
The Paternal Brain in Action: A Review of Human Fathers’ fMRI Brain Responses to Child-Related Stimuli

 ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
1.1. The Key Role of Fathering Behavior
1.2. Neurobiology of Fathering in Animal Models
1.3. Challenges in Exploring the Human Paternal Brain
1.4. The Present Study
2. Materials and Methods
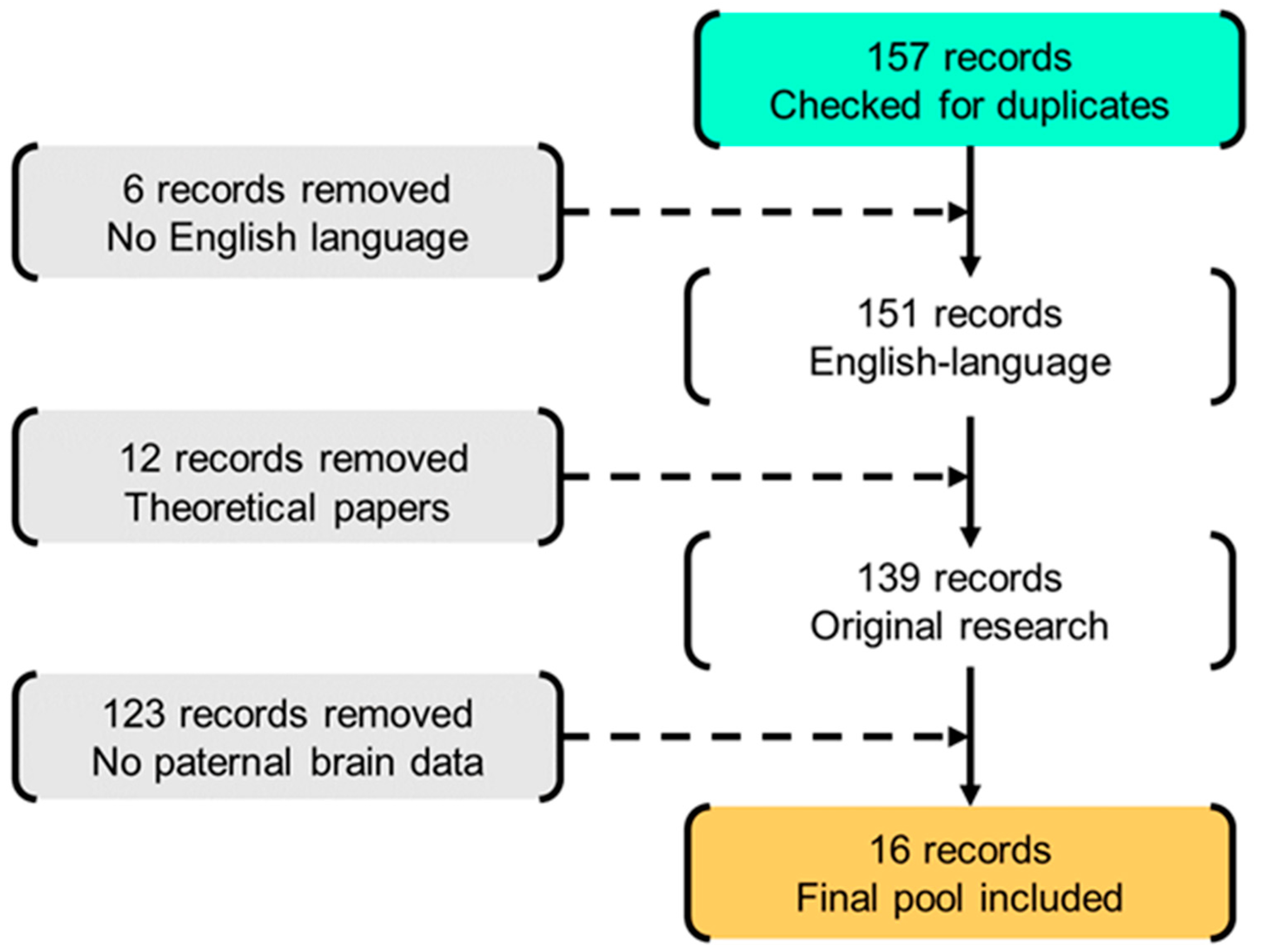
2.1. Literature Search
2.2. Selection of Retrieved Records
2.3. Data Abstracting
2.4. Data Synthesis
3. Results
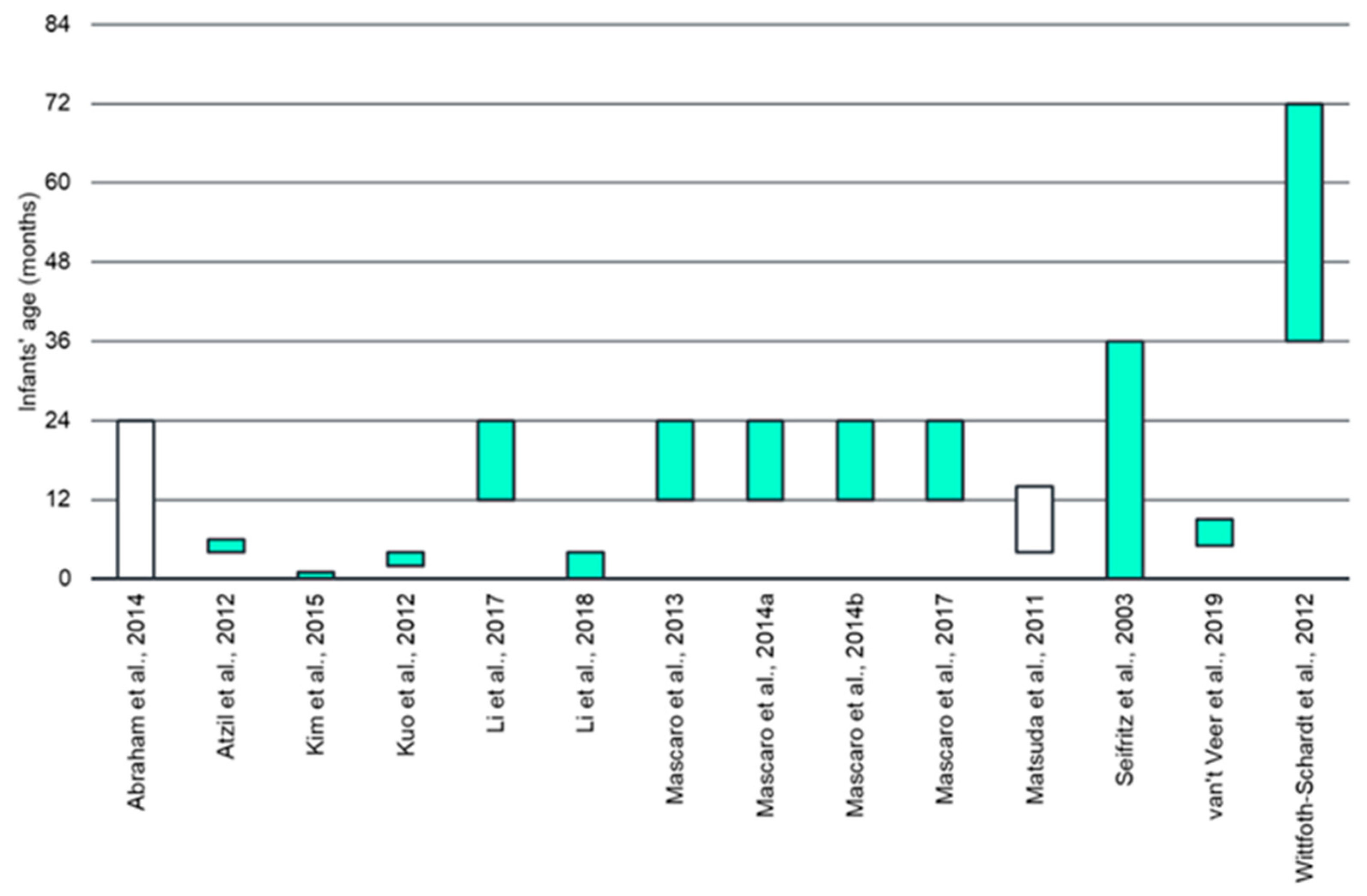
3.1. Participant Characteristics
3.2. Methodology Review
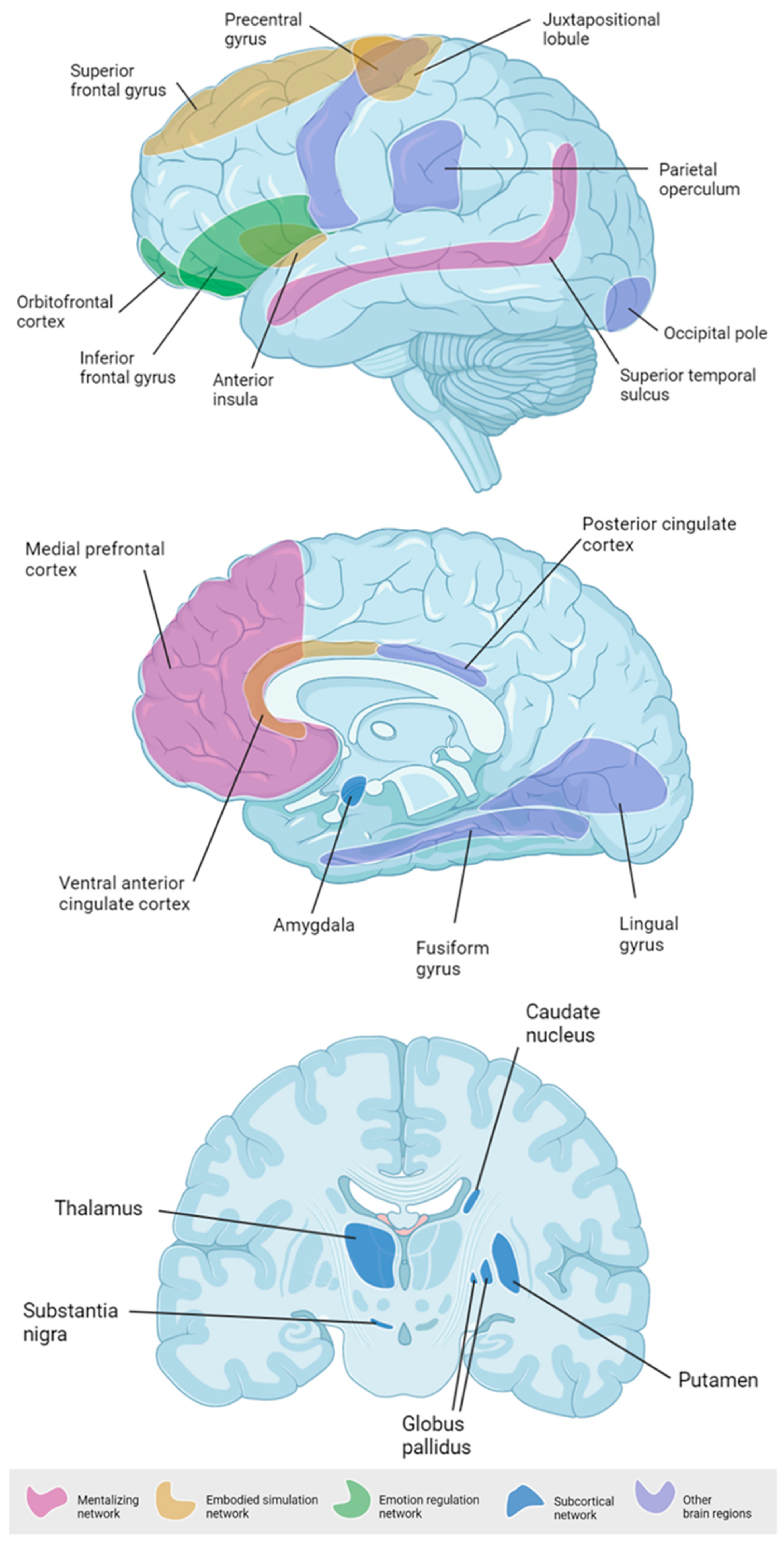
3.3. Emerging Paternal Brain Areas
3.3.1. Mentalizing Network
3.3.2. Embodied Simulation Network
3.3.3. Emotion Regulation Network
3.3.4. Subcortical Parenting Network
3.3.5. Other Brain Regions
4. Discussion
4.1. Preliminary Methodological Issues
4.2. Toward an Integrated View of the Human Paternal Brain
4.3. Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McGill, B.S. Navigating New Norms of Involved Fatherhood. J. Fam. Issues 2014, 35, 1089–1106. [Google Scholar] [CrossRef]
- Bianchi, S.M. Family Change and Time Allocation in American Families. Ann. Am. Acad. Pol. Soc. Sci. 2011, 638, 21–44. [Google Scholar] [CrossRef]
- Brown, G.L.; Mangelsdorf, S.C.; Shigeto, A.; Wong, M.S. Associations between father involvement and father-child attachment security: Variations based on timing and type of involvement. J. Fam. Psychol. 2018, 32, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Lee, S.J. Caring is masculine: Stay-at-home fathers and masculine identity. Psychol. Men Masc. 2018, 19, 47–58. [Google Scholar] [CrossRef]
- Sigurdardottir, H.M.; Garðarsdóttir, Ó. Backlash in gender equality? Fathers’ parental leave during a time of economic crisis. J. Eur. Soc. Policy 2018, 28, 342–356. [Google Scholar] [CrossRef]
- Berik, G.; Kongar, E. Time Allocation of Married Mothers and Fathers in Hard Times: The 2007–09 US Recession. Fem. Econ. 2013, 19, 208–237. [Google Scholar] [CrossRef]
- Almqvist, A.L.; Duvander, A.Z. Changes in gender equality? Swedish fathers’ parental leave, division of childcare and housework1. J. Fam. Stud. 2014, 20, 19–27. [Google Scholar] [CrossRef]
- Brandth, B.; Kvande, E. Fathers and flexible parental leave. Work. Employ. Soc. 2016, 30, 275–290. [Google Scholar] [CrossRef]
- Gettler, L.T. Becoming DADS: Considering the role of cultural context and developmental plasticity for paternal socioendocrinology. Curr. Anthropol. 2016, 57, S38–S51. [Google Scholar] [CrossRef]
- Hunter, S.C.; Riggs, D.W.; Augoustinos, M. Hegemonic masculinity versus a caring masculinity: Implications for understanding primary caregiving fathers. Soc. Personal. Psychol. Compass 2017, 11, e12307. [Google Scholar] [CrossRef]
- Gimenez-Nadal, J.I.; Sevilla, A. Trends in time allocation: A cross-country analysis. Eur. Econ. Rev. 2012, 56, 1338–1359. [Google Scholar] [CrossRef]
- Cano, T.; Perales, F.; Baxter, J. A Matter of Time: Father Involvement and Child Cognitive Outcomes. J. Marriage Fam. 2019, 81, 164–184. [Google Scholar] [CrossRef]
- Palkovitz, R. Expanding Our Focus From Father Involvement to Father–Child Relationship Quality. J. Fam. Theory Rev. 2019, 11, 576–591. [Google Scholar] [CrossRef]
- Rogers, F.D.; Bales, K.L. Mothers, Fathers, and Others: Neural Substrates of Parental Care. Trends Neurosci. 2019, 42, 552–562. [Google Scholar] [CrossRef]
