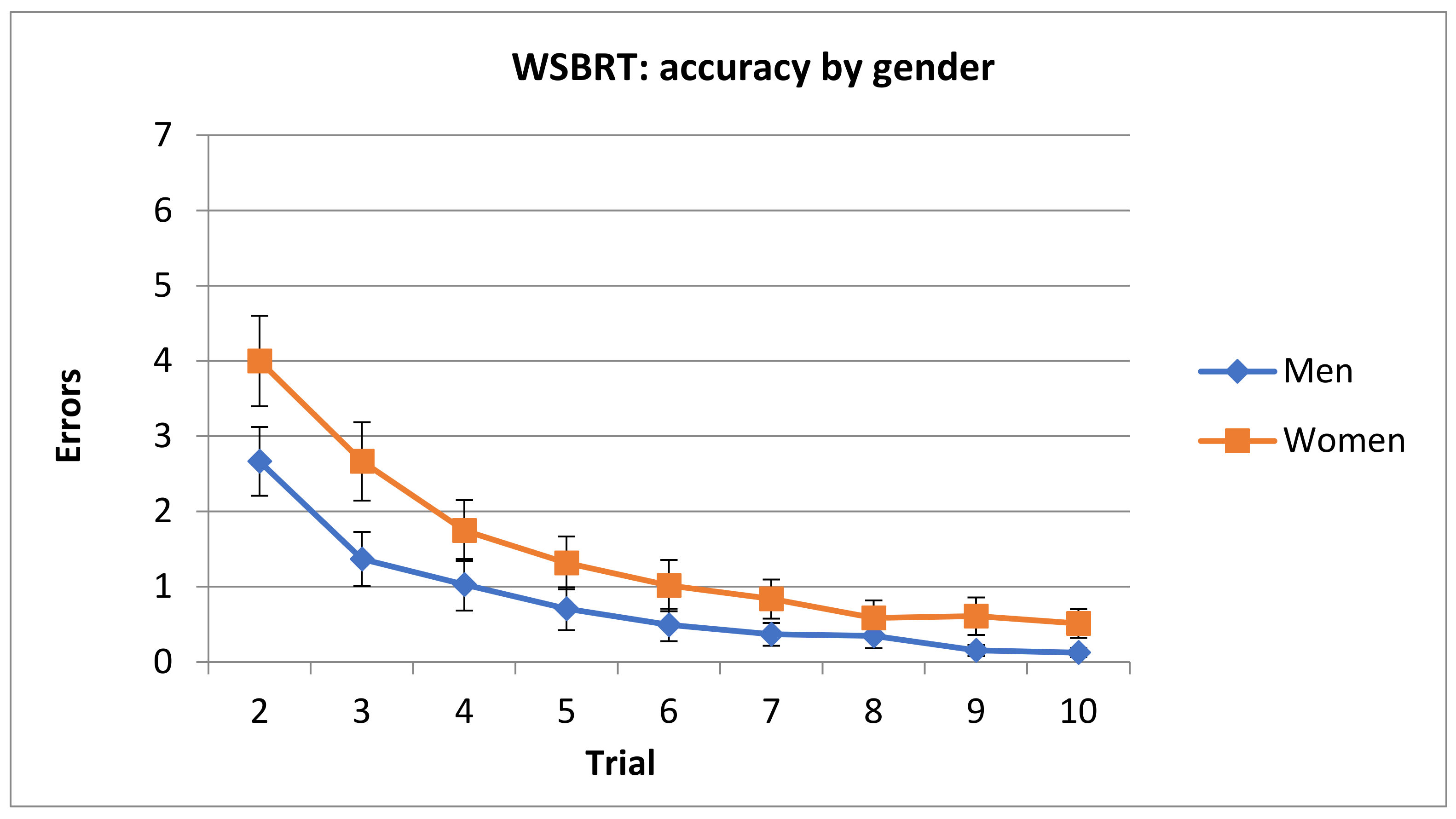
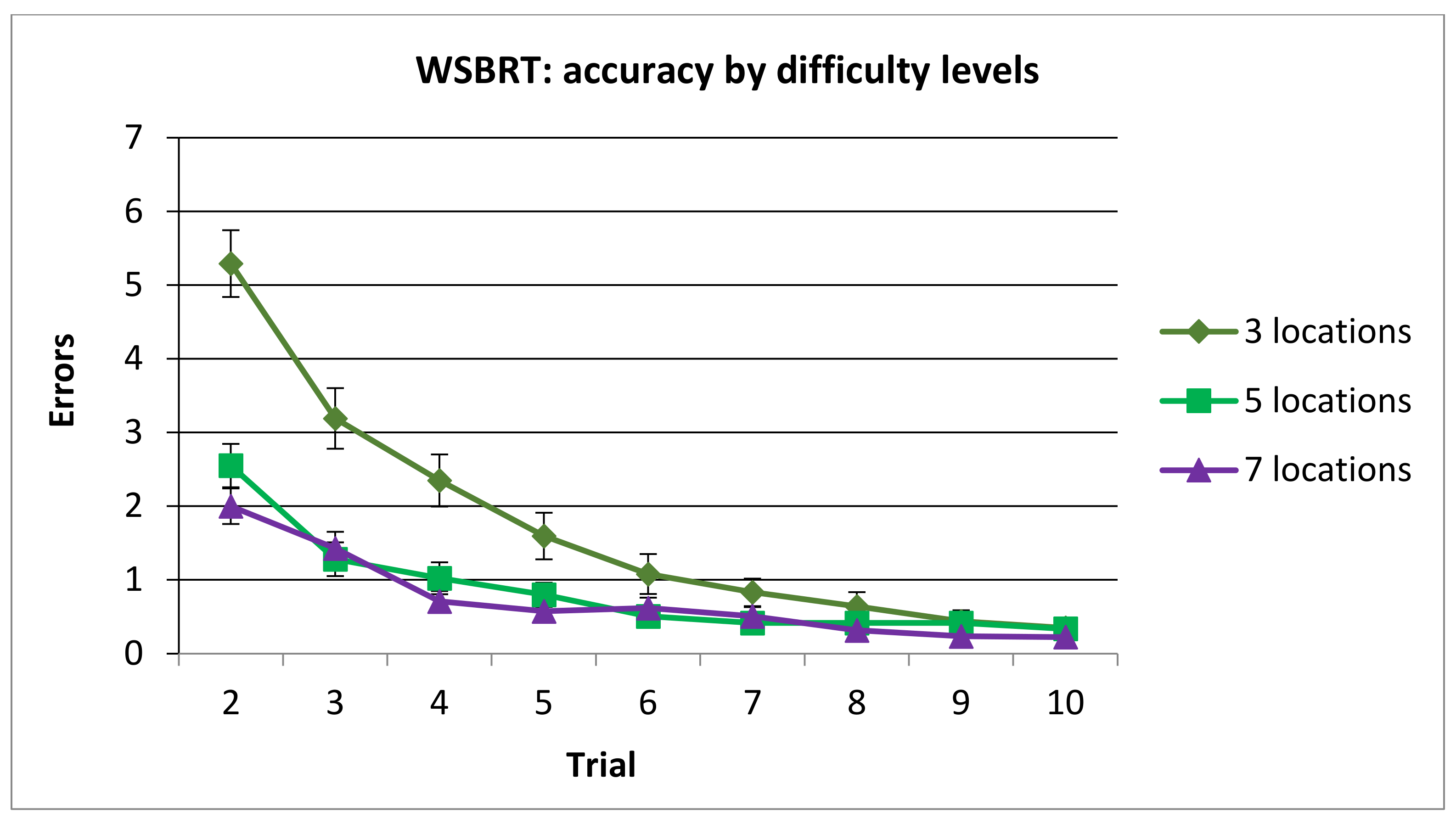
3.1.1. Number of Errors
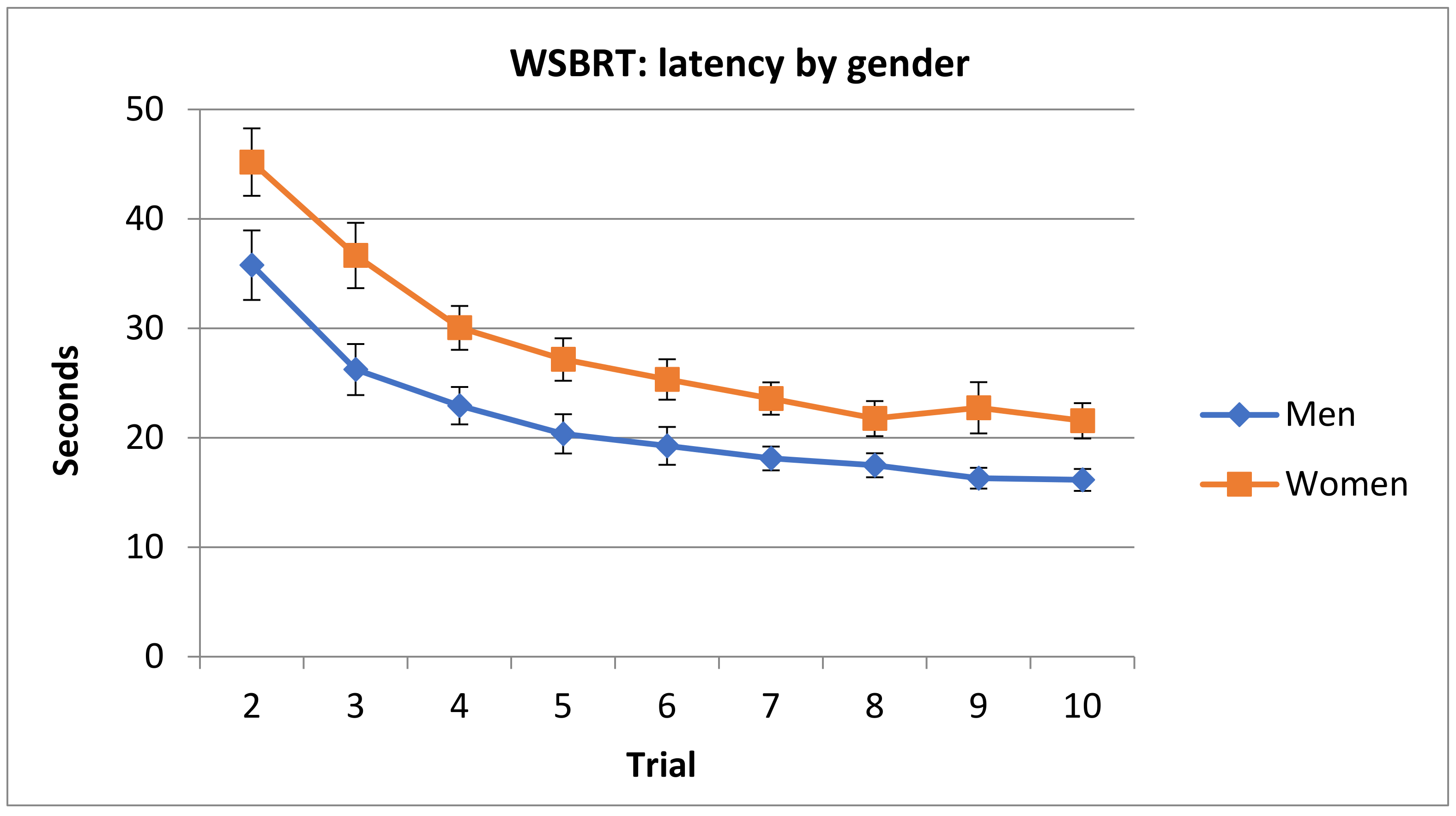
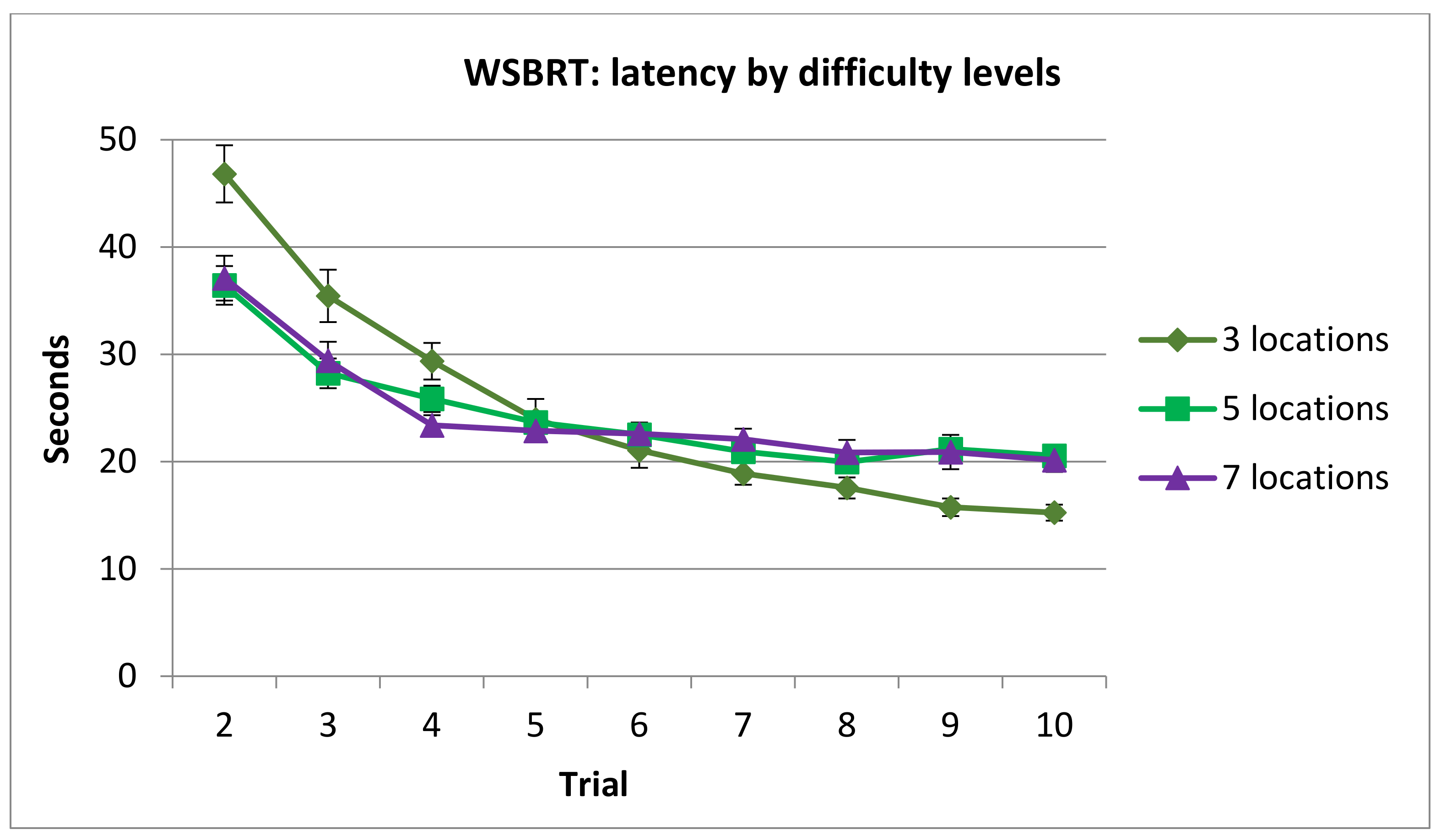
3.1.2. Latency
3.2. NWSBRT
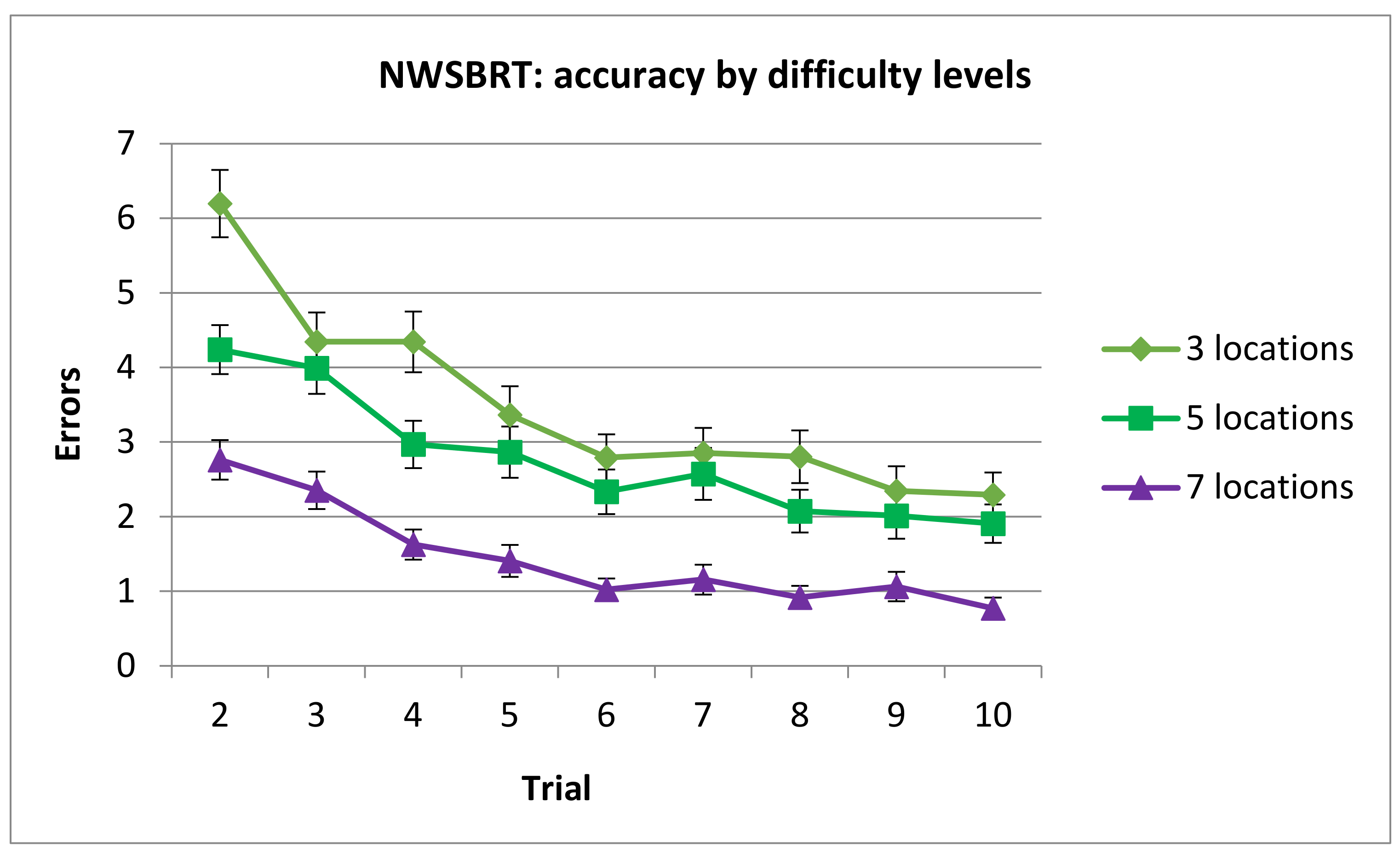
3.2.1. Number of Errors
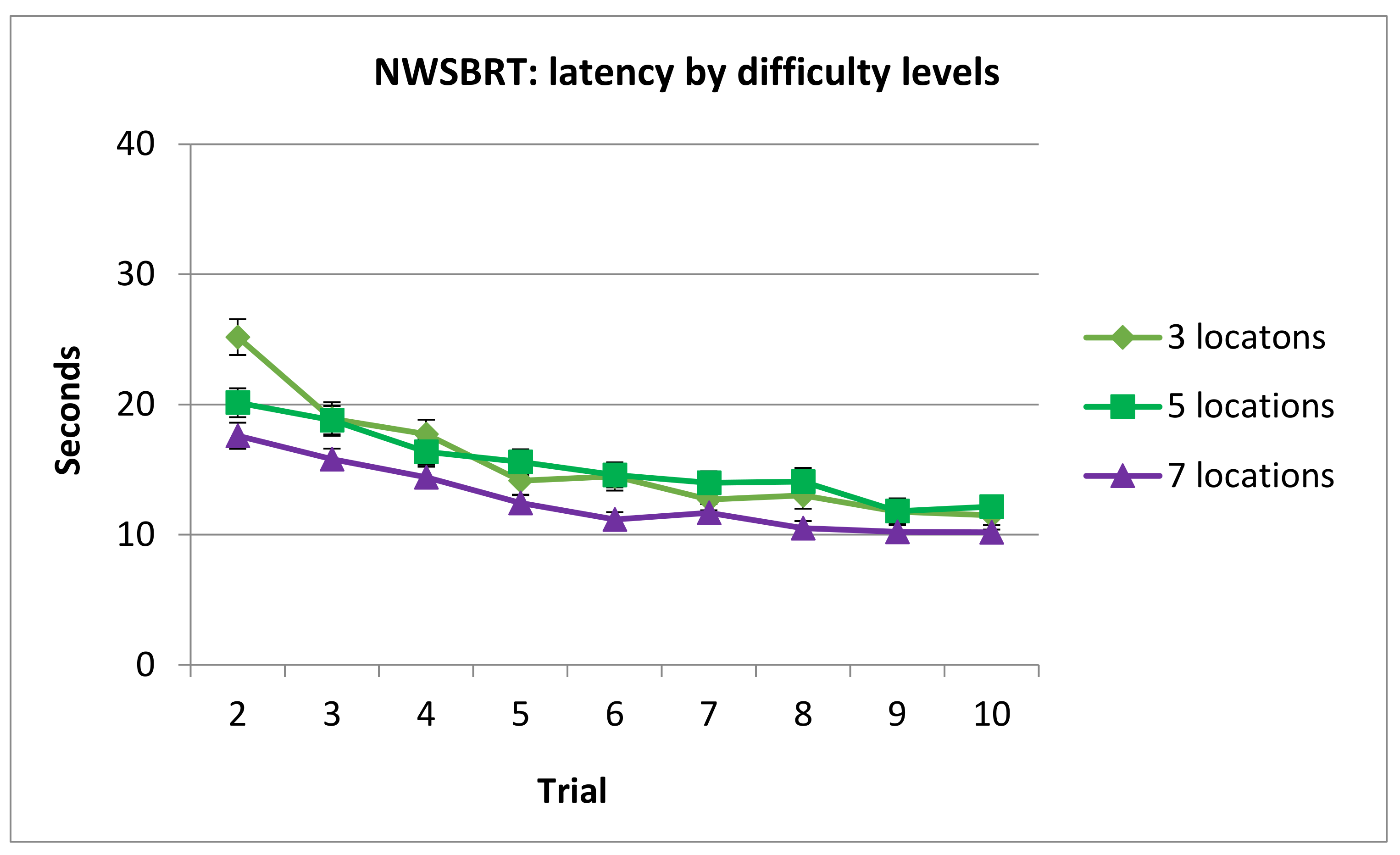
3.2.2. Latency
3.3. ASMRT
3.3.1. Number of Correct Answers
3.3.2. Reaction Time
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vorhees, C.V.; Williams, M.T. Assessing spatial learning and memory in rodents. ILAR J. 2014, 55, 310–332. [Google Scholar] [CrossRef] [PubMed]
- Montello, D.R. Geographic orientation and disorientation: Getting lost and getting found in real and information spaces. User Exp. Mag. 2009, 8, 24–26. [Google Scholar] [CrossRef]
- Nemmi, F.; Boccia, M.; Piccardi, L.; Galati, G.; Guariglia, C. Segregation of neural circuits involved in spatial learning in reaching and navigational space. Neuropsychologia 2013, 51, 1561–1570. [Google Scholar] [CrossRef]
- Piccardi, L.; Berthoz, A.; Baulac, M.; Denos, M.; Dupont, S.; Samson, S.; Guariglia, C. Different spatial memory systems are involved in small-and large-scale environments: Evidence from patients with temporal lobe epilepsy. Exp. Brain Res. 2010, 206, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Edler, D.; Bestgen, A.-K.; Kuchinke, L.; Dickmann, F. Grids in topographic maps reduce distorsions in the recall or learned object locations. PLoS ONE 2014, 28. [Google Scholar] [CrossRef]
- Gronau, N.; Neta, M.; Bar, M. Integrated Contextual representation for objects’ identities and their locations. J. Cogn. Neurosci. 2008, 20, 371–388. [Google Scholar] [CrossRef] [PubMed]
- Wolbers, T.; Hegarty, M. What determines our navigational abilities? Trends Cog. Sci. 2010, 14, 138–146. [Google Scholar] [CrossRef]
- Gagnon, K.T.; Thomas, B.J.; Munion, A.; Creem-Regehr, S.H.; Cashdan, E.A.; Stefanucci, J.K. Not all those who wander are lost: Spatial exploration patterns and their relationship to gender and spatial memory. Cognition 2018, 180, 108–117. [Google Scholar] [CrossRef]
