
Presence of the Endocannabinoid System in the Inferior Pulvinar of the Vervet Monkey

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
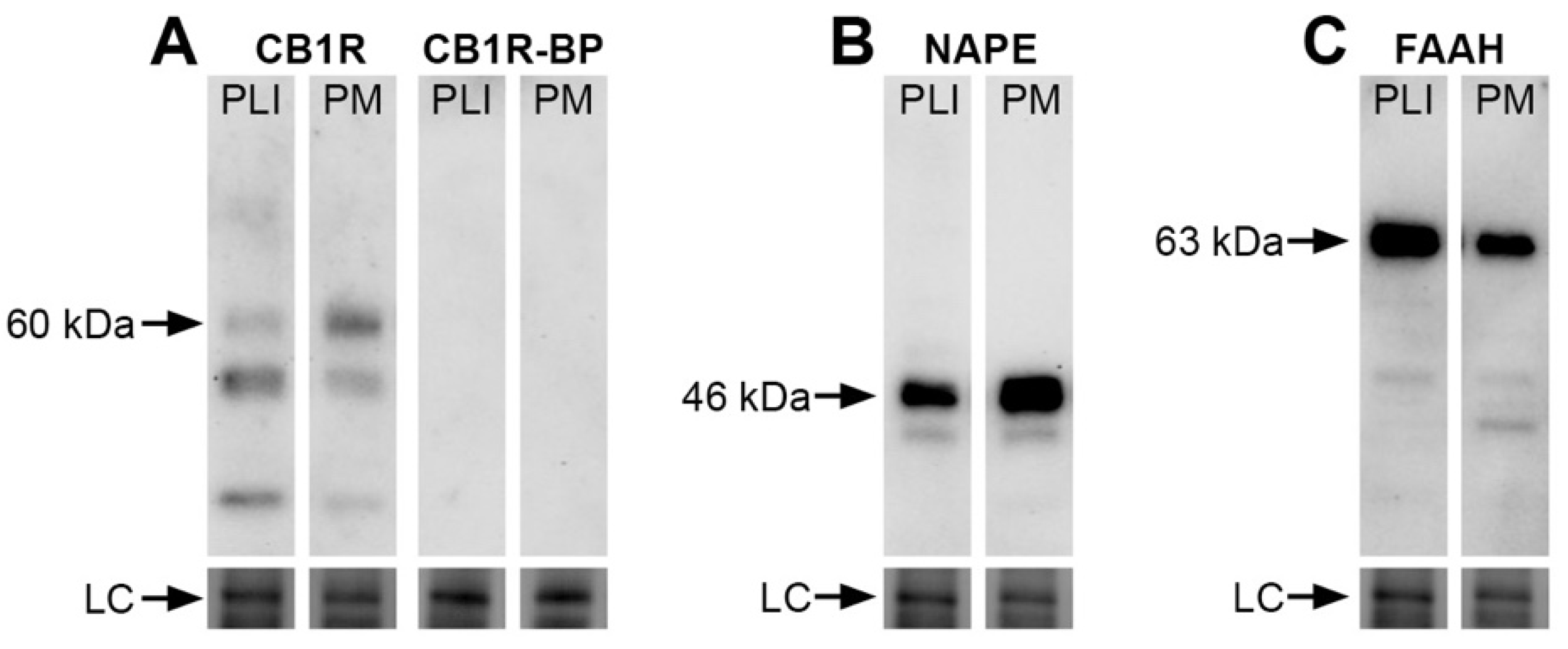
3.1. Western Blot Analysis
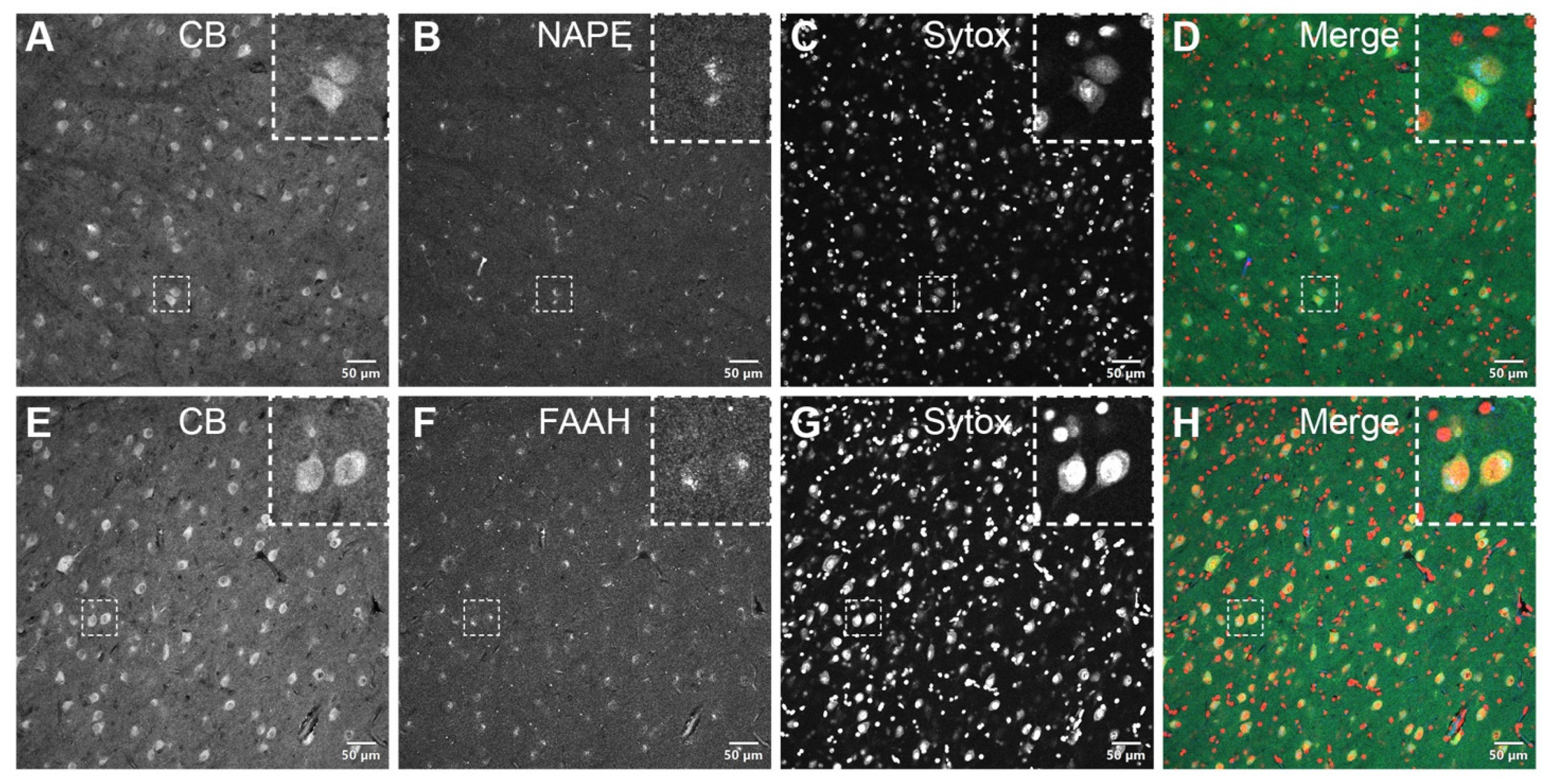
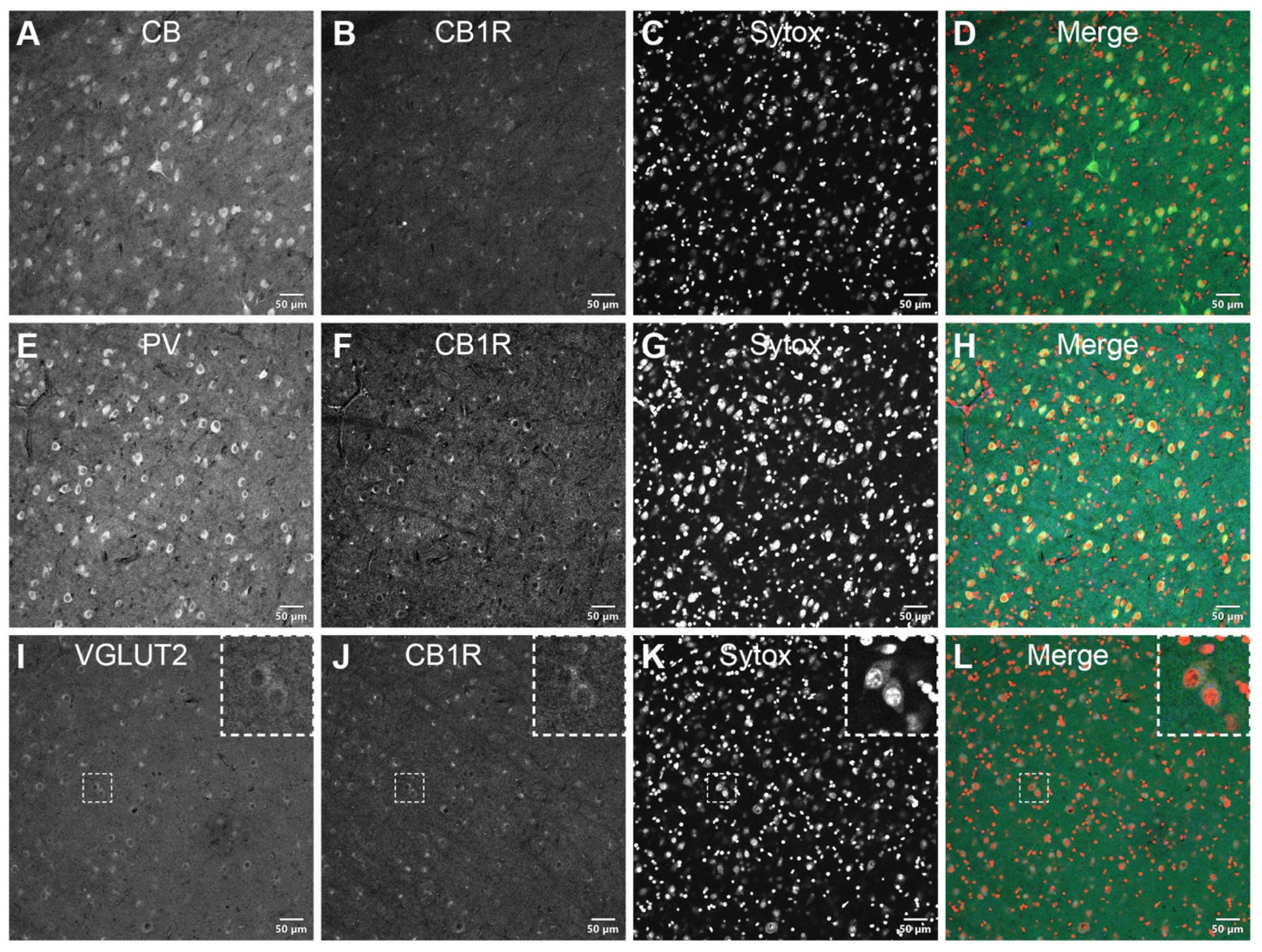
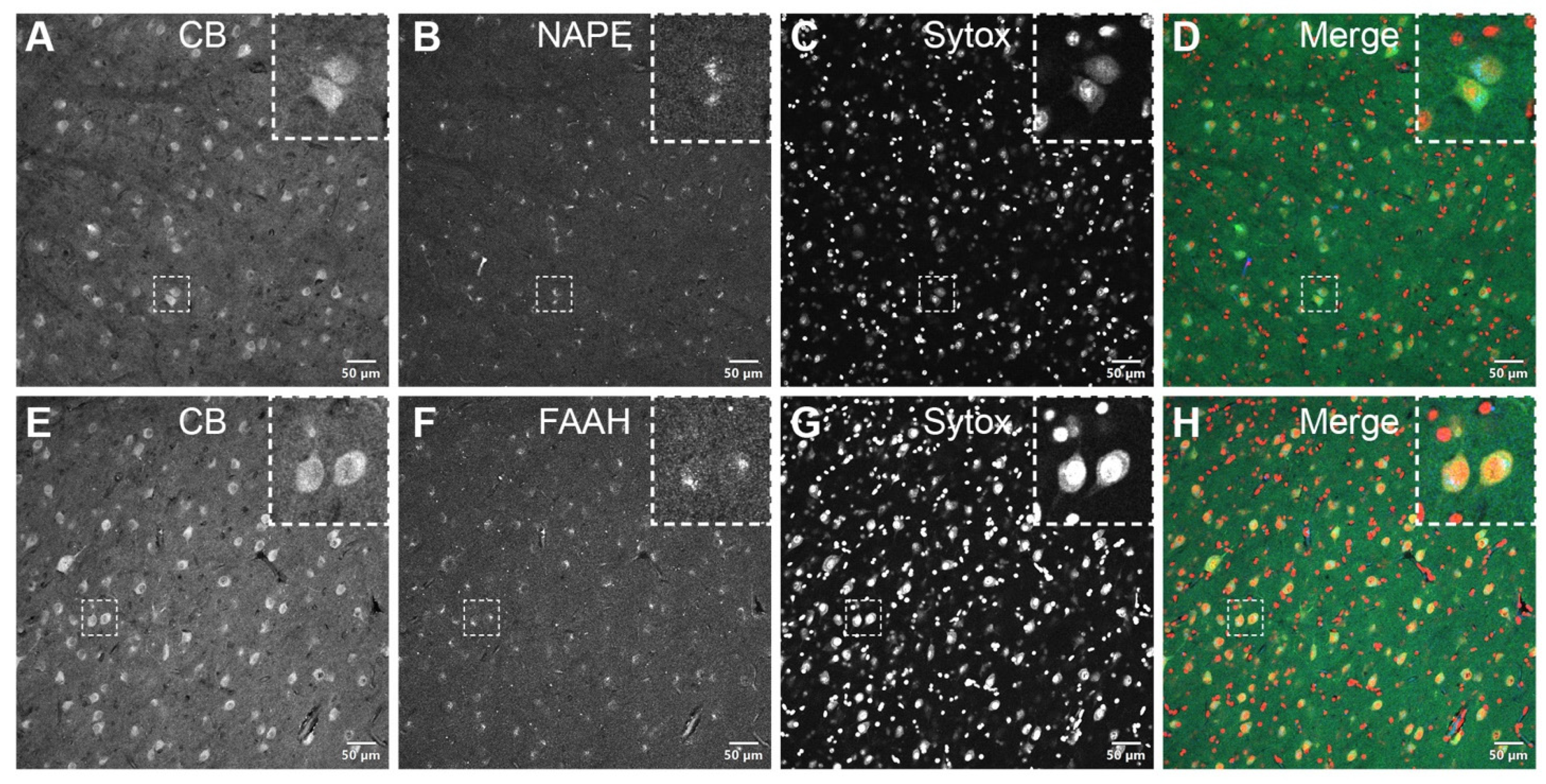
3.2. DAB Immunohistochemistry
3.3. Double Immunofluorescence
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petersen, S.E.; Robinson, D.L.; Morris, J.D. Contributions of the pulvinar to visual spatial attention. Neuropsychologia 1987, 25, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.L.; Petersen, S.E. The pulvinar and visual salience. Trends Neurosci. 1992, 15, 127–132. [Google Scholar] [CrossRef]
- Villeneuve, M.Y.; Kupers, R.; Gjedde, A.; Ptito, M.; Casanova, C. Pattern–motion selectivity in the human pulvinar. Neuroimage 2005, 28, 474–480. [Google Scholar] [CrossRef]
- Cortes, N.; de Souza, B.O.; Casanova, C. Pulvinar Modulates Synchrony across Visual Cortical Areas. Vision 2020, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Stepniewska, I.; Kaas, J.H. Architectonic subdivisions of the inferior pulvinar in New World and Old World monkeys. Vis. Neurosci. 1997, 14, 1043–1060. [Google Scholar] [CrossRef] [PubMed]
- Itaya, S.K.; Van Hoesen, G.W. Retinal projections to the inferior and medial pulvinar nuclei in the Old-World monkey. Brain Res. 1983, 269, 223–230. [Google Scholar] [CrossRef]
- Stepniewska, I.; QI, H.-X.; Kaas, J.H. Projections of the superior colliculus to subdivisions of the inferior pulvinar in New World and Old World monkeys. Vis. Neurosci. 2000, 17, 529–549. [Google Scholar] [CrossRef] [PubMed]