- Storey, A.E.; Alloway, H.; Walsh, C.J. Dads: Progress in understanding the neuroendocrine basis of human fathering behavior. Horm. Behav. 2020, 119, 104660. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; McCoy, D.C.; Yousafzai, A.K.; Salhi, C.; Fink, G. Paternal stimulation and early child development in low-and middle-income countries. Pediatrics 2016, 138, e20161357. [Google Scholar] [CrossRef] [PubMed]
- Bakermans-Kranenburg, M.J.; Lotz, A.; Alyousefi-van Dijk, K.; IJzendoorn, M. Birth of a Father: Fathering in the First 1000 Days. Child Dev. Perspect. 2019, 13, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Gettler, L.T.; Boyette, A.H.; Rosenbaum, S. Broadening Perspectives on the Evolution of Human Paternal Care and Fathers’ Effects on Children. Annu. Rev. Anthropol. 2020, 49, 141–160. [Google Scholar] [CrossRef]
- Lucassen, N.; Tharner, A.; Van IJzendoorn, M.H.; Bakermans-Kranenburg, M.J.; Volling, B.L.; Verhulst, F.C.; Lambregtse-Van den Berg, M.P.; Tiemeier, H. The Association Between Paternal Sensitivity and Infant-Father Attachment Security: A Meta-Analysis of Three Decades of Research. J. Fam. Psychol. 2011, 25, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Craig, L.; Mullan, K. How Mothers and Fathers Share Childcare. Am. Sociol. Rev. 2011, 76, 834–861. [Google Scholar] [CrossRef]
- Amodia-Bidakowska, A.; Laverty, C.; Ramchandani, P.G. Father-child play: A systematic review of its frequency, characteristics and potential impact on children’s development. Dev. Rev. 2020, 57, 100924. [Google Scholar] [CrossRef]
- Leech, K.A.; Salo, V.C.; Rowe, M.L.; Cabrera, N.J. Father input and child vocabulary development: The importance of Wh questions and clarification requests. Semin. Speech Lang. 2013, 34, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, N.J.; Shannon, J.D.; La Taillade, J.J. Predictors of co-parenting in Mexican American families and direct effects on parenting and child social emotional development. Infant Ment. Health J. 2009, 30, 523–548. [Google Scholar] [CrossRef] [PubMed]
- Majdandžić, M.; Möller, E.L.; De Vente, W.; Bögels, S.M.; Van Den Boom, D.C. Fathers’ challenging parenting behavior prevents social anxiety development in their 4-year-old children: A longitudinal observational study. J. Abnorm. Child Psychol. 2014, 42, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Braun, K.; Champagne, F.A. The neural mechanisms and consequences of paternal caregiving. Nat. Rev. Neurosci. 2019, 20, 205–224. [Google Scholar] [CrossRef] [PubMed]
- de Jong, T.R.; Chauke, M.; Harris, B.N.; Saltzman, W. From here to paternity: Neural correlates of the onset of paternal behavior in California mice (Peromyscus californicus). Horm. Behav. 2008, 56, 220–231. [Google Scholar] [CrossRef]
- Bales, K.L.; Saltzman, W. Fathering in rodents: Neurobiological substrates and consequences for offspring. Horm. Behav. 2016, 77, 249–259. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Williams, J.R.; Slotnick, B.M.; Carter, C.S. Olfactory bulbectomy decreases social behavior in male prairie voles (M. ochrogaster). Physiol. Behav. 1994, 55, 885–889. [Google Scholar] [CrossRef]
- Kozorovitskiy, Y.; Hughes, M.; Lee, K.; Gould, E. Fatherhood affects dendritic spines and vasopressin V1a receptors in the primate prefrontal cortex. Nat. Neurosci. 2006, 9, 1094–1095. [Google Scholar] [CrossRef] [PubMed]
- Rilling, J.K.; Mascaro, J.S. The neurobiology of fatherhood. Curr. Opin. Psychol. 2017, 15, 26–32. [Google Scholar] [CrossRef]
- Witteman, J.; Van IJzendoorn, M.H.; Rilling, J.K.; Bos, P.A.; Schiller, N.O.; Bakermans-Kranenburg, M.J. Towards a neural model of infant cry perception. Neurosci. Biobehav. Rev. 2019, 99, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339. [Google Scholar] [CrossRef]