- Guderian, S.; Dzieciol, A.M.; Gadian, D.G.; Jentschke, S.; Doeller, C.F.; Burgess, N.; Mishkin, M.; Vargha-Khadem, F. Hippocampal volume reduction in humans predicts impaired allocentric spatial memory in virtual-reality navigation. J. Neurosci. 2015, 35, 14123–14131. [Google Scholar] [CrossRef]
- Nowak, N.T.; Murali, A.; Driscoll, I. Factors related to sex differences in navigating a computerized maze. J. Environ. Psychol. 2015, 43, 136–144. [Google Scholar] [CrossRef]
- Lee, C.H.; Ryu, J.; Lee, S.H.; Kim, H.; Lee, I. Functional cross-hemispheric shift between object-place paired associate memory and spatial memory in the human hippocampus. Hippocampus 2016, 26, 1061–1077. [Google Scholar] [CrossRef]
- Lancia, S.; Cofini, V.; Carrieri, M.; Ferrari, M.; Quaresima, V. Are ventrolateral and dorsolateral prefrontal cortices involved in the computerized Corsi block-tapping test execution? An fNIRS study. Neurophotonics 2018, 5, 011019. [Google Scholar] [CrossRef] [PubMed]
- Snopková, D.; Ugwuitz, P.; Stachon, Z.; Hladík, J.; Jurik, V.; Kvarda, O.; Kubícek, P. Retracing evacuation strategy: A virtual reality game-based investigation into the influence of building’s spatial configuration in an emergency. Spa. Cogn. Comp. 2021. [Google Scholar] [CrossRef]
- Ugwitz, P.; Jurik, V.; Herman, L.; Stachon, Z.; Kubicek, P.; Sasinka, C. Spatial analysis of navigation in virtual geographic environments. Appl. Sci. 2019, 9, 1873. [Google Scholar] [CrossRef]
- Sogol, M.; Moussavi, Z. Does practicing with a virtual reality driving simulator improve spatial cognition in older adults? A pilot study. Neurosci Insights 2020, 15. [Google Scholar] [CrossRef]
- Montefinese, M.; Sulpizio, V.; Galati, G.; Committeri, G. Age-related effects on spatial memory across viewpoint changes relative to different reference frames. Psychol. Res. 2015, 79, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Plancher, G.; Barra, J.; Orriols, E.; Piolino, P. The influence of action on episodic memory: A virtual reality study. Q. J. Exp. Psychol. 2013, 66, 895–909. [Google Scholar] [CrossRef]
