
Microglial Metamorphosis in Three Dimensions in Virus Limbic Encephalitis: An Unbiased Pictorial Representation Based on a Stereological Sampling Approach of Surveillant and Reactive Microglia

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Experimental Procedures
2.1. Animals and Infection
2.2. Perfusion and Microtomy
2.3. Morphometry Based on 3D Reconstruction
2.4. Statistical Analysis of Morphometry
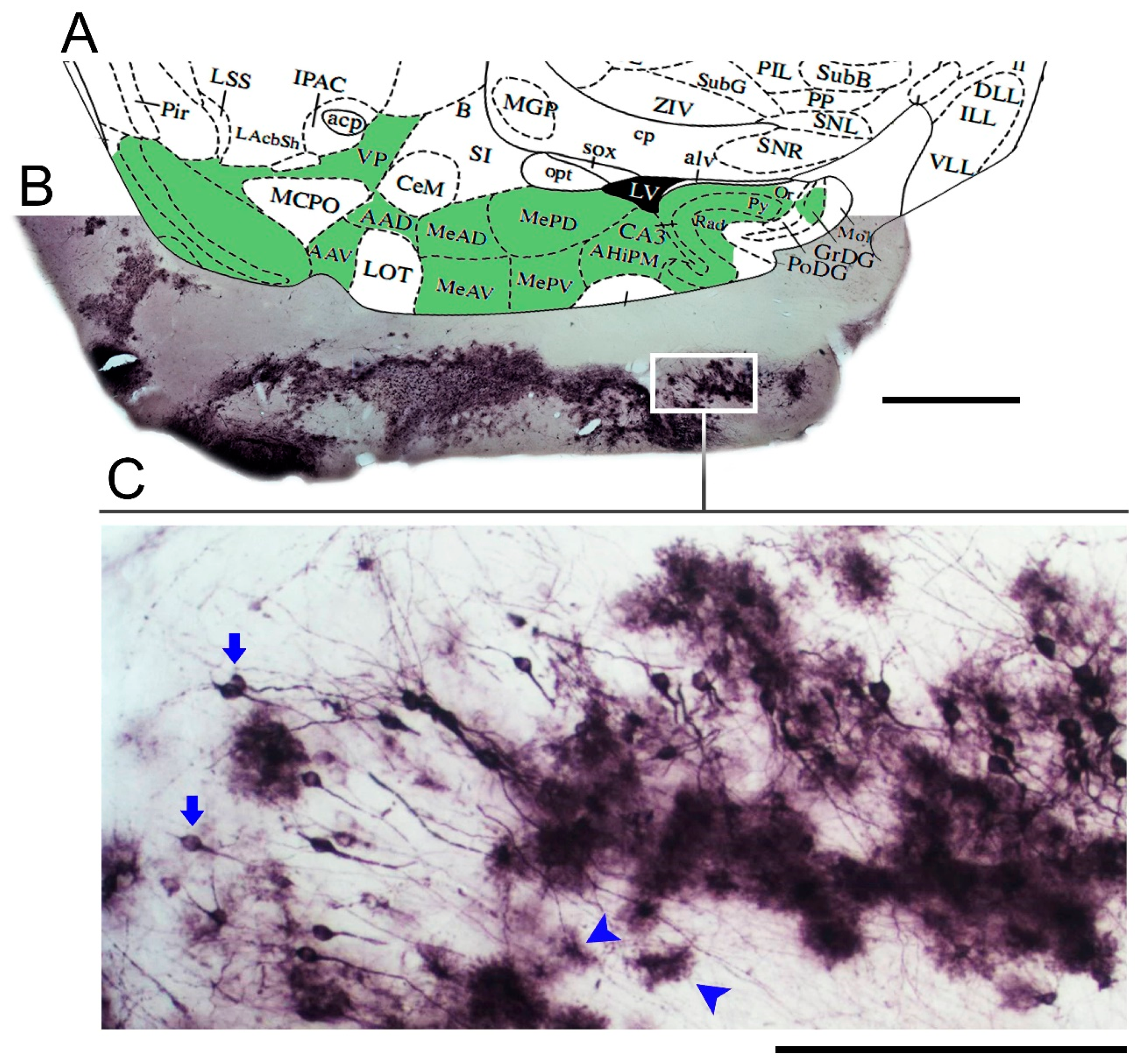
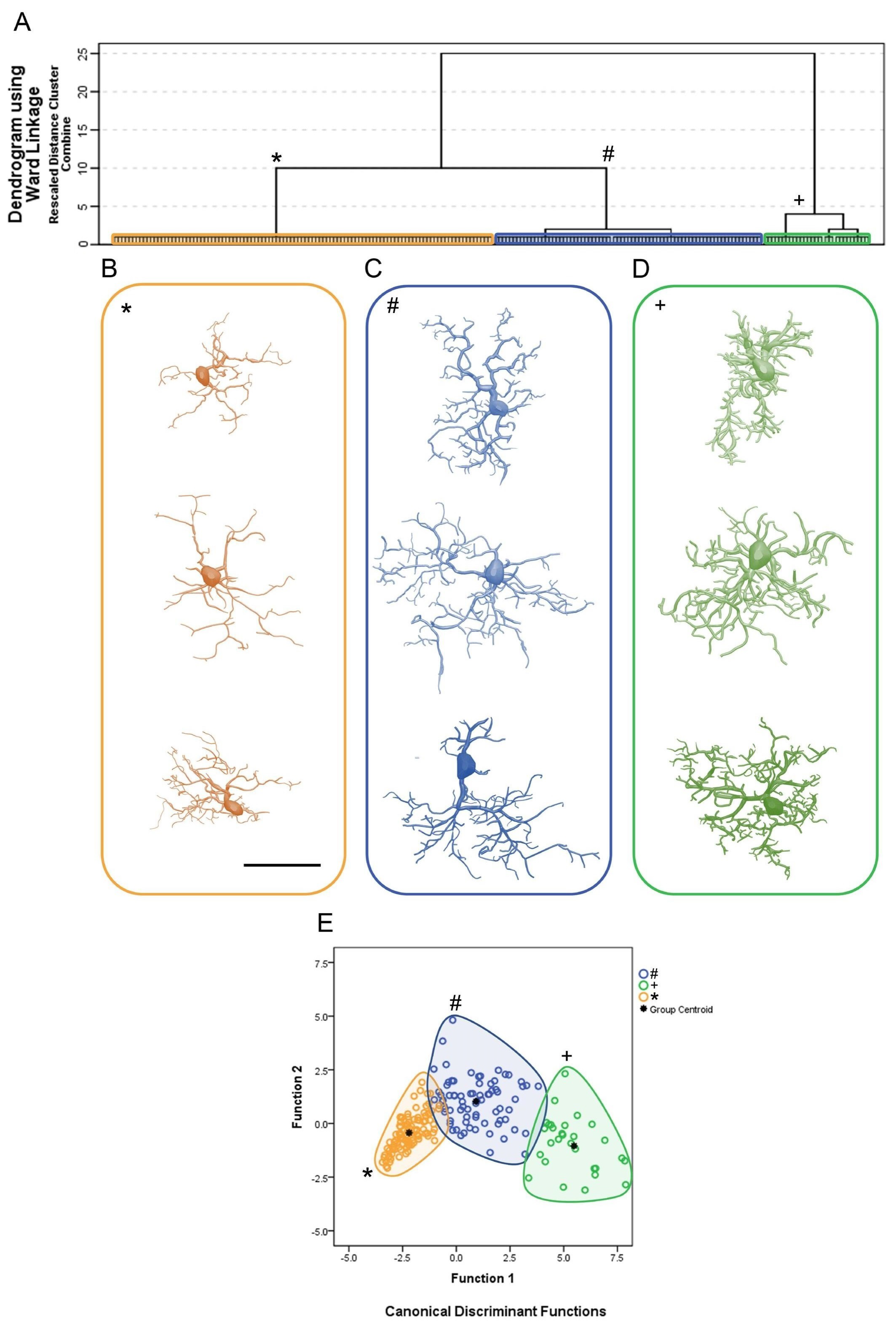
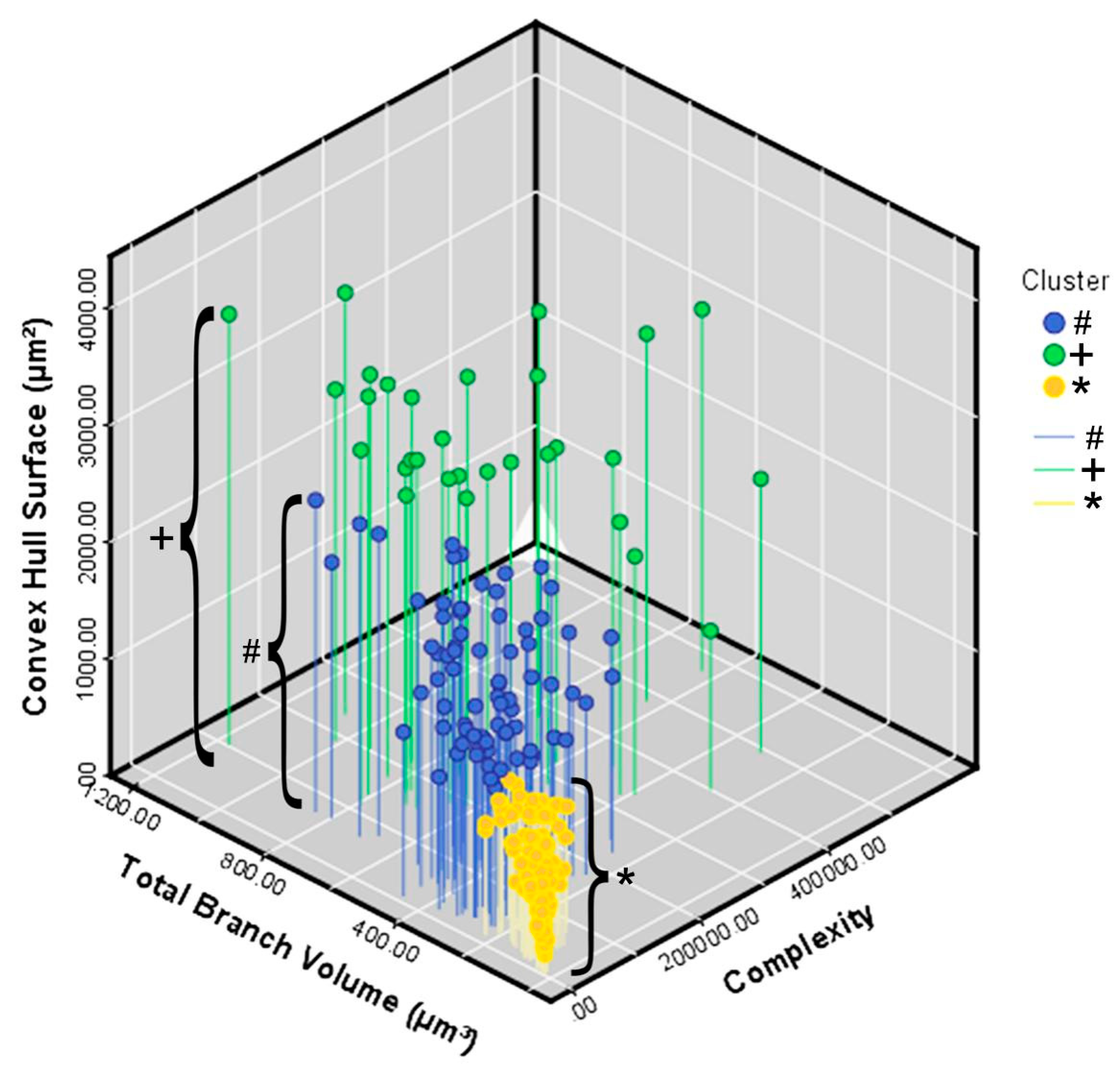
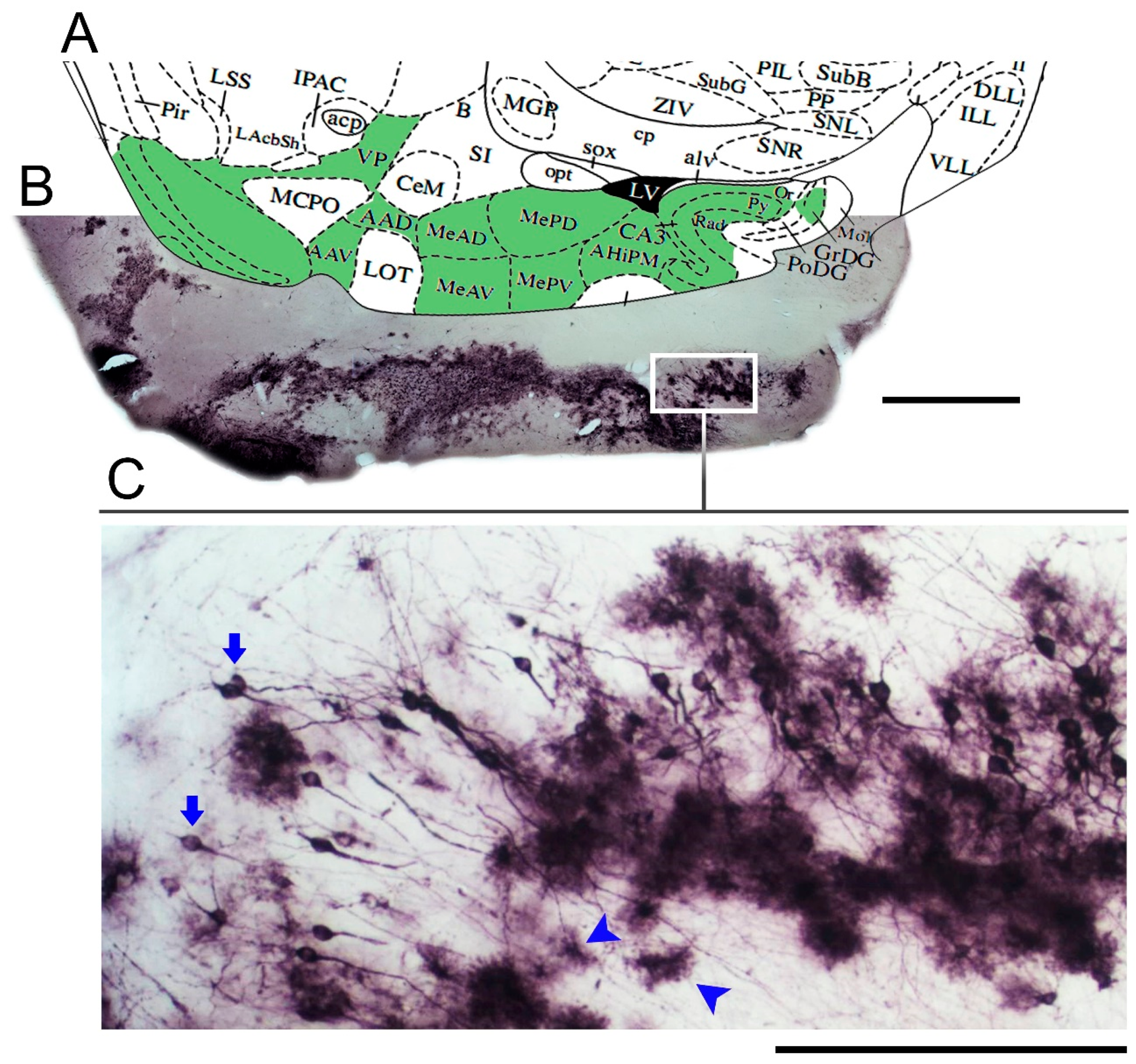
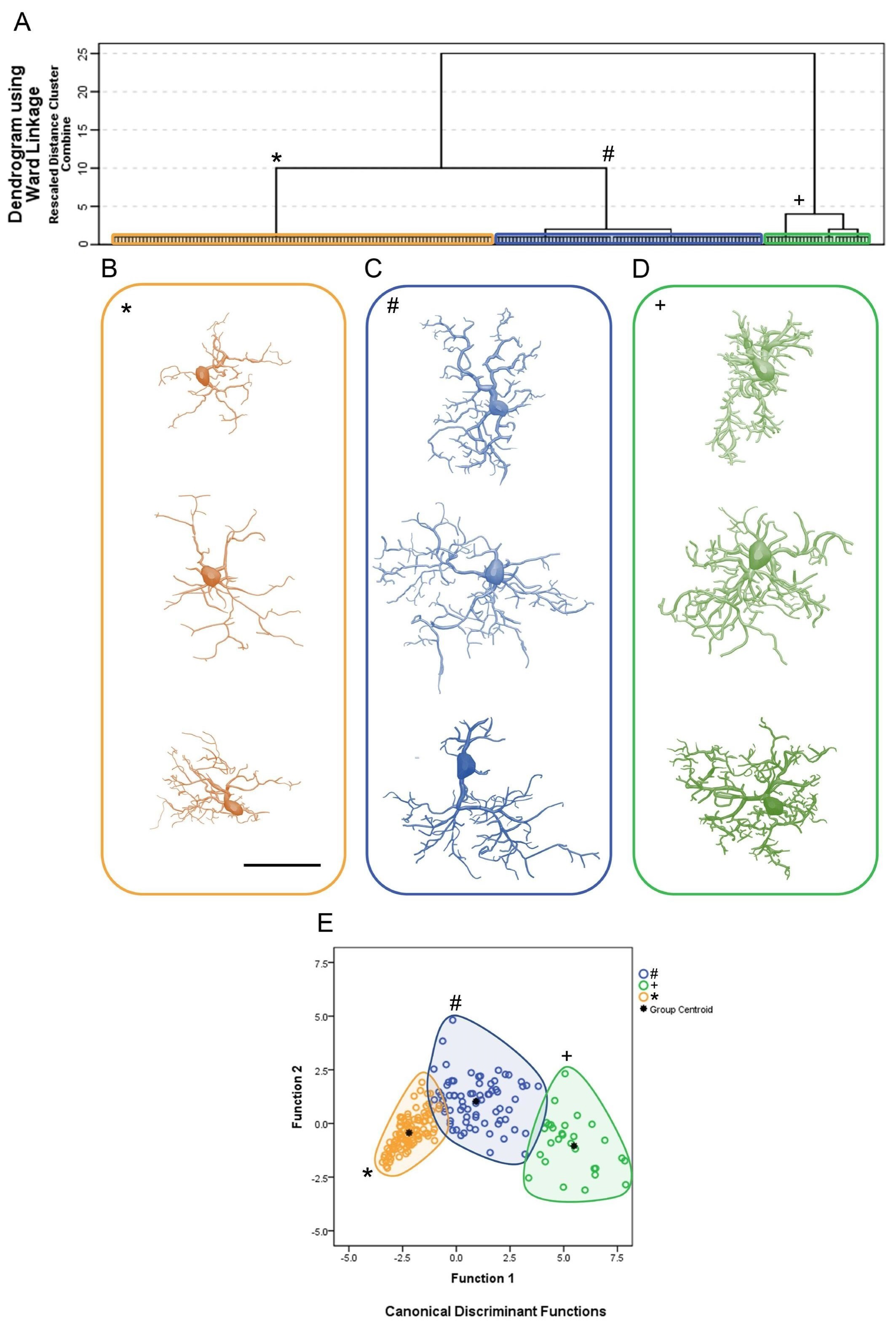
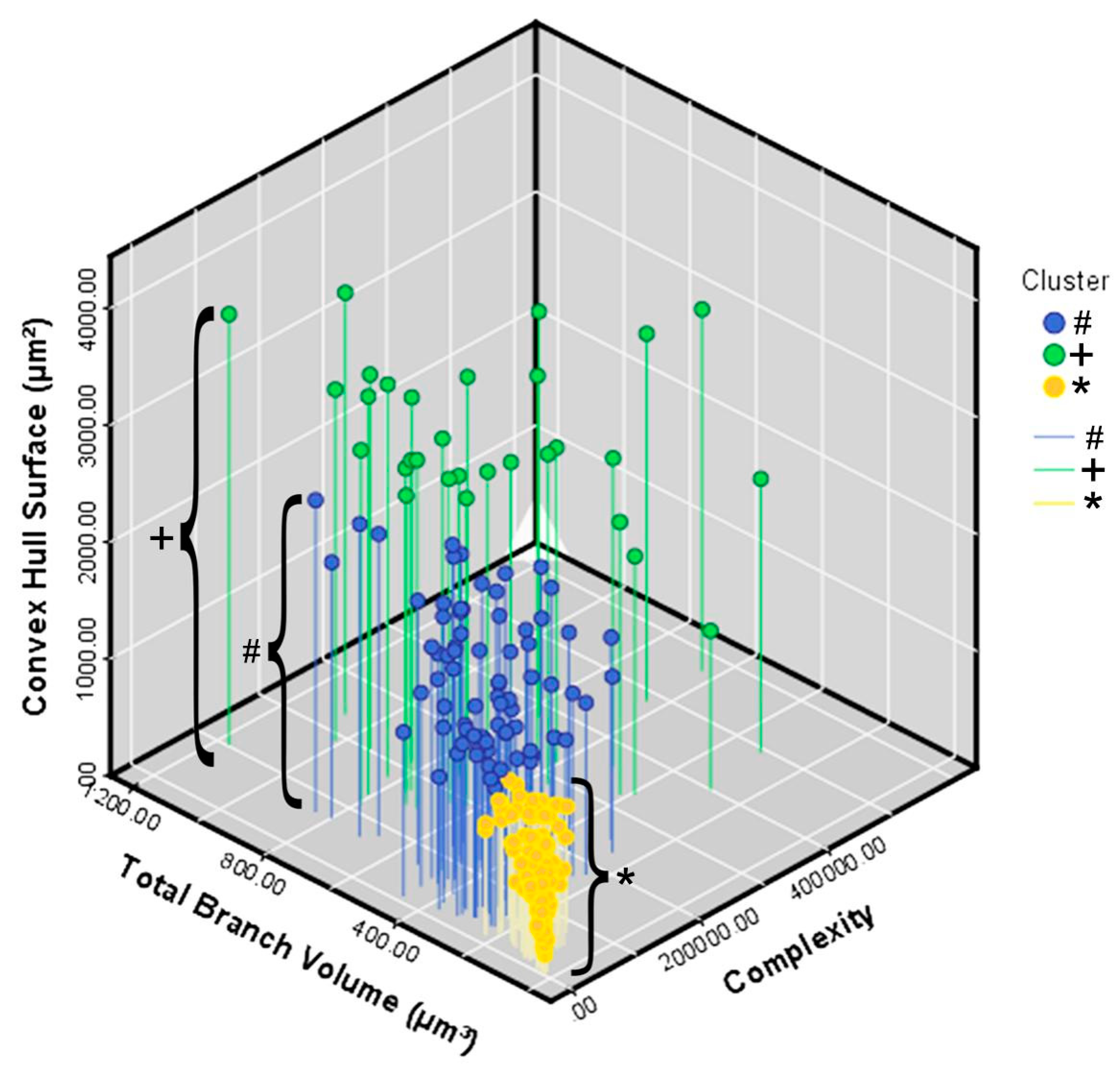
3. Results
Microglial Metamorphosis: From Surveillance to Reactivity to Virus Encephalitis
4. Discussion
4.1. Piry Virus Neuroinvasion and Microglial Response
4.2. Reactive Microglial Morphology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Data Sharing
Additional Information
References
- George, B.P.; Schneider, E.B.; Venkatesan, A. Encephalitis hospitalization rates and inpatient mortality in the United States, 2000–2010. PLoS ONE 2014, 9, e104169. [Google Scholar] [CrossRef] [Green Version]
- Barzon, L. Ongoing and emerging arbovirus threats in Europe. J. Clin. Virol. 2018, 107, 38–47. [Google Scholar] [CrossRef]
- Go, Y.Y.; Balasuriya, U.B.; Lee, C.K. Zoonotic encephalitides caused by arboviruses: Transmission and epidemiology of alphaviruses and flaviviruses. Clin. Exp. Vaccine Res. 2014, 3, 58–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhong, D.; Li, G. The role of microglia in viral encephalitis: A review. J. Neuroinflamm. 2019, 16, 76. [Google Scholar] [CrossRef] [PubMed]
- Klein, R.S.; Garber, C.; Funk, K.E.; Salimi, H.; Soung, A.; Kanmogne, M.; Manivasagam, S.; Agner, S.; Cain, M. Neuroinflammation During RNA Viral Infections. Annu. Rev. Immunol. 2019, 37, 73–95. [Google Scholar] [CrossRef]
- Clé, M.; Eldin, P.; Briant, L.; Lannuzel, A.; Simonin, Y.; Van de Perre, P.; Cabié, A.; Salinas, S. Neurocognitive impacts of arbovirus infections. J. Neuroinflamm. 2020, 17, 233. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, S.E.; Medeiros, M.; Porfirio, J.; Tavares, W.; Pessôa, L.; Grinberg, L.; Leite, R.E.P.; Ferretti-Rebustini, R.E.L.; Suemoto, C.K.; Filho, W.J.; et al. Similar Microglial Cell Densities across Brain Structures and Mammalian Species: Implications for Brain Tissue Function. J. Neurosci. 2020, 40, 4622–4643. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Sankowski, R.; Staszewski, O.; Prinz, M. Microglia Heterogeneity in the Single-Cell Era. Cell Rep. 2020, 30, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Geirsdottir, L.; David, E.; Keren-Shaul, H.; Weiner, A.; Bohlen, S.C.; Neuber, J.; Balic, A.; Giladi, A.; Sheban, F.; Dutertre, C.A.; et al. Cross-Species Single-Cell Analysis Reveals Divergence of the Primate Microglia Program. Cell 2019, 179, 1609–1622.e1616. [Google Scholar] [CrossRef] [Green Version]
- Perry, V.H. Microglia. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [Green Version]
- Wake, H.; Moorhouse, A.J.; Jinno, S.; Kohsaka, S.; Nabekura, J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J. Neurosci. 2009, 29, 3974–3980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinhard, L.; di Bartolomei, G.; Bolasco, G.; Machado, P.; Schieber, N.L.; Neniskyte, U.; Exiga, M.; Vadisiute, A.; Raggioli, A.; Schertel, A.; et al. Microglia remodel synapses by presynaptic trogocytosis and spine head filopodia induction. Nat. Commun. 2018, 9, 1228. [Google Scholar] [CrossRef] [Green Version]
- Ikegami, A.; Haruwaka, K.; Wake, H. Microglia: Lifelong modulator of neural circuits. Neuropathology 2019, 39, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illes, P.; Rubini, P.; Ulrich, H.; Zhao, Y.; Tang, Y. Regulation of Microglial Functions by Purinergic Mechanisms in the Healthy and Diseased CNS. Cells 2020, 9, 1108. [Google Scholar] [CrossRef]