- Sherman, S.M.; Guillery, R. On the actions that one nerve cell can have on another: Distinguishing “drivers” from “modulators”. Proc. Natl. Acad. Sci. USA 1998, 95, 7121–7126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, M.K.; Balaram, P.; Kaas, J.H. Projections of the superior colliculus to the pulvinar in prosimian galagos (Otolemur garnettii) and VGLUT2 staining of the visual pulvinar. J. Comp. Neurol. 2013, 521, 1664–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaas, J.H.; Baldwin, M.K. The evolution of the pulvinar complex in primates and its role in the dorsal and ventral streams of cortical processing. Vision 2020, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Warner, C.E.; Goldshmit, Y.; Bourne, J.A. Retinal afferents synapse with relay cells targeting the middle temporal area in the pulvinar and lateral geniculate nuclei. Front. Neuroanat. 2010, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Mundinano, I.-C.; Kwan, W.C.; Bourne, J.A. Retinotopic specializations of cortical and thalamic inputs to area MT. Proc. Natl. Acad. Sci. USA 2019, 116, 23326–23331. [Google Scholar] [CrossRef] [PubMed]
- Bridge, H.; Leopold, D.A.; Bourne, J.A. Adaptive pulvinar circuitry supports visual cognition. Trends Cogn. Sci. 2016, 20, 146–157. [Google Scholar] [CrossRef] [Green Version]
- Zou, S.; Kumar, U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar]
- Howlett, A.C.; Mukhopadhyay, S. Cellular signal transduction by anandamide and 2-arachidonoylglycerol. Chem. Phys. Lipids 2000, 108, 53–70. [Google Scholar] [CrossRef]
- Di Marzo, V.; Fontana, A.; Cadas, H.; Schinelli, S.; Cimino, G.; Schwartz, J.-C.; Piomelli, D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 1994, 372, 686–691. [Google Scholar] [CrossRef] [Green Version]