- De Pisapia, N.; Bornstein, M.H.; Rigo, P.; Esposito, G.; De Falco, S.; Venuti, P. Sex differences in directional brain responses to infant hunger cries. Neuroreport 2013, 24, 142–146. [Google Scholar] [CrossRef]
- Mascaro, J.S.; Hackett, P.D.; Rilling, J.K. Differential neural responses to child and sexual stimuli in human fathers and non-fathers and their hormonal correlates. Psychoneuroendocrinology 2014, 46, 153–163. [Google Scholar] [CrossRef]
- Wittfoth-Schardt, D.; Grunding, J.; Wittfoth, M.; Lanfermann, H.; Heinrichs, M.; Domes, G.; Buchheim, A.; Gundel, H.; Waller, C. Oxytocin modulates neural reactivity to children’s faces as a function of social salience. Neuropsychopharmacology 2012, 37, 1799–1807. [Google Scholar] [CrossRef]
- Kuo, P.X.; Carp, J.; Light, K.C.; Grewen, K.M. Neural responses to infants linked with behavioral interactions and testosterone in fathers. Biol. Psychol. 2012, 91, 302–306. [Google Scholar] [CrossRef]
- Seifritz, E.; Esposito, F.; Neuhoff, J.G.; Lüthi, A.; Mustovic, H.; Dammann, G.; Von Bardeleben, U.; Radue, E.W.; Cirillo, S.; Tedeschi, G.; et al. Differential sex-independent amygdala response to infant crying and laughing in parents versus nonparents. Biol. Psychiatry 2003, 54, 1367–1375. [Google Scholar] [CrossRef]
- Mascaro, J.S.; Hackett, P.D.; Gouzoules, H.; Lori, A.; Rilling, J.K. Behavioral and genetic correlates of the neural response to infant crying among human fathers. Soc. Cogn. Affect. Neurosci. 2014, 9, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Van’t Veer, A.E.; Thijssen, S.; Witteman, J.; Van Ijzendoorn, M.H.; Bakermans-Kranenburg, M.J. Exploring the neural basis for paternal protection: An investigation of the neural response to infants in danger. Soc. Cogn. Affect. Neurosci. 2019, 14, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Mascaro, J.S.; Hackett, P.D.; Rilling, J.K. Testicular volume is inversely correlated with nurturing-related brain activity in human fathers. Proc. Natl. Acad. Sci. USA 2013, 110, 15746–15751. [Google Scholar] [CrossRef] [PubMed]
- Atzil, S.; Hendler, T.; Zagoory-Sharon, O.; Winetraub, Y.; Feldman, R. Synchrony and specificity in the maternal and the paternal brain: Relations to oxytocin and vasopressin. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 798–811. [Google Scholar] [CrossRef]
- Li, T.; Chen, X.; Mascaro, J.; Haroon, E.; Rilling, J.K. Intranasal oxytocin, but not vasopressin, augments neural responses to toddlers in human fathers. Horm. Behav. 2017, 93, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Horta, M.; Mascaro, J.S.; Bijanki, K.; Arnal, L.H.; Adams, M.; Barr, R.G.; Rilling, J.K. Explaining individual variation in paternal brain responses to infant cries. Physiol. Behav. 2018, 193, 43–54. [Google Scholar] [CrossRef]
- Abraham, E.; Hendler, T.; Shapira-Lichter, I.; Kanat-Maymon, Y.; Zagoory-Sharon, O.; Feldman, R. Father’s brain is sensitive to childcare experiences. Proc. Natl. Acad. Sci. USA 2014, 111, 9792–9797. [Google Scholar] [CrossRef] [PubMed]
- Mascaro, J.S.; Rentscher, K.E.; Hackett, P.D.; Mehl, M.R.; Rilling, J.K. Child gender influences paternal behavior, language, and brain function. Behav. Neurosci. 2017, 131, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Rigo, P.; Leckman, J.F.; Mayes, L.C.; Cole, P.M.; Feldman, R.; Swain, J.E. A prospective longitudinal study of perceived infant outcomes at 18-24 months: Neural and psychological correlates of parental thoughts and actions assessed during the first month postpartum. Front. Psychol. 2015, 6, 1772. [Google Scholar] [CrossRef]