- Madry, C.; Kyrargyri, V.; Arancibia-Cárcamo, I.L.; Jolivet, R.; Kohsaka, S.; Bryan, R.M.; Attwell, D. Microglial Ramification, Surveillance, and Interleukin-1β Release Are Regulated by the Two-Pore Domain K. Neuron 2018, 97, 299–312.e296. [Google Scholar] [CrossRef] [PubMed]
- Bernier, L.P.; Bohlen, C.J.; York, E.M.; Choi, H.B.; Kamyabi, A.; Dissing-Olesen, L.; Hefendehl, J.K.; Collins, H.Y.; Stevens, B.; Barres, B.A.; et al. Nanoscale Surveillance of the Brain by Microglia via cAMP-Regulated Filopodia. Cell Rep. 2019, 27, 2895–2908.e2894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanamsagar, R.; Alter, M.D.; Block, C.S.; Sullivan, H.; Bolton, J.L.; Bilbo, S.D. Generation of a microglial developmental index in mice and in humans reveals a sex difference in maturation and immune reactivity. Glia 2017, 65, 1504–1520. [Google Scholar] [CrossRef]
- Holtman, I.R.; Skola, D.; Glass, C.K. Transcriptional control of microglia phenotypes in health and disease. J. Clin. Investig. 2017, 127, 3220–3229. [Google Scholar] [CrossRef] [Green Version]
- Minten, C.; Terry, R.; Deffrasnes, C.; King, N.J.; Campbell, I.L. IFN regulatory factor 8 is a key constitutive determinant of the morphological and molecular properties of microglia in the CNS. PLoS ONE 2012, 7, e49851. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.M. Structural LTP: From synaptogenesis to regulated synapse enlargement and clustering. Curr. Opin. Neurobiol. 2020, 63, 189–197. [Google Scholar] [CrossRef]
- Davis, E.J.; Foster, T.D.; Thomas, W.E. Cellular forms and functions of brain microglia. Brain Res. Bull 1994, 34, 73–78. [Google Scholar] [CrossRef]
- de Sousa, A.A.; Reis, R.; Bento-Torres, J.; Trevia, N.; Lins, N.A.D.; Passos, A.; Santos, Z.; Diniz, J.A.P.; Vasconcelos, P.F.D.; Cunningham, C.; et al. Influence of Enriched Environment on Viral Encephalitis Outcomes: Behavioral and Neuropathological Changes in Albino Swiss Mice. PLoS ONE 2011, 6, e15597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa, A.A.; Dos Reis, R.R.; de Lima, C.M.; de Oliveira, M.A.; Fernandes, T.N.; Gomes, G.F.; Diniz, D.G.; Magalhães, N.M.; Diniz, C.G.; Sosthenes, M.C.; et al. Three-dimensional morphometric analysis of microglial changes in a mouse model of virus encephalitis: Age and environmental influences. Eur. J. Neurosci. 2015. [Google Scholar] [CrossRef]
- Glaser, J.R.; Glaser, E.M. Stereology, morphometry, and mapping: The whole is greater than the sum of its parts. J. Chem. Neuroanat. 2000, 20, 115–126. [Google Scholar] [CrossRef]
- West, M.J. Design-based stereological methods for counting neurons. Prog. Brain Res. 2002, 135, 43–51. [Google Scholar] [PubMed]
- Slomianka, L.; West, M. Estimators of the precision of stereological estimates: An example based on the CA1 pyramidal cell layer of rats. Neuroscience 2005, 136, 757–767. [Google Scholar] [CrossRef]
- Slomianka, L. Basic quantitative morphological methods applied to the central nervous system. J. Comp. Neurol. 2021, 529, 694–756. [Google Scholar] [CrossRef]
- Glaser, E.M.; Vanderloos, H. A Semi-Automatic Computer-Microscope for the Analysis of Neuronal Morphology. IEEE Trans. Biomed. Eng. 1965, 12, 22–31. [Google Scholar] [CrossRef]
- Wann, D.F.; Woolsey, T.A.; Dierker, M.L.; Cowan, W.M. An on-line digital-computer system for the semiautomatic analysis of Golgi-impregnated neurons. IEEE Trans. Biomed Eng. 1973, 20, 233–247. [Google Scholar] [CrossRef]
- Capowski, J.J. Computer-aided reconstruction of neuron trees from several serial sections. Comput. Biomed Res. 1977, 10, 617–629. [Google Scholar] [CrossRef]
- Zsuppán, F. A new approach to merging neuronal tree segments traced from serial sections. J. Neurosci. Methods 1984, 10, 199–204. [Google Scholar] [CrossRef]