- Egertova, M.; Giang, D.K.; Cravatt, B.F.; Elphick, M.R. A new perspective on cannabinoid signalling: Complimentary localization of fatty acid amide hydrolase and the CB1 receptor in rat brain. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 2081–2085. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Ohno-Shosaku, T.; Maejima, T.; Kano, M. Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron 2001, 29, 729–738. [Google Scholar] [CrossRef]
- Heifets, B.D.; Castillo, P.E. Endocannabinoid signaling and long-term synaptic plasticity. Annu. Rev. Physiol. 2009, 71, 283–306. [Google Scholar] [CrossRef] [Green Version]
- Schwitzer, T.; Schwan, R.; Angioi-Duprez, K.; Lalanne, L.; Giersch, A.; Laprevote, V. Cannabis use and human retina: The path for the study of brain synaptic transmission dysfunctions in cannabis users. Neurosci. Biobehav. Rev. 2019, 106, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Peregrina, S.; Ortiz, C.; Casares-López, M.; Jiménez, J.R.; Anera, R.G. Effects of cannabis on visual function and self-perceived visual quality. Sci. Rep. 2021, 11, 1655. [Google Scholar] [CrossRef] [PubMed]
- Bouskila, J.; Burke, M.; Zabouri, N.; Casanova, C.; Ptito, M.; Bouchard, J.-F. Expression and localization of the cannabinoid receptor type 1 and the enzyme fatty acid amide hydrolase in the retina of vervet monkeys. Neuroscience 2012, 202, 117–130. [Google Scholar] [CrossRef]
- Javadi, P.; Bouskila, J.; Bouchard, J.-F.; Ptito, M. The endocannabinoid system within the dorsal lateral geniculate nucleus of the vervet monkey. Neuroscience 2015, 288, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Kucera, R.; Bouskila, J.; Toutoungy, M.; Dow, R.; Palmour, R.; Ptito, M.; Bouchard, J.-F. AB007. Expression and localization of CB1R, NAPE-PLD, and FAAH in the primary visual cortex of vervet monkeys. Ann. Eye Sci. 2020, 4, AB007. [Google Scholar] [CrossRef]
- Kucera, R.; Bouskila, J.; Elkrief, L.; Fink-Jensen, A.; Palmour, R.; Bouchard, J.-F.; Ptito, M. Expression and localization of CB1R, NAPE-PLD, and FAAH in the vervet monkey nucleus accumbens. Sci. Rep. 2018, 8, 8689. [Google Scholar] [CrossRef]