- Matsuda, Y.T.; Ueno, K.; Waggoner, R.A.; Erickson, D.; Shimura, Y.; Tanaka, K.; Cheng, K.; Mazuka, R. Processing of infant-directed speech by adults. Neuroimage 2011, 54, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, S.; Van ’t Veer, A.E.; Witteman, J.; Meijer, W.M.; van IJzendoorn, M.H.; Bakermans-Kranenburg, M.J. Effects of vasopressin on neural processing of infant crying in expectant fathers. Horm. Behav. 2018, 103, 19–27. [Google Scholar] [CrossRef]
- Abraham, E.; Hendler, T.; Zagoory-Sharon, O.; Feldman, R. Interoception sensitivity in the parental brain during the first months of parenting modulates children’s somatic symptoms six years later: The role of oxytocin. Int. J. Psychophysiol. 2019, 136, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Demetriou, L.; Kowalczyk, O.S.; Tyson, G.; Bello, T.; Newbould, R.D.; Wall, M.B. A comprehensive evaluation of increasing temporal resolution with multiband-accelerated protocols and effects on statistical outcome measures in fMRI. Neuroimage 2018, 176, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, N.K. What we can do and what we cannot do with fMRI. Nature 2008, 453, 869–878. [Google Scholar] [CrossRef]
- Petersen, S.E.; Dubis, J.W. The mixed block/event-related design. Neuroimage 2012, 62, 1177–1184. [Google Scholar] [CrossRef]
- Fedorenko, E.; Hsieh, P.J.; Nieto-Castañón, A.; Whitfield-Gabrieli, S.; Kanwisher, N. New method for fMRI investigations of language: Defining ROIs functionally in individual subjects. J. Neurophysiol. 2010, 104, 1177–1194. [Google Scholar] [CrossRef]
- Sohn, W.S.; Yoo, K.; Lee, Y.B.; Seo, S.W.; Na, D.L.; Jeong, Y. Influence of ROI selection on resting functional connectivity: An individualized approach for resting fMRI analysis. Front. Neurosci. 2015, 9, 280. [Google Scholar] [CrossRef]
- Belin, P.; Zatorre, R.J.; Lafallie, P.; Ahad, P.; Pike, B. Voice-selective areas in human auditory cortex. Nature 2000, 403, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, D.; Japee, S.; Rauth, L.; Ungerleider, L.G. The superior temporal sulcus is causally connected to the amygdala: A combined TBS-fMRI study. J. Neurosci. 2017, 37, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Deen, B.; Koldewyn, K.; Kanwisher, N.; Saxe, R. Functional organization of social perception and cognition in the superior temporal sulcus. Cereb. Cortex 2015, 25, 4596–4609. [Google Scholar] [CrossRef] [PubMed]
- Pelphrey, K.A.; Morris, J.P. Brain mechanisms for interpreting the actions of others from biological-motion cues. Curr. Dir. Psychol. Sci. 2006, 15, 136–140. [Google Scholar] [CrossRef]
- Ciaramidaro, A.; Adenzato, M.; Enrici, I.; Erk, S.; Pia, L.; Bara, B.G.; Walter, H. The intentional network: How the brain reads varieties of intentions. Neuropsychologia 2007, 45, 3105–3113. [Google Scholar] [CrossRef]
- Van Overwalle, F. Social cognition and the brain: A meta-analysis. Hum. Brain Mapp. 2009, 30, 829–858. [Google Scholar] [CrossRef]
- Corradi-Dell’Acqua, C.; Hofstetter, C.; Vuilleumier, P. Cognitive and affective theory of mind share the same local patterns of activity in posterior temporal but not medial prefrontal cortex. Soc. Cogn. Affect. Neurosci. 2014, 9, 1175–1184. [Google Scholar] [CrossRef]
- Lundy, B.L. Father- and mother-infant face-to-face interactions: Differences in mind-related comments and infant attachment. Infant Behav. Dev. 2003, 26, 200–212. [Google Scholar] [CrossRef]
- Gallese, V.; Sinigaglia, C. What is so special about embodied simulation? Trends Cogn. Sci. 2011, 15, 512–519. [Google Scholar] [CrossRef]
- Tsakiris, M.; Schütz-Bosbach, S.; Gallagher, S. On agency and body-ownership: Phenomenological and neurocognitive reflections. Conscious. Cogn. 2007, 16, 645–660. [Google Scholar] [CrossRef]