- Parekh, R.; Ascoli, G.A. Neuronal morphology goes digital: A research hub for cellular and system neuroscience. Neuron 2013, 77, 1017–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackman, A.V.; Grabuschnig, S.; Legenstein, R.; Sjöström, P.J. A comparison of manual neuronal reconstruction from biocytin histology or 2-photon imaging: Morphometry and computer modeling. Front. Neuroanat. 2014, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, L.; Renehan, W.E. The use of cluster analysis for cell typing. Brain Res. Brain Res. Protoc. 1997, 1, 100–108. [Google Scholar] [CrossRef]
- Yamada, J.; Jinno, S. Novel objective classification of reactive microglia following hypoglossal axotomy using hierarchical cluster analysis. J. Comp. Neurol. 2013, 521, 1184–1201. [Google Scholar] [CrossRef]
- Fernández-Arjona, M.D.M.; Grondona, J.M.; Fernández-Llebrez, P.; López-Ávalos, M.D. Microglial Morphometric Parameters Correlate with the Expression Level of IL-1β, and Allow Identifying Different Activated Morphotypes. Front. Cell Neurosci. 2019, 13, 472. [Google Scholar] [CrossRef] [Green Version]
- Sheth, C.; Ombach, H.; Olson, P.; Renshaw, P.F.; Kanekar, S. Increased Anxiety and Anhedonia in Female Rats Following Exposure to Altitude. High Alt. Med. Biol. 2018, 19, 81–90. [Google Scholar] [CrossRef]
- Xiao, S.Y.; Guzman, H.; Zhang, H.; Travassos da Rosa, A.P.; Tesh, R.B. West Nile virus infection in the golden hamster (Mesocricetus auratus): A model for West Nile encephalitis. Emerg. Infect Dis. 2001, 7, 714–721. [Google Scholar] [CrossRef]
- Reed, H.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Hawk, C.; Leary, S.; Morris, T. Formulary for Laboratory Animals, 3rd ed.; Blackwell Publishing: Iowa, IA, USA, 2005. [Google Scholar]
- Ohsawa, K.; Imai, Y.; Sasaki, Y.; Kohsaka, S. Microglia/macrophage-specific protein Iba1 binds to fimbrin and enhances its actin-bundling activity. J. Neurochem. 2004, 88, 844–856. [Google Scholar] [CrossRef]
- Ohsawa, K.; Kohsaka, S. Dynamic motility of microglia: Purinergic modulation of microglial movement in the normal and pathological brain. Glia 2011. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K. The Mouse Brain in Stereotaxic Coordinates; Academic Press: London, UK, 2001. [Google Scholar]
- Carlo, C.N.; Stevens, C.F. Analysis of differential shrinkage in frozen brain sections and its implications for the use of guard zones in stereology. J. Comp. Neurol. 2011, 519, 2803–2810. [Google Scholar] [CrossRef]
- Ward, J. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Costa, L.A.F.; Velte, T.J. Automatic characterization and classification of ganglion cells from the salamander retina. J. Comp. Neurol. 1999, 404, 33–51. [Google Scholar] [CrossRef]
- Kolb, H.; Fernandez, E.; Schouten, J.; Ahnelt, P.; Linberg, K.A.; Fisher, S.K. Are there three types of horizontal cell in the human retina? J. Comp. Neurol. 1994, 343, 370–386. [Google Scholar] [CrossRef] [PubMed]
- McGavern, D.B.; Kang, S.S. Illuminating viral infections in the nervous system. Nat. Rev. Immunol. 2011, 11, 318–329. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.S.; Garber, C.; Howard, N. Infectious immunity in the central nervous system and brain function. Nat. Immunol. 2017, 18, 132–141. [Google Scholar] [CrossRef]
- Payus, A.O.; Liew Sat Lin, C.; Mohd Noh, M.; Jeffree, M.S.; Ali, R.A. SARS-CoV-2 infection of the nervous system: A review of the literature on neurological involvement in novel coronavirus disease-(COVID-19). Bosn. J. Basic Med. Sci. 2020, 20, 283–292. [Google Scholar] [CrossRef]
- Weaver, S.C. Evolutionary influences in arboviral disease. Curr. Top Microbiol. Immunol. 2006, 299, 285–314. [Google Scholar]
- Marriott, A.C. Complete genome sequences of Chandipura and Isfahan vesiculoviruses. Arch. Virol. 2005, 150, 671–680. [Google Scholar] [CrossRef]