- Reggente, N.; Essoe, J.K.Y.; Aghajan, Z.M.; Tavakoli, A.V.; McGuire, J.F.; Suthana, N.A.; Rissman, J. Enhancing the ecological validity of fMRI memory research using virtual reality. Front. Neurosci. 2018, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tascón, L.; Boccia, M.; Piccardi, L.; Cimadevilla, J.M. Differences in spatial memory recognition due to cognitive style. Front. Pharmacol. 2017, 8, 1–7. [Google Scholar] [CrossRef]
- Antonova, E.; Parslow, D.; Brammer, M.; Simmons, A.; Williams, S.; Dawson, G.R.; Morris, R. Scopolamine disrupts hippocampal activity during allocentric spatial memory in humans: An fMRI study using a virtual reality analogue of the Morris Water Maze. J. Psychopharmacol. 2011, 25, 1256–1265. [Google Scholar] [CrossRef]
- Korman, M.; Weiss, P.L.; Hochhauser, M.; Kizony, R. Effect of age on spatial memory performance in real museum vs. computer simulation. BMC Geriatr. 2019, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Frick, A.; Möhring, W.; Newcombe, N.S. Picturing perspectives: Development of perspective-taking abilities in 4- to 8-year-olds. Front. Psychol. 2014, 5, 1–7. [Google Scholar] [CrossRef]
- Plancher, G.; Tirard, A.; Gyselinck, V.; Nicolas, S.; Piolino, P. Using virtual reality to characterize episodic memory profiles in amnestic mild cognitive impairment and Alzheimer’s disease: Influence of active and passive encoding. Neuropsychologia 2012, 50, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Tascón, L.; León, I.; Cimadevilla, J.M. Viewpoint-related gender differences in a spatial recognition task. Learn. Individ. Differ. 2016, 50, 270–274. [Google Scholar] [CrossRef]
- Tascón, L.; García-Moreno, L.M.; Cimadevilla, J.M. Almeria spatial memory recognition test (ASMRT): Gender differences emerged in a new passive spatial task. Neurosci. Lett. 2017, 651, 188–191. [Google Scholar] [CrossRef]
- Tascón, L.; Castillo, J.; León, I.; Cimadevilla, J.M. Walking and non-walking space in an equivalent virtual reality task: Sexual dimorphism and aging decline of spatial abilities. Behav. Brain Res. 2018, 347, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Bures, J.; Fenton, A.A. Neurophysiology of spatial cognition. News Physiol. Sci. 2000, 15, 233–240. [Google Scholar] [CrossRef]