- Craig, A.D. Emotional moments across time: A possible neural basis for time perception in the anterior insula. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Montirosso, R.; McGlone, F. The body comes first. Embodied reparation and the co-creation of infant bodily-self. Neurosci. Biobehav. Rev. 2020, 113, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Afif, A.; Bouvier, R.; Buenerd, A.; Trouillas, J.; Mertens, P. Development of the human fetal insular cortex: Study of the gyration from 13 to 28 gestational weeks. Brain Struct. Funct. 2007, 212, 335–346. [Google Scholar] [CrossRef]
- Fotopoulou, A.; Tsakiris, M. Mentalizing homeostasis: The social origins of interoceptive inference. Neuropsychoanalysis 2017, 19, 3–28. [Google Scholar] [CrossRef]
- Della Longa, L.; Gliga, T.; Farroni, T. Tune to touch: Affective touch enhances learning of face identity in 4-month-old infants. Dev. Cogn. Neurosci. 2019, 35, 42–46. [Google Scholar] [CrossRef]
- Crucianelli, L.; Wheatley, L.; Filippetti, M.L.; Jenkinson, P.M.; Kirk, E.; Fotopoulou, A. (Katerina) The mindedness of maternal touch: An investigation of maternal mind-mindedness and mother-infant touch interactions. Dev. Cogn. Neurosci. 2019, 35, 47–56. [Google Scholar] [CrossRef]
- Provenzi, L.; Rosa, E.; Visintin, E.; Mascheroni, E.; Guida, E.; Cavallini, A.; Montirosso, R. Understanding the role and function of maternal touch in children with neurodevelopmental disabilities. Infant Behav. Dev. 2020, 58, 101420. [Google Scholar] [CrossRef]
- Feldman, R.; Gordon, I.; Schneiderman, I.; Weisman, O.; Zagoory-Sharon, O. Natural variations in maternal and paternal care are associated with systematic changes in oxytocin following parent-infant contact. Psychoneuroendocrinology 2010, 35, 1133–1141. [Google Scholar] [CrossRef]
- Colonnesi, C.; Zeegers, M.A.J.; Majdandžić, M.; van Steensel, F.J.A.; Bögels, S.M. Fathers’ and Mothers’ Early Mind-Mindedness Predicts Social Competence and Behavior Problems in Childhood. J. Abnorm. Child Psychol. 2019, 47, 1421–1435. [Google Scholar] [CrossRef] [PubMed]
- Möller, E.L.; Majdandžić, M.; Bögels, S.M. Fathers’ versus mothers’ social referencing signals in relation to infant anxiety and avoidance: A visual cliff experiment. Dev. Sci. 2014, 17, 1012–1028. [Google Scholar] [CrossRef]
- Goldin, P.R.; McRae, K.; Ramel, W.; Gross, J.J. The Neural Bases of Emotion Regulation: Reappraisal and Suppression of Negative Emotion. Biol. Psychiatry 2008, 63, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Welborn, B.L.; Papademetris, X.; Reis, D.L.; Rajeevan, N.; Bloise, S.M.; Gray, J.R. Variation in orbitofrontal cortex volume: Relation to sex, emotion regulation and affect. Soc. Cogn. Affect. Neurosci. 2009, 4, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Tabei, K.I. Inferior Frontal Gyrus Activation Underlies the Perception of Emotions, while Precuneus Activation Underlies the Feeling of Emotions during Music Listening. Behav. Neurol. 2015, 2015. [Google Scholar] [CrossRef]
- Grecucci, A.; Giorgetta, C.; Van’t Wout, M.; Bonini, N.; Sanfey, A.G. Reappraising the ultimatum: An fMRI study of emotion regulation and decision making. Cereb. Cortex 2013, 23, 399–410. [Google Scholar] [CrossRef]
- Bornstein, M.H.; Suwalsky, J.T.D.; Breakstone, D.A. Emotional relationships between mothers and infants: Knowns, unknowns, and unknown unknowns. Dev. Psychopathol. 2012, 24, 113. [Google Scholar] [CrossRef] [PubMed]