- Burke, M.W.; Zangenehpour, S.; Ptito, M. Brain banking: Making the most of your research specimens. JoVE (J. Vis. Exp.) 2009, e1260. [Google Scholar] [CrossRef] [Green Version]
- Perry, V.; Cowey, A. Retinal ganglion cells that project to the superior colliculus and pretectum in the macaque monkey. Neuroscience 1984, 12, 1125–1137. [Google Scholar] [CrossRef]
- Kwan, W.C.; Mundinano, I.C.; de Souza, M.J.; Lee, S.C.; Martin, P.R.; Grünert, U.; Bourne, J.A. Unravelling the subcortical and retinal circuitry of the primate inferior pulvinar. J. Comp. Neurol. 2019, 527, 558–576. [Google Scholar] [CrossRef]
- Masri, R.A.; Percival, K.A.; Koizumi, A.; Martin, P.R.; Grünert, U. Survey of retinal ganglion cell morphology in marmoset. J. Comp. Neurol. 2019, 527, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Bittar, R.; Ptito, M.; Faubert, J.; Dumoulin, S.; Ptito, A. Activation of the remaining hemisphere following stimulation of the blind hemifield in hemispherectomized subjects. Neuroimage 1999, 10, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Ptito, A.; Leh, S.E. Neural substrates of blindsight after hemispherectomy. Neuroscientist 2007, 13, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.; MacLean, M.W.; Hadid, V.; Lazzouni, L.; Nguyen, D.K.; Tremblay, J.; Dehaes, M.; Lepore, F. Neuronal mechanisms of motion detection underlying blindsight assessed by functional magnetic resonance imaging (fMRI). Neuropsychologia 2019, 128, 187–197. [Google Scholar] [CrossRef]
- Baldwin, M.K.; Balaram, P.; Kaas, J.H. The evolution and functions of nuclei of the visual pulvinar in primates. J. Comp. Neurol. 2017, 525, 3207–3226. [Google Scholar] [CrossRef] [PubMed]