- Wang, C.; Chen, X.; Knierim, J.J. Egocentric and allocentric representations of space in the rodent brain. Curr. Opin. Neurobiol. 2020, 60, 12–20. [Google Scholar] [CrossRef]
- Miltiadous, P.; Stamatakis, A.; Koutsoudaki, P.N.; Tiniakos, D.G.; Stylianopoulou, F. IGF-I ameliorates hippocampal neurodegeneration and protects against cognitive deficits in an animal model of temporal lobe epilepsy. Exp. Neurol. 2011, 231, 223–235. [Google Scholar] [CrossRef]
- Ekstrom, A.D.; Arnold, A.E.G.F.; Iaria, G. A critical review of the allocentric spatial representation and its neural underpinnings: Toward a network-based perspective. Front. Hum. Neurosci. 2014, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nori, R.; Piccardi, L.; Maialetti, A.; Goro, M.; Rossetti, A.; Argento, O.; Guariglia, C. No gender differences in egocentric and allocentric environmental transformation after compensating for male advantage by manipulating familiarity. Front. Neurosci. 2018, 12, 1–9. [Google Scholar] [CrossRef]
- Palmiero, M.; Nori, R.; Rogolino, C.; D’Amico, S.; Piccardi, L. Situated navigational working memory: The role of positive mood. Cogn. Process. 2015, 16, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Piccardi, L.; Risetti, M.; Nori, R.; Tanzilli, A.; Bernardi, L.; Guariglia, C. Perspective changing in primary and secondary learning: A gender difference study. Learn. Individ. Differ. 2011, 21, 114–118. [Google Scholar] [CrossRef]
- Cánovas, R.; Espínola, M.; Iribarne, L.; Cimadevilla, J.M. A new virtual task to evaluate human place learning. Behav. Brain Res. 2008, 190, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Coluccia, E.; Louse, G. Gender differences in spatial orientation: A review. J. Environ. Psychol. 2004, 24, 329–340. [Google Scholar] [CrossRef]
- Nori, R.; Piccardi, L.; Migliori, M.; Guidazzoli, A.; Frasca, F.; de Luca, D.; Giusberti, F. The virtual reality Walking Corsi Test. Comput. Human Behav. 2015, 48, 72–77. [Google Scholar] [CrossRef]
- Piccardi, L.; Bianchini, F.; Iasevoli, L.; Giannone, G.; Guariglia, C. Sex differences in a landmark environmental re-orientation task only during the learning phase. Neurosci. Lett. 2011, 503, 181–185. [Google Scholar] [CrossRef]