- Leerkes, E.M.; Su, J.; Calkins, S.D.; Supple, A.J.; O’Brien, M. Pathways by which mothers’ physiological arousal and regulation while caregiving predict sensitivity to infant distress. J. Fam. Psychol. 2016, 30, 769–779. [Google Scholar] [CrossRef]
- Smith, A.P.R.; Stephan, K.E.; Rugg, M.D.; Dolan, R.J. Task and content modulate amygdala-hippocampal connectivity in emotional retrieval. Neuron 2006, 49, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Musser, E.D.; Kaiser-Laurent, H.; Ablow, J.C. The neural correlates of maternal sensitivity: An fMRI study. Dev. Cogn. Neurosci. 2012, 2, 428–436. [Google Scholar] [CrossRef]
- Gallese, V. Bodily selves in relation: Embodied simulation as second-person perspective on intersubjectivity. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130177. [Google Scholar] [CrossRef]
- Maffongelli, L.; Ferrari, E.; Bartoli, E.; Campus, C.; Olivier, E.; Fadiga, L.; D’Ausilio, A. Role of sensorimotor areas in early detection of motor errors: An EEG and TMS study. Behav. Brain Res. 2020, 378, 112248. [Google Scholar] [CrossRef]
- Dandolo, L.C.; Schwabe, L. Time-dependent motor memory representations in prefrontal cortex. Neuroimage 2019, 197, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Lindner, I.; Schain, C.; Echterhoff, G. Other-self confusions in action memory: The role of motor processes. Cognition 2016, 149, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Feldman, R.; Mayes, L.C.; Eicher, V.; Thompson, N.; Leckman, J.F.; Swain, J.E. Breastfeeding, brain activation to own infant cry, and maternal sensitivity. J. Child Psychol. Psychiatry Allied Discip. 2011, 52, 907–915. [Google Scholar] [CrossRef]
- Wright, D.B.; Laurent, H.K.; Ablow, J.C. Mothers Who Were Neglected in Childhood Show Differences in Neural Response to Their Infant’s Cry. Child Maltreat. 2017, 22, 158–166. [Google Scholar] [CrossRef]
- Kim, P.; Leckman, J.F.; Mayes, L.C.; Newman, M.A.; Feldman, R.; Swain, J.E. Perceived quality of maternal care in childhood and structure and function of mothers’ brain. Dev. Sci. 2010, 13, 662–673. [Google Scholar] [CrossRef]
- Elliott, M.L.; Knodt, A.R.; Ireland, D.; Morris, M.L.; Poulton, R.; Ramrakha, S.; Sison, M.L.; Moffitt, T.E.; Caspi, A.; Hariri, A.R. What Is the Test-Retest Reliability of Common Task-Functional MRI Measures? New Empirical Evidence and a Meta-Analysis. Psychol. Sci. 2020, 31, 792–806. [Google Scholar] [CrossRef]
- Lindstedt, J.; Korja, R.; Vilja, S.; Ahlqvist-Björkroth, S. Fathers’ prenatal attachment representations and the quality of father–child interaction in infancy and toddlerhood. J. Fam. Psychol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rilling, J.K.; Young, L.J. The biology of mammalian parenting and its effect on offspring social development. Science 2014, 345, 771–776. [Google Scholar] [CrossRef]
- Swain, J.E. Baby stimuli and the parent brain: Functional neuroimaging of the neural substrates of parent-infant attachment. Psychiatry 2008, 5, 28–36. [Google Scholar]
- Perone, S.; Gartstein, M.A.; Anderson, A.J. Dynamics of frontal alpha asymmetry in mother-infant dyads: Insights from the Still Face Paradigm. Infant Behav. Dev. 2020, 61, 101500. [Google Scholar] [CrossRef] [PubMed]
- Kinreich, S.; Djalovski, A.; Kraus, L.; Louzoun, Y.; Feldman, R. Brain-to-Brain Synchrony during Naturalistic Social Interactions. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Azhari, A.; Leck, W.Q.; Gabrieli, G.; Bizzego, A.; Rigo, P.; Setoh, P.; Bornstein, M.H.; Esposito, G. Parenting Stress Undermines Mother-Child Brain-to-Brain Synchrony: A Hyperscanning Study. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Study | Journal | Fathers N | Other Group(s) | Stimulus Type | Brain NetWork(s) § |