- Warner, C.E.; Kwan, W.C.; Wright, D.; Johnston, L.A.; Egan, G.F.; Bourne, J.A. Preservation of vision by the pulvinar following early-life primary visual cortex lesions. Curr. Biol. 2015, 25, 424–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, J.A.; Rosa, M.G. Hierarchical development of the primate visual cortex, as revealed by neurofilament immunoreactivity: Early maturation of the middle temporal area (MT). Cereb. Cortex 2006, 16, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, C.E.; Kwan, W.C.; Bourne, J.A. The early maturation of visual cortical area MT is dependent on input from the retinorecipient medial portion of the inferior pulvinar. J. Neurosci. 2012, 32, 17073–17085. [Google Scholar] [CrossRef] [PubMed]
- Mundinano, I.-C.; Fox, D.M.; Kwan, W.C.; Vidaurre, D.; Teo, L.; Homman-Ludiye, J.; Goodale, M.A.; Leopold, D.A.; Bourne, J.A. Transient visual pathway critical for normal development of primate grasping behavior. Proc. Natl. Acad. Sci. USA 2018, 115, 1364–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, J.; Braddick, O. Visual development. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 173, pp. 121–142. [Google Scholar]
- Mason, A.; Braddick, O.; Wattam-Bell, J. Motion coherence thresholds in infants—Different tasks identify at least two distinct motion systems. Vis. Res. 2003, 43, 1149–1157. [Google Scholar] [CrossRef] [Green Version]
- Soares, S.C.; Maior, R.S.; Isbell, L.A.; Tomaz, C.; Nishijo, H. Fast detector/first responder: Interactions between the superior colliculus-pulvinar pathway and stimuli relevant to primates. Front. Neurosci. 2017, 11, 67. [Google Scholar] [CrossRef]