- Cerles, M.; Gomez, A.; Rousset, S. An implicit spatial memory alignment effect. Cogn. Process. 2015, 16, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Tlauka, M.; Carter, P.; Mahlberg, T.; Wilson, P.N. The first-perspective alignment effect: The role of environmental complexity and familiarity with surroundings. Q. J. Exp. Psychol. 2011, 64, 2236–2250. [Google Scholar] [CrossRef]
- Török, Á.; Peter Nguyen, T.; Kolozsvári, O.; Buchanan, R.J.; Nadasdy, Z. Reference frames in virtual spatial navigation are viewpoint dependent. Front. Hum. Neurosci. 2014, 8, 1–10. [Google Scholar] [CrossRef]
- Piber, D.; Nowacki, J.; Mueller, S.C.; Wingenfeld, K.; Otte, C. Sex effects on spatial learning but not on spatial memory retrieval in healthy young adults. Behav. Brain Res. 2018, 336, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Picucci, L.; Caffò, A.O.; Bosco, A. Besides navigation accuracy: Gender differences in strategy selection and level of spatial confidence. J. Environ. Psychol. 2011, 31, 430–438. [Google Scholar] [CrossRef]
- Yuan, L.; Kong, F.; Luo, Y.; Zeng, S.; Lan, J.; You, X. Gender differences in large-scale and small-scale spatial ability: A systematic review based on behavioral and neuroimaging research. Front. Behav. Neurosci. 2019, 13, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Bocchi, A.; Palmiero, M.; Piccardi, L. Travel planning in men and women. Who is better? Curr. Psychol. 2021. [Google Scholar] [CrossRef]
- Corre, C.; Friedel, M.; Vousden, D.A.; Metcalf, A.; Spring, S.; Qiu, L.R.; Lerch, J.P.; Palmert, M.R. Separate effects of sex hormones and sex chromosomes on brain structure and function revealed by high-resolution magnetic resonance imaging and spatial navigation assessment of the Four Core Genotype mouse model. Brain Struct. Funct. 2016, 221, 997–1016. [Google Scholar] [CrossRef]
- Guigueno, M.F.; MacDougallShackleton, S.A.; Sherry, D.F. Sex differences in spatial memory in brown-headed cowbirds: Males outperform females on a touchscreen task. PLoS ONE 2015, 17, 1–18. [Google Scholar] [CrossRef]