|---|---|---|---|---|---|
| Abraham et al., 2014 [44] | PNAS | 21 | Mothers; homosexual fathers | Visual | M, ES |
| Atzil et al., 2012 [41] | JAACAP | 15 | Mothers | Visual | M, ES |
| De Pisapia et al., 2013 [33] | Neuroreport | 5 | Mothers; non-parent adults | Sound | M, O |
| Kim et al., 2015 [46] | Front Psychol | 19 | Mothers | Sound | SC |
| Kuo et al., 2012 [36] | Biol Psychol | 10 | N/A | Visual | ER, SC, O |
| Li et al., 2017 [42] | Horm Behav | 31 | N/A | Sound | M, SC |
| Li et al., 2018 [43] | Physiol Behav | 42 | N/A | Sound | M, ER |
| Mascaro et al., 2013 [40] | PNAS | 70 | N/A | Visual | M, |
| Mascaro et al., 2014a [38] | SCAN | 36 | N/A | Sound | ES, ER, SC |
| Mascaro et al., 2014b [34] | PNEC | 88 | Non-father men | Visual | ES, ER, O |
| Mascaro et al., 2017 [45] | Behav Neurosci | 69 | N/A | Visual | ER |
| Matsuda et al., 2011 [47] | Neuroimage | 15 | Mothers; non-parent men | Sound | NSA |
| Seifritz et al., 2003 [37] | Biol Psychiatry | 10 | Mothers; non-parent women | Sound | SC |
| Thijssen et al., 2018 [48] | Horm Behav | 25 | N/A | Sound | O |
| van ‘t Veer et al., 2019 [39] | SCAN | 21 | N/A | Visual | ES, O |
| Wittfoth-Schardt et al., 2012 [35] | Neuropsychoph | 19 | N/A | Visual | SC |
| Stimulus Condition | Tasks | References |
|---|---|---|
| Auditory stimuli | ||
| Crying/laughing | 6 | De Pisapia et al., 2013 [33]; Kim et al., 2015 [46]; Li et al., 2017 [42]; Mascaro et al., 2014a [38]; Seifritz et al., 2003 [37]; Thijssen et al., 2018 [48] |
| Infant-directed speech | 1 | Matsuda et al., 2011 [47] |
| Baby vocalization | 1 | Mascaro et al., 2014a [38] |
| Visual stimuli | ||
| Face pictures | 5 | Li et al., 2017 [42]; Mascaro et al., 2014b [34]; Mascaro et al., 2013 [40]; Wittfoth-Schardt et al., 2012 [35]; Mascaro et al., 2017 [45] |
| Father-infant videotapes | 3 | Abraham et al., 2015 [44]; Atzil et al., 2012 [41]; Kuo et al., 2012 [36] |
| Infant alone videotapes | 2 | Atzil et al., 2012 [41]; van’t Veer et al., 2019 [39] |
| Study | Field Strength | TR [ms] | TE [ms] | Slice Thickness (Slice Gap) [mm] | In Plane Resolution [mm2] | Number of Slices |
|---|---|---|---|---|---|---|
| Abraham et al., 2014 [44] | 3 T | 3000 | 35 | 3 | 2.33 × 2.33 | 39 |
| Atzil et al., 2012 [41] | 3 T | 3000 | 35 | 3 | 1.56 × 1.56 | 39 |
| De Pisapia et al., 2013 [33] | 4 T | 2000 | 33 | 3 (0.45) | 3 × 3 | 34 |
| Kim et al., 2015 [46] | 3 T | 2000 | 30 | 5 | 3.125 × 3.125 | 30 |
| Kuo et al., 2012 [36] | 3 T | 1500 | 30 | 4 | 4 × 4 | 28 |
| Li et al., 2017 [42] | 3 T | 2000 | 28 | 2.5 (1.05) | 3.5 × 3.5 | 34 |
| Li et al., 2018 [43] | 3 T | 2380 | 30 | 3 | 3 × 3 | 38 |
| Mascaro et al., 2013 [40] | 3 T | 2000 | 28 | 2.5 (1.05) | 3.5 × 3.5 | 34 |
| Mascaro et al., 2014a [38] | 3 T | 2000 | 28 | 2.5 (1.05) | 3.5 × 3.5 | 34 |
| Mascaro et al., 2014b [34] | 3 T | 2000 | 28 | 2.5 (1.05) | 3.5 × 3.5 | 34 |
| Mascaro et al., 2017 [45] | 3 T | 2000 | 28 | 2.5 (1.05) | 3.5 × 3.5 | 34 |
| Matsuda et al., 2011 [47] | 4 T | 2600 | 25 | 5 | 3.75 × 3.75 | 23 |
| Seifritz et al., 2003 [37] | 1.5 T | 2675 | / | 4 | 2.8 × 2.8 | 25 |
| Thijssen et al., 2018 [48] | 3 T | 2200 | 30 | 2.75 (0.275) | 2.75 × 2.75 | 38 |
| van ‘t Veer et al., 2019 [39] | 3 T | 2200 | 30 | 2.75 (0.275) | 2.75 × 2.75 | 38 |
| Wittfoth-Schardt et al., 2012 [35] | 1.5 T | 2100 | 30 | 3 (0.75) | 3 × 3 | 35 |
| Study | Voxel Level | ROI Purpose | ROI Selection | Summary of Selected ROIs | 2nd Level Analysis | Software |
|---|---|---|---|---|---|---|
| Abraham et al., 2014 [44] | Yes | Sampling ROI | Mixed | Amyg, STS, TP, vmPFC, FP | VOXEL: random effect models; ROI: correlations | Brain Voyager |