- Gomes-Leal, W.; Martins, L.C.; Diniz, J.A.P.; Dos Santos, Z.A.; Borges, J.A.; Macedo, C.A.C.; Medeiros, A.C.; De Paula, L.S.; Guimaraes, J.S.; Freire, M.A.M.; et al. Neurotropism and neuropathological effects of selected rhabdoviruses on intranasally-infected newborn mice. Acta Trop. 2006, 97, 126–139. [Google Scholar] [CrossRef]
- Wilks, C.R.; House, J.A. Susceptibility of various animals to the vesiculovirus Piry. J. Hyg. 1984, 93, 147–155. [Google Scholar] [CrossRef] [Green Version]
- da Cruz, E.R. Ultrastructural lesions of the brain in albino mice experimentally inoculated with Piry virus (BeAn 24232). Rev. Inst. Med. Trop. Sao Paulo 1981, 23, 194–203. [Google Scholar] [PubMed]
- Berge, T. International Catalogue of Arboviruses, 2nd ed.; Department of Health, Education and Welfare, Centers for Disease Control: Atlanta, GA, USA, 1975. [Google Scholar]
- Vasconcelos, P.F.; Da Rosa, J.F.; Da Rosa, A.P.; Degallier, N.; Pinheiro, F.P.; Sa Filho, G.C. Epidemiology of encephalitis caused by arbovirus in the Brazilian Amazonia. Rev. Inst. Med. Trop. Sao Paulo 1991, 33, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Esiri, M.M.; Tomlinson, A.H. Herpes simplex encephalitis. Immunohistological demonstration of spread of virus via olfactory and trigeminal pathways after infection of facial skin in mice. J. Neurol. Sci. 1984, 64, 213–217. [Google Scholar] [CrossRef]
- Barnett, E.M.; Perlman, S. The olfactory nerve and not the trigeminal nerve is the major site of CNS entry for mouse hepatitis virus, strain JHM. Virology 1993, 194, 185–191. [Google Scholar] [CrossRef]
- Babic, N.; Mettenleiter, T.C.; Ugolini, G.; Flamand, A.; Coulon, P. Propagation of pseudorabies virus in the nervous system of the mouse after intranasal inoculation. Virology 1994, 204, 616–625. [Google Scholar] [CrossRef]
- Charles, P.C.; Walters, E.; Margolis, F.; Johnston, R.E. Mechanism of neuroinvasion of Venezuelan equine encephalitis virus in the mouse. Virology 1995, 208, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Lafay, F.; Coulon, P.; Astic, L.; Saucier, D.; Riche, D.; Holley, A.; Flamand, A. Spread of the CVS strain of rabies virus and of the avirulent mutant AvO1 along the olfactory pathways of the mouse after intranasal inoculation. Virology 1991, 183, 320–330. [Google Scholar] [CrossRef]
- Menassa, D.A.; Gomez-Nicola, D. Microglial Dynamics During Human Brain Development. Front. Immunol. 2018, 9, 1014. [Google Scholar] [CrossRef] [Green Version]
- Wierzba-Bobrowicz, T.; Kosno-Kruszewska, E.; Gwiazda, E.; Lechowicz, W. The comparison of microglia maturation in different structures of the human nervous system. Folia Neuropathol. 1998, 36, 152–160. [Google Scholar]
- Karperien, A.; Ahammer, H.; Jelinek, H.F. Quantitating the subtleties of microglial morphology with fractal analysis. Front. Cell Neurosci. 2013, 7. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Arjona, M.D.M.; Grondona, J.M.; Granados-Durán, P.; Fernández-Llebrez, P.; López-Ávalos, M.D. Microglia Morphological Categorization in a Rat Model of Neuroinflammation by Hierarchical Cluster and Principal Components Analysis. Front. Cell Neurosci. 2017, 11, 235. [Google Scholar] [CrossRef] [Green Version]
- Pillai, A.G.; de Jong, D.; Kanatsou, S.; Krugers, H.; Knapman, A.; Heinzmann, J.M.; Holsboer, F.; Landgraf, R.; Joëls, M.; Touma, C. Dendritic morphology of hippocampal and amygdalar neurons in adolescent mice is resilient to genetic differences in stress reactivity. PLoS ONE 2012, 7, e38971. [Google Scholar] [CrossRef] [Green Version]