- León, I.; Cimadevilla, J.M.; Tascón, L. Developmental gender differences in children in a virtual spatial memory task. Neuropsychology 2014, 28, 485–495. [Google Scholar] [CrossRef] [PubMed]
- León, I.; Tascón, L.; Cimadevilla, J.M. Age and gender-related differences in a spatial memory task in humans. Behav. Brain Res. 2016, 306, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Boccia, M.; Piccardi, L.; Di Marco, M.; Pizzamiglio, L.; Guariglia, C. Does field independence predict visuo-spatial abilities underpinning human navigation? Behavioural evidence. Exp. Brain Res. 2016, 234, 2799–2807. [Google Scholar] [CrossRef] [PubMed]
- Piccardi, L.; Risetti, M.; Nori, R. Familiarity and environmental representations of a city: A self-report study. Psychol. Rep. 2011, 109, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Nori, R.; Piccardi, L. Familiarity and spatial cognitive style: How important are they for spatial representation? In Spatial Memory: Visuospatial Processes, Cognitive Performance and Developmental Effects; Thomas, J.B., Ed.; Nova Sciences Publiser, Inc: New York, NY, USA, 2011; pp. 123–144. [Google Scholar]
- Cimadevilla, J.M.; Piccardi, L. Spatial skills. In Sex Differences in Neurology and Psychiatry; Lanzenberger, R., Kranz, G.S., Savic, I., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 175, pp. 65–79. [Google Scholar]
- Lawton, C.A. Gender, spatial abilities, and wayfinding. In Handbook of gender research in psychology; Chrisler, J., McCreary, D., Eds.; Springer: New York, NY, USA, 2010; pp. 317–341. [Google Scholar] [CrossRef]
- Zancada-Menendez, C.; Sampedro-Piquero, P.; Lopez, L.; McNamara, T.P.; Zancada-Menendez, C.; Sampedro-Piquero, P.; Lopez, L.; McNamara, T.P. Age and gender differences in spatial perspective taking. Aging Clin. Exp. Res. 2016, 28, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Grön, G.; Wunderlich, A.; Spitzer, M.; Tomczak, R.; Riepe, M.W. Brain activation during human navigation: Gender-different neural networks as substrate of performance. Nat. Neurosci. 2000, 3, 404–408. [Google Scholar] [CrossRef]
- León, I.; Tascón, L.; Ortells-Pareja, J.J.; Cimadevilla, J.M. Virtual reality assessment of walking and nonwalking space in men and women with virtual reality-based tasks. PLoS ONE 2018, 13, 1–15. [Google Scholar] [CrossRef]