- Elorette, C.; Forcelli, P.A.; Saunders, R.C.; Malkova, L. Colocalization of tectal inputs with amygdala-projecting neurons in the macaque pulvinar. Front. Neural Circuits 2018, 12, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elphick, M.R. The evolution and comparative neurobiology of endocannabinoid signalling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3201–3215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, R.J. The endocannabinoid system of animals. Animals 2019, 9, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikulskaya, E.; Martin, F.H. Contrast sensitivity and motion discrimination in cannabis users. Psychopharmacology 2018, 235, 2459–2469. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Anstice, N.S.; Jacobs, R.J.; LaGasse, L.L.; Lester, B.M.; Wouldes, T.A.; Thompson, B. Prenatal exposure to recreational drugs affects global motion perception in preschool children. Sci. Rep. 2015, 5, 16921. [Google Scholar] [CrossRef] [Green Version]
- Craft, R.M.; Marusich, J.A.; Wiley, J.L. Sex differences in cannabinoid pharmacology: A reflection of differences in the endocannabinoid system? Life Sci. 2013, 92, 476–481. [Google Scholar] [CrossRef] [Green Version]
- Homman-Ludiye, J.; Bourne, J.A. The medial pulvinar: Function, origin and association with neurodevelopmental disorders. J. Anat. 2019, 235, 507–520. [Google Scholar] [CrossRef]
- Koller, K.; Rafal, R.D.; Platt, A.; Mitchell, N.D. Orienting toward threat: Contributions of a subcortical pathway transmitting retinal afferents to the amygdala via the superior colliculus and pulvinar. Neuropsychologia 2019, 128, 78–86. [Google Scholar] [CrossRef]
- Platt, B.; Kamboj, S.; Morgan, C.J.; Curran, H.V. Processing dynamic facial affect in frequent cannabis-users: Evidence of deficits in the speed of identifying emotional expressions. Drug Alcohol. Depend. 2010, 112, 27–32. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Immunogen | Source | Working Dilution | RRID |
|---|---|---|---|---|
| CB | Recombinant protein specific to amino terminus of human Calbindin D-28K | Cell Signaling Technology, Danvers, MA, USA | DAB 1:150 | AB_2687400 |