| Atzil et al., 2012 [41] | Yes | - | - | - | Random effect models; correlation | Not specified |
| De Pisapia et al., 2013 [33] | Yes | Inclusive Mask | Data-driven | STL, dmPFC, PCC | Random effect models | Brain Voyager |
| Kim et al., 2015 [46] | Yes | Inclusive Mask | Theory-driven | Limbic system, striatum, midbrain regions (STG, Put, SN) | Random effect models; Mediation models | SPM8 |
| Kuo et al., 2012 [36] | Yes | Sampling ROI | Mixed | OFC, SFG, mFG, IFG, Cau, SPL, mTL, CG | Random effect models; correlations | SPM8 |
| Li et al., 2017 [42] | Yes | Sampling ROI | Theory-driven | Cau, ACC | Random effect models | FSL |
| Li et al., 2018 [43] | Yes | Sampling ROI | Data-driven | 15 ROIs for different analyses | Mixed effect models; correlation analysis | FSL |
| Mascaro et al., 2013 [40] | Yes (exploratory) | Sampling ROI | Theory-driven | VTA, SN | VOXEL/ROI: Correlation analysis | Brain Voyager |
| Mascaro et al., 2014a [38] | Yes | Sampling ROI | Theory-driven | AI | VOXEL: Random effect models, correlation analysis; ROI: correlation analysis | Brain voyager |
| Mascaro et al., 2014b [34] | Yes | Sampling ROI | Data-driven | mFG, AI, vmPFC, mOFC | VOXEL: random effect model, correlation analysis. ROIs: t-test, correlation analysis. | Brain Voyager |
| Mascaro et al., 2014b [34] | Yes | Sampling ROI | Data-driven | Hyp, Accu, Cau, SN/VTA | VOXEL: random effect model, correlation analysis. ROIs: t-test, correlation analysis. | Brain Voyager |
| Mascaro et al., 2017 [45] | Yes | - | - | - | ANCOVA; correlation analysis | Brain Voyager |
| Matsuda et al., 2011 [47] | Yes | Sampling ROI | Data-driven | pIFG, SPL, ACC | VOXEL: Random effect model; ROIs: ANOVA, correlation analysis | Brain voyager |
| Seifritz et al., 2003 [37] | Yes | - | - | - | VOXEL: t-test; ROIs: | Brain voyager |
| Thijssen et al., 2018 [48] | Yes | Inclusive Mask | Data-driven | Infant cry network | 2 × 2 models and post-hoc t-test, F-test | FSL |
| van ‘t Veer et al., 2019 [39] | Yes (exploratory) | Sampling ROI | Theory-driven | Amyg | VOXEL: mixed effect model. ROI: GLM repeated measure model (SPSS) | FSL + SPSS |
| Wittfoth-Schardt et al., 2012 [35] | Yes | Sampling ROI | Theory-driven | GP, Hip | VOXEL: random effect model; ROIs: ANOVA, connectivity analysis | SPM8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Provenzi, L.; Lindstedt, J.; De Coen, K.; Gasparini, L.; Peruzzo, D.; Grumi, S.; Arrigoni, F.; Ahlqvist-Björkroth, S. The Paternal Brain in Action: A Review of Human Fathers’ fMRI Brain Responses to Child-Related Stimuli. Brain Sci. 2021, 11, 816. https://doi.org/10.3390/brainsci11060816
Provenzi L, Lindstedt J, De Coen K, Gasparini L, Peruzzo D, Grumi S, Arrigoni F, Ahlqvist-Björkroth S. The Paternal Brain in Action: A Review of Human Fathers’ fMRI Brain Responses to Child-Related Stimuli. Brain Sciences. 2021; 11(6):816. https://doi.org/10.3390/brainsci11060816
Chicago/Turabian StyleProvenzi, Livio, Johanna Lindstedt, Kris De Coen, Linda Gasparini, Denis Peruzzo, Serena Grumi, Filippo Arrigoni, and Sari Ahlqvist-Björkroth. 2021. "The Paternal Brain in Action: A Review of Human Fathers’ fMRI Brain Responses to Child-Related Stimuli" Brain Sciences 11, no. 6: 816. https://doi.org/10.3390/brainsci11060816
APA StyleProvenzi, L., Lindstedt, J., De Coen, K., Gasparini, L., Peruzzo, D., Grumi, S., Arrigoni, F., & Ahlqvist-Björkroth, S. (2021). The Paternal Brain in Action: A Review of Human Fathers’ fMRI Brain Responses to Child-Related Stimuli. Brain Sciences, 11(6), 816. https://doi.org/10.3390/brainsci11060816