- Merriman, N.A.; Ondřej, J.; Roudaia, E.; O’Sullivan, C.; Newell, F.N. Familiar environments enhance object and spatial memory in both younger and older adults. Exp. Brain Res. 2016, 234, 1555–1574. [Google Scholar] [CrossRef]
- Tascón, L.; Castillo, J.; Cimadevilla, J.M. Age-related differences in the elderly in a spatial recognition task. Memory 2019, 27, 1415–1422. [Google Scholar] [CrossRef]
- Wang, L.; Carr, M. Working memory and strategy use contribute to gender differences in spatial ability. Educ. Psychol. 2014, 49, 261–282. [Google Scholar] [CrossRef]
- Castillo Escamilla, J.; Fernández Castro, J.J.; Baliyan, S.; Ortells-Pareja, J.J.; Ortells Rodríguez, J.J.; Cimadevilla, J.M. Allocentric spatial memory performance in a virtual reality-based task is conditioned by visuospatial working memory capacity. Brain Sci. 2020, 10, 552. [Google Scholar] [CrossRef]
- Nori, R.; Piccardi, L.; Pelosi, A.; de Luca, D.; Frasca, F.; Giusberti, F. Perspective changing in WalCT and VR-WalCT: A gender difference study [WalCT–VR-WalCT: Gender differences]. Comput. Hum. Behav. 2015, 53, 316–323. [Google Scholar] [CrossRef]
- Liu, I.; Levy, R.M.; Barton, J.J.S.; Iaria, G. Age and gender differences in various topographical orientation strategies. Brain Res. 2011, 1410, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, G.; Sergi, I.; Iachini, T. Gender differences in remembering and inferring spatial distances. Memory 2008, 16, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Iachini, T.; Ruotolo, F.; Ruggiero, G. The effects of familiarity and gender on spatial representation. J. Environ. Psychol. 2009, 29, 227–234. [Google Scholar] [CrossRef]
- Cheng, C.-H.; Chang, W.-C.; Chang, W.-T. Sex differences in relation to wayfinding strategies, navigational support design, and wayfinding task difficulty. J. Environ. Psychol. 2009, 21, 114–118. [Google Scholar] [CrossRef]
- Terlecki, M.S.; Newcombe, N.S. How important is the digital divide? The relation of computer and videogame usage to gender differences in mental rotation ability. Sex Roles 2005, 53, 433–441. [Google Scholar] [CrossRef]