| CB | Calbindin D-28k purified from chicken gut | Swant, Marly, Fribourg, Switzerland | IF 1:150 | AB_10000347 |
| CB1R | Synthetic peptide corresponding with the C terminal aa 461–472 of human CB1R | Abcam, Cambridge, UK | IF 1:100, WB 1:1000 | AB_447623 |
| FAAH | Synthetic peptide corresponding with the aa 561–579 of rat FAAH | Cayman Chemical, Ann Arbor, MI, USA | IF 1:100, WB 1:1000 | AB_10078701 |
| NAPE-PLD | Synthetic peptide from human NAPE-PLD aa 159–172 | Cayman Chemical, Ann Arbor, MI, USA | IF 1:100, WB 1:1000 | AB_10507996 |
| PV | Parvalbumin purified from carp muscle | Swant, Marly, Fribourg, Switzerland | IF 1:150 | AB_10000343 |
| VGLUT2 | Recombinant full-length rat Vesicular Glutamate Transporter 2 (VGLUT2) | Millipore, Burlington, MA, USA | IF 1:100 | AB_287552 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Micaelo-Fernandes, C.; Bouskila, J.; Bouchard, J.-F.; Ptito, M. Presence of the Endocannabinoid System in the Inferior Pulvinar of the Vervet Monkey. Brain Sci. 2021, 11, 770. https://doi.org/10.3390/brainsci11060770
Micaelo-Fernandes C, Bouskila J, Bouchard J-F, Ptito M. Presence of the Endocannabinoid System in the Inferior Pulvinar of the Vervet Monkey. Brain Sciences. 2021; 11(6):770. https://doi.org/10.3390/brainsci11060770
Chicago/Turabian StyleMicaelo-Fernandes, Catarina, Joseph Bouskila, Jean-François Bouchard, and Maurice Ptito. 2021. "Presence of the Endocannabinoid System in the Inferior Pulvinar of the Vervet Monkey" Brain Sciences 11, no. 6: 770. https://doi.org/10.3390/brainsci11060770
APA StyleMicaelo-Fernandes, C., Bouskila, J., Bouchard, J. -F., & Ptito, M. (2021). Presence of the Endocannabinoid System in the Inferior Pulvinar of the Vervet Monkey. Brain Sciences, 11(6), 770. https://doi.org/10.3390/brainsci11060770