- Hinwood, M.; Morandini, J.; Day, T.A.; Walker, F.R. Evidence that microglia mediate the neurobiological effects of chronic psychological stress on the medial prefrontal cortex. Cereb. Cortex. 2012, 22, 1442–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinwood, M.; Tynan, R.J.; Charnley, J.L.; Beynon, S.B.; Day, T.A.; Walker, F.R. Chronic stress induced remodeling of the prefrontal cortex: Structural re-organization of microglia and the inhibitory effect of minocycline. Cereb. Cortex. 2013, 23, 1784–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couch, Y.; Anthony, D.C.; Dolgov, O.; Revischin, A.; Festoff, B.; Santos, A.I.; Steinbusch, H.W.; Strekalova, T. Microglial activation, increased TNF and SERT expression in the prefrontal cortex define stress-altered behaviour in mice susceptible to anhedonia. Brain Behav. Immun. 2013, 29, 136–146. [Google Scholar] [CrossRef]
- Gomes, G.F.; Peixoto, R.D.D.F.; Maciel, B.G.; Santos, K.F.D.; Bayma, L.R.; Feitoza Neto, P.A.; Fernandes, T.N.; de Abreu, C.C.; Casseb, S.M.M.; de Lima, C.M.; et al. Differential Microglial Morphological Response, TNFα, and Viral Load in Sedentary-like and Active Murine Models After Systemic Non-neurotropic Dengue Virus Infection. J. Histochem. Cytochem. 2019, 67, 419–439. [Google Scholar] [CrossRef] [PubMed]
- Diniz, D.G.; Silva, G.O.; Naves, T.B.; Fernandes, T.N.; Araujo, S.C.; Diniz, J.A.P.; de Farias, L.H.S.; Sosthenes, M.C.K.; Diniz, C.G.; Anthony, D.C.; et al. Hierarchical Cluster Analysis of Three-Dimensional Reconstructions of Unbiased Sampled Microglia Shows not Continuous Morphological Changes from Stage 1 to 2 after Multiple Dengue Infections in Callithrix penicillata. Front. Neuroanat. 2016, 10. [Google Scholar] [CrossRef]
- Vasconcelos, B.C.B.; Vieira, J.A.; Silva, G.O.; Fernandes, T.N.; Rocha, L.C.; Viana, A.P.; Serique, C.D.S.; Santos, C.; Bringel, R.A.R.; Teixeira, F.; et al. Antibody-enhanced dengue disease generates a marked CNS inflammatory response in the black-tufted marmoset Callithrix penicillata. Neuropathology 2016, 36, 3–16. [Google Scholar] [CrossRef]
- Ayoub, A.E.; Salm, A.K. Increased morphological diversity of microglia in the activated hypothalamic supraoptic nucleus. J. Neurosci. 2003, 23, 7759–7766. [Google Scholar] [CrossRef]
- Kloss, C.U.; Bohatschek, M.; Kreutzberg, G.W.; Raivich, G. Effect of lipopolysaccharide on the morphology and integrin immunoreactivity of ramified microglia in the mouse brain and in cell culture. Exp. Neurol. 2001, 168, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Madeddu, S.; Woods, T.A.; Mukherjee, P.; Sturdevant, D.; Butchi, N.B.; Peterson, K.E. Identification of Glial Activation Markers by Comparison of Transcriptome Changes between Astrocytes and Microglia following Innate Immune Stimulation. PLoS ONE 2015, 10, e0127336. [Google Scholar] [CrossRef] [PubMed]
- Getts, D.R.; Terry, R.L.; Getts, M.T.; Müller, M.; Rana, S.; Shrestha, B.; Radford, J.; Van Rooijen, N.; Campbell, I.L.; King, N.J. Ly6c+ “inflammatory monocytes” are microglial precursors recruited in a pathogenic manner in West Nile virus encephalitis. J. Exp. Med. 2008, 205, 2319–2337. [Google Scholar] [CrossRef] [Green Version]
- D’Agostino, P.M.; Kwak, C.; Vecchiarelli, H.A.; Toth, J.G.; Miller, J.M.; Masheeb, Z.; McEwen, B.S.; Bulloch, K. Viral-induced encephalitis initiates distinct and functional CD103+ CD11b+ brain dendritic cell populations within the olfactory bulb. Proc. Natl. Acad. Sci. USA 2012, 109, 6175–6180. [Google Scholar] [CrossRef] [Green Version]
- Steel, C.D.; Hahto, S.M.; Ciavarra, R.P. Peripheral dendritic cells are essential for both the innate and adaptive antiviral immune responses in the central nervous system. Virology 2009, 387, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Chhatbar, C.; Detje, C.N.; Grabski, E.; Borst, K.; Spanier, J.; Ghita, L.; Elliott, D.A.; Jordão, M.J.C.; Mueller, N.; Sutton, J.; et al. Type I Interferon Receptor Signaling of Neurons and Astrocytes Regulates Microglia Activation during Viral Encephalitis. Cell Rep. 2018, 25, 118–129.e114. [Google Scholar] [CrossRef] [Green Version]
- Hickman, S.E.; El Khoury, J. Analysis of the Microglial Sensome. Methods Mol. Biol. 2019, 2034, 305–323. [Google Scholar] [CrossRef] [PubMed]
- Garaschuk, O.; Verkhratsky, A. Physiology of Microglia. Methods Mol. Biol. 2019, 2034, 27–40. [Google Scholar] [CrossRef]
- Sierra, A.; Paolicelli, R.C.; Kettenmann, H. Cien Años de Microglía: Milestones in a Century of Microglial Research. Trends Neurosci. 2019, 42, 778–792. [Google Scholar] [CrossRef] [Green Version]
- Hanisch, U.K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. A polarizing question: Do M1 and M2 microglia exist? Nat. Neurosci. 2016, 19, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Perry, V.H. Microglial physiology: Unique stimuli, specialized responses. Annu. Rev. Immunol. 2009, 27, 119–145. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Nicola, D.; Perry, V.H. Microglial dynamics and role in the healthy and diseased brain: A paradigm of functional plasticity. Neuroscientist 2015, 21, 169–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karperien, A.L.; Jelinek, H.F. Fractal, multifractal, and lacunarity analysis of microglia in tissue engineering. Front. Bioeng. Biotechnol. 2015, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubbelaar, M.L.; Kracht, L.; Eggen, B.J.L.; Boddeke, E.W.G.M. The Kaleidoscope of Microglial Phenotypes. Front. Immunol. 2018, 9, 1753. [Google Scholar] [CrossRef]
- Savage, J.C.; Carrier, M.; Tremblay, M. Morphology of Microglia Across Contexts of Health and Disease. Methods Mol. Biol. 2019, 2034, 13–26. [Google Scholar] [CrossRef]
- Carvalho-Paulo, D.; Bento Torres Neto, J.; de Oliveira TC, G.; de Sousa, A.A.; dos Reis, R.R.; dos Santos, Z.A.; de lima, C.M.; de Oliveira, M.A.; Said, N.M.; Freitas, S.F.; et al. Microglial morphology across distantly related species: Phylogenetic, environmental and age influences on microglia reactive and surveillance states. Front. Immunol. 2021, 12, 683026. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Branched Structure Analysis | |
|---|---|
| Segment | Any portion of microglia branched structure with endings that are either nodes or terminations with no intermediate nodes |
| Segments/mm | Number of segments/total length of the segments, expressed in millimeters |
| No of trees | Number of trees in the microglia |
| Total No of segments | Total number of segments in the tree |
| Single branch length | Total length of the line segments used to trace the branch of interest |
| Mean branch length (µm) | Mean = [Total length]/[Number of branches] |
| Total branch length (µm) | Total length for all branches in the tree |
| Tortuosity | Tortuosity = [Actual length of the segment] /[Distance between the endpoints of the segment]; smallest value is 1, which represents a straight segment; tortuosity allows segments of different lengths to be compared in terms of the complexity of the paths they take |
| Mean branch surface area (µm2) | Computed by modeling each branch as a frustum (truncated right circular cone) divided by the number of branches |
| Total tree surface area (µm2) | Two-dimensional (2D) surface area of a microglia arbor computed based on the area defined by the endpoints of all trees |
| Branch volume (µm3) | Computed by modeling each piece of each branch as a frustum |
| Total branch volume | Total volume for all branches in the tree |
| Base diameter of primary branch (µm) | Diameter at the start of the first segment |
| Planar angle | Computed based on the endpoints of the segments; references the change in direction of a segment relative to the previous segment |
| Fractal dimension k-dim | The “k-dim” of the fractal analysis, describing how the structure of interest fills space; significant statistical differences in k-dim suggest morphological dissimilarities |
| Convex hull: perimeter (µm), area (µm2), 2D surface area (µm2), 3D or volume (µm3) | Convex hull measures the size of the branching field by interpreting a branched structure as a solid object controlling a given amount of physical space; the amount of physical space is defined in terms of convex-hull volume, surface area, area, and/or perimeter. |
| Vertex analysis | Describes the overall structure of a branched object based on topological and metrical properties. Root (or origin) point: For neurons, microglia or astrocytes, the origin is the point at which the structure is attached to the soma. Main types of vertices: Vd (bifurcation) or Vt (trifurcation), nodal (or branching) points. Vp: Terminal (or pendant) vertices. Va: primary vertices connecting 2 pendant vertices; Vb: secondary vertices connecting 1 pendant vertex (Vp) to 1 bifurcation (Vd) or 1 trifurcation (Vt); Vc: tertiary vertices connecting either 2 bifurcations (Vd), 2 trifurcations (Vt), or 1 bifurcation (Vd) and 1 trifurcation (Vt). In the present report, we measured the number of vertices Va, Vb, and Vc. |
| Complexity | Complexity = [Sum of the terminal orders + Number of terminals] × [Total branch length/Number of primary branches] |
| Descriptive Results | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| N | Mean | Std. Deviation | Std. Error | 95% Confidence Interval for Mean | Minimum | Maximum | |||
| Lower Bound | Upper Bound | ||||||||
| Tortuosity | 1 | 75 | 1.33 | 0.12 | 0.01 | 1.30 | 1.36 | 1.09 | 1.56 |
| 2 | 30 | 1.35 | 0.09 | 0.02 | 1.31 | 1.38 | 1.18 | 1.59 | |
| 3 | 106 | 1.20 | 0.09 | 0.01 | 1.18 | 1.21 | 1.08 | 1.66 | |
| Total | 211 | 1.27 | 0.12 | 0.01 | 1.25 | 1.28 | 1.08 | 1.66 | |
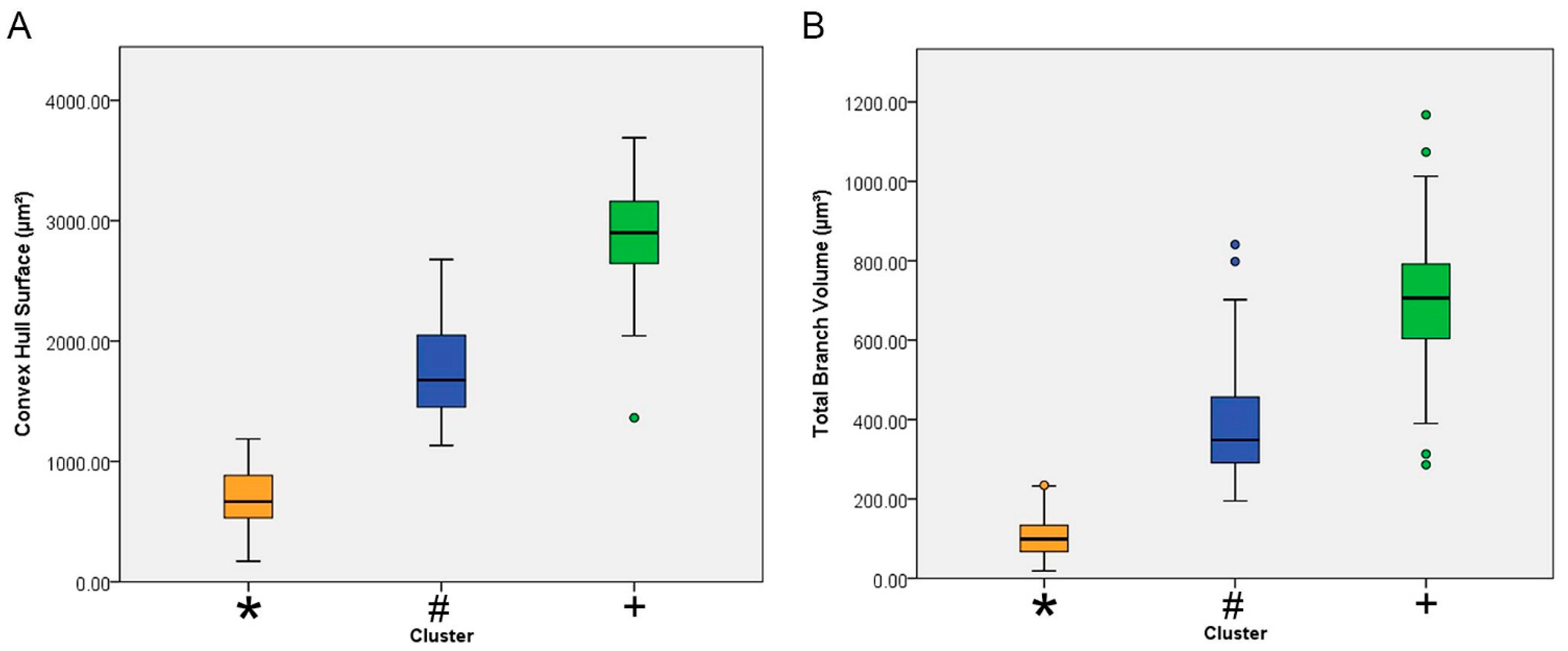
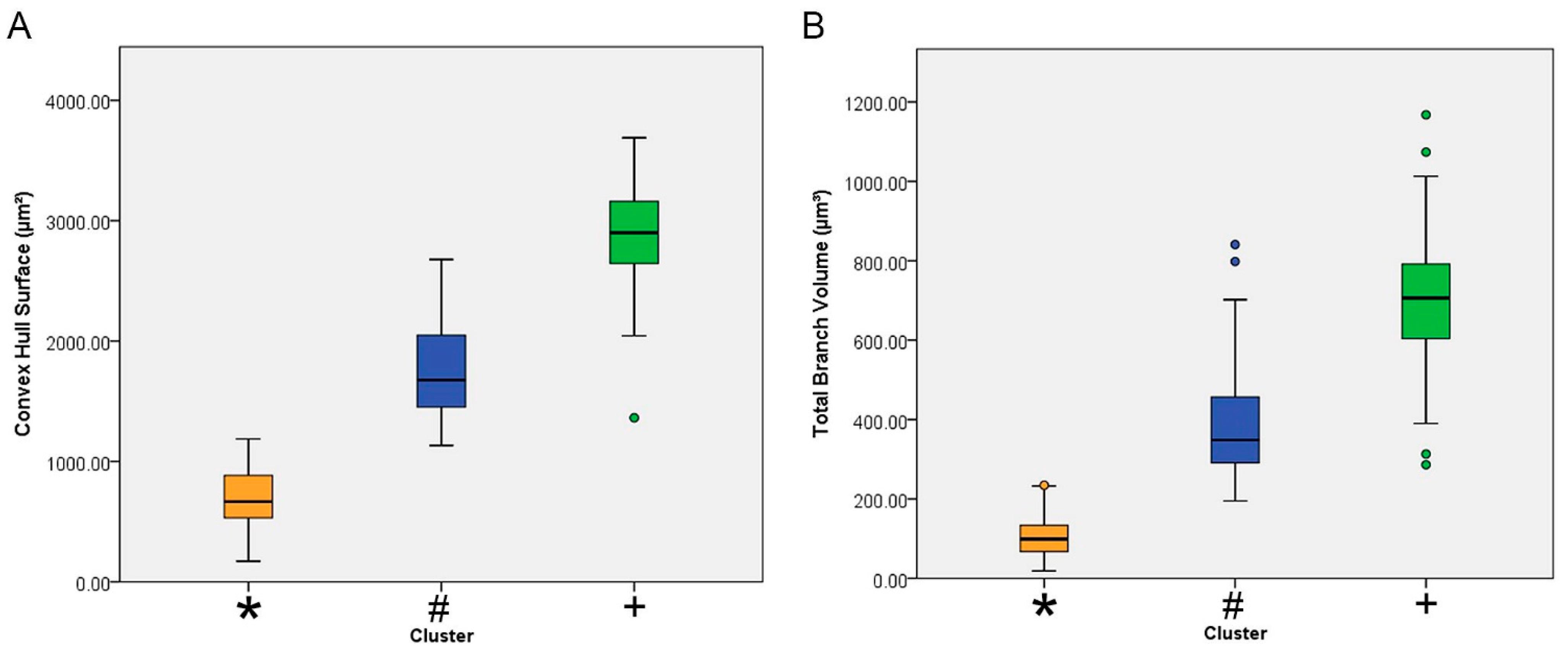
| Total branch volume (µm3) | 1 | 75 | 381.99 | 131.21 | 15.15 | 351.80 | 412.17 | 194.82 | 840.86 |
| 2 | 30 | 696.56 | 202.32 | 36.94 | 621.01 | 772.11 | 286.29 | 1167.60 | |
| 3 | 106 | 104.39 | 52.92 | 5.14 | 94.20 | 114.58 | 18.69 | 234.52 | |
| Total | 211 | 287.26 | 239.03 | 16.46 | 254.82 | 319.70 | 18.69 | 1167.60 | |
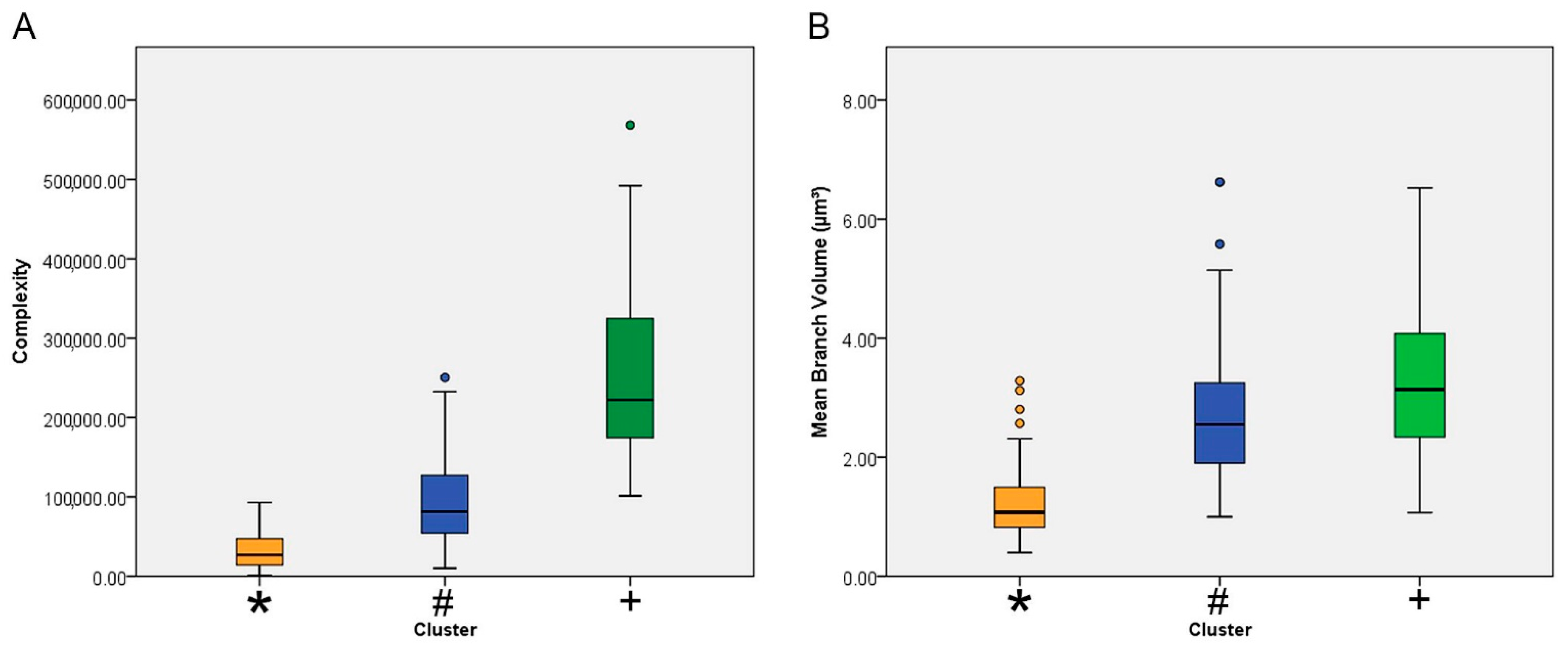
| Mean branch volume (µm3) | 1 | 75 | 2.79 | 1.13 | 0.13 | 2.53 | 3.05 | 1.00 | 6.62 |
| 2 | 30 | 3.34 | 1.36 | 0.25 | 2.83 | 3.85 | 1.07 | 6.52 | |
| 3 | 106 | 1.22 | 0.55 | 0.05 | 1.12 | 1.33 | 0.40 | 3.28 | |
| Total | 211 | 2.08 | 1.28 | 0.09 | 1.91 | 2.25 | 0.40 | 6.62 | |
| Convex hull surface (µm2) | 1 | 75 | 1768.35 | 392.52 | 45.32 | 1678.04 | 1858.66 | 1133.26 | 2677.56 |
| 2 | 30 | 2882.78 | 473.73 | 86.49 | 2705.89 | 3059.67 | 1362.55 | 3689.12 | |
| 3 | 106 | 689.23 | 247.31 | 24.02 | 641.60 | 736.86 | 170.98 | 1186.62 | |
| Total | 211 | 1384.68 | 856.26 | 58.95 | 1268.48 | 1500.89 | 170.98 | 3689.12 | |
| Complexity | 1 | 75 | 945,72.82 | 53,188.84 | 6141.72 | 82,335.18 | 106,810.46 | 10,038.40 | 250,383.00 |
| 2 | 30 | 261,064.33 | 117,350.51 | 21,425.17 | 217,244.93 | 304,883.73 | 101,354.00 | 568,527.00 | |
| 3 | 106 | 31,720.23 | 21,318.02 | 2070.59 | 27,614.64 | 35,825.83 | 1451.06 | 92,769.60 | |
| Total | 211 | 86,669.37 | 94,958.17 | 6537.19 | 73,782.44 | 99,556.29 | 1451.06 | 568,527.00 | |
| One-Way ANOVA | ||||||
|---|---|---|---|---|---|---|
| Sum of Squares | df | Mean Square | F | Sig. | ||
| Tortuosity | Between groups | 1.01 | 2.00 | 0.50 | 51.64 | 0.00 |
| Within groups | 2.03 | 208.00 | 0.01 | |||
| Total | 3.03 | 210.00 | ||||
| Total branch volume (µm3) | Between groups | 9,243,466.58 | 2.00 | 4,621,733.29 | 348.92 | 0.00 |
| Within groups | 2,755,100.53 | 208.00 | 13,245.68 | |||
| Total | 11,998,567.12 | 210.00 | ||||
| Mean branch volume (µm3) | Between groups | 163.21 | 2.00 | 81.61 | 94.20 | 0.00 |
| Within groups | 180.20 | 208.00 | 0.87 | |||
| Total | 343.41 | 210.00 | ||||
| Convex hull surface (µm2) | Between groups | 129,635,754.77 | 2.00 | 64,817,877.38 | 554.10 | 0.00 |
| Within groups | 24,331,445.08 | 208.00 | 116,978.10 | |||
| Total | 153,967,199.85 | 210.00 | ||||
| Complexity | Between groups | 1,237,150,125,189.31 | 2.00 | 618,575,062,594.66 | 196.00 | 0.00 |
| Within groups | 656,431,092,622.02 | 208.00 | 3,155,918,714.53 | |||
| Total | 1,893,581,217,811.34 | 210.00 | ||||
| Multiple Comparisons | ||||||||
|---|---|---|---|---|---|---|---|---|
| Dependent Variable | Between-Groups Comparisons | Mean Difference (I-J) | Std. Error | Sig. | 95% Confidence Interval | |||
| Lower Bound | Upper Bound | |||||||
| Tortuosity | Tukey HSD | 1 vs. | 2 | −0.017 | 0.021 | 0.710 | −0.067 | 0.034 |
| 3 | 0.13291 * | 0.015 | 0.000 | 0.098 | 0.168 | |||
| 2 | 1 | 0.017 | 0.021 | 0.710 | −0.034 | 0.067 | ||
| 3 | 0.14976 * | 0.020 | 0.000 | 0.102 | 0.198 | |||
| 3 | 1 | −0.13291 * | 0.015 | 0.000 | −0.168 | −0.098 | ||
| 2 | −0.14976 * | 0.020 | 0.000 | −0.198 | −0.102 | |||
| Total branch volume (µm3) | Tukey HSD | 1 vs. | 2 | −314.57115 * | 24.862 | 0.000 | −373.261 | −255.881 |
| 3 | 277.59531 * | 17.366 | 0.000 | 236.602 | 318.589 | |||
| 2 | 1 | 314.57115 * | 24.862 | 0.000 | 255.881 | 373.261 | ||
| 3 | 592.16647 * | 23.801 | 0.000 | 535.982 | 648.351 | |||
| 3 | 1 | −277.59531 * | 17.366 | 0.000 | −318.589 | −236.602 | ||
| 2 | −592.16647 * | 23.801 | 0.000 | −648.351 | −535.982 | |||
| Mean branch volume (µm3) | Tukey HSD | 1 vs. | 2 | −0.55013 * | 0.201 | 0.018 | −1.025 | −0.075 |
| 3 | 1.56653 * | 0.140 | 0.000 | 1.235 | 1.898 | |||
| 2 | 1 | 0.55013 * | 0.201 | 0.018 | 0.075 | 1.025 | ||
| 3 | 2.11666 * | 0.192 | 0.000 | 1.662 | 2.571 | |||
| 3 | 1 | −1.56653 * | 0.140 | 0.000 | −1.898 | −1.235 | ||
| 2 | −2.11666 * | 0.192 | 0.000 | −2.571 | −1.662 | |||
| Convex hull surface (µm2) | Tukey HSD | 1 vs. | 2 | −1114.43247 * | 73.885 | 0.000 | −1288.846 | −940.019 |
| 3 | 1079.11491 * | 51.607 | 0.000 | 957.291 | 1200.939 | |||
| 2 | 1 | 1114.43247 * | 73.885 | 0.000 | 940.019 | 1288.846 | ||
| 3 | 2193.54738 * | 70.731 | 0.000 | 2026.579 | 2360.515 | |||
| 3 | 1 | −1079.11491 * | 51.607 | 0.000 | −1200.939 | −957.291 | ||
| 2 | −2193.54738 * | 70.731 | 0.000 | −2360.515 | −2026.579 | |||
| Complexity | Tukey HSD | 1 vs. | 2 | −166,491.51467 * | 12,135.741 | 0.000 | −195,139.323 | −137,843.707 |
| 3 | 62,852.58386 * | 8476.540 | 0.000 | 42,842.739 | 82,862.429 | |||
| 2 | 1 | 166,491.51467 * | 12,135.741 | 0.000 | 137,843.707 | 195,139.323 | ||
| 3 | 229,344.09852 * | 11,617.664 | 0.000 | 201,919.271 | 256,768.926 | |||
| 3 | 1 | −62,852.58386 * | 8476.540 | 0.000 | −82,862.429 | −42,842.739 | ||
| 2 | −229,344.09852 * | 11,617.664 | 0.000 | −256,768.926 | −201,919.271 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva Creão, L.S.; Neto, J.B.T.; de Lima, C.M.; dos Reis, R.R.; de Sousa, A.A.; dos Santos, Z.A.; Diniz, J.A.P.; Diniz, D.G.; Diniz, C.W.P. Microglial Metamorphosis in Three Dimensions in Virus Limbic Encephalitis: An Unbiased Pictorial Representation Based on a Stereological Sampling Approach of Surveillant and Reactive Microglia. Brain Sci. 2021, 11, 1009. https://doi.org/10.3390/brainsci11081009
da Silva Creão LS, Neto JBT, de Lima CM, dos Reis RR, de Sousa AA, dos Santos ZA, Diniz JAP, Diniz DG, Diniz CWP. Microglial Metamorphosis in Three Dimensions in Virus Limbic Encephalitis: An Unbiased Pictorial Representation Based on a Stereological Sampling Approach of Surveillant and Reactive Microglia. Brain Sciences. 2021; 11(8):1009. https://doi.org/10.3390/brainsci11081009
Chicago/Turabian Styleda Silva Creão, Leonardo Sávio, João Bento Torres Neto, Camila Mendes de Lima, Renata Rodrigues dos Reis, Aline Andrade de Sousa, Zaire Alves dos Santos, José Antonio Picanço Diniz, Daniel Guerreiro Diniz, and Cristovam Wanderley Picanço Diniz. 2021. "Microglial Metamorphosis in Three Dimensions in Virus Limbic Encephalitis: An Unbiased Pictorial Representation Based on a Stereological Sampling