- De Castell, S.; Larios, H.; Jenson, J. Gender, videogames and navigation in virtual space. Acta Psychol. 2019, 199, 102895. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Men | Women | Total | |
|---|---|---|---|
| WSBRT | n = 48 (47% Spanish) Age = 22.85 ± 2.89 | n = 41 (56% Spanish) Age = 20.56 ± 2.17 | n = 89 (51% Spanish) Age = 21.8 ± 2.82 |
| ASMRT | n = 64 (31% Spanish) Age = 22.67 ± 3.34 | n = 38 (44% Spanish) Age = 21.1 ± 2.6 | n = 102 (36% Spanish) Age = 22.17 ± 3.19 |
| NWSBRT | n = 46 (39% Spanish) Age = 22.42 ± 2.49 | n = 50 (40% Spanish) Age = 21.62 ± 2.82 | n = 96 (39% Spanish) Age = 22.01 ± 2.68 |
| n = 287 (42% Spanish) Age = 22 ± 2.9 | |||
| WSBRT | ASMRT | NWSBRT | |
|---|---|---|---|
| Total number of boxes | 16 | 9 | 16 |
| Locations to memorize | 3, 5 and 7 | 1, 2 and 3 | 3, 5 and 7 |
| Trials of each difficulty level | 10 (9 analyzed) | 4 | 10 (9 analyzed) |
| View | First-person view | First-person view | Survey perspective |
| Participant’s interaction with the context | Active | Passive | Passive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tascón, L.; Di Cicco, C.; Piccardi, L.; Palmiero, M.; Bocchi, A.; Cimadevilla, J.M. Sex Differences in Spatial Memory: Comparison of Three Tasks Using the Same Virtual Context. Brain Sci. 2021, 11, 757. https://doi.org/10.3390/brainsci11060757
Tascón L, Di Cicco C, Piccardi L, Palmiero M, Bocchi A, Cimadevilla JM. Sex Differences in Spatial Memory: Comparison of Three Tasks Using the Same Virtual Context. Brain Sciences. 2021; 11(6):757. https://doi.org/10.3390/brainsci11060757
Chicago/Turabian StyleTascón, Laura, Carmen Di Cicco, Laura Piccardi, Massimiliano Palmiero, Alessia Bocchi, and José Manuel Cimadevilla. 2021. "Sex Differences in Spatial Memory: Comparison of Three Tasks Using the Same Virtual Context" Brain Sciences 11, no. 6: 757. https://doi.org/10.3390/brainsci11060757
APA StyleTascón, L., Di Cicco, C., Piccardi, L., Palmiero, M., Bocchi, A., & Cimadevilla, J. M. (2021). Sex Differences in Spatial Memory: Comparison of Three Tasks Using the Same Virtual Context. Brain Sciences, 11(6), 757. https://doi.org/10.3390/brainsci11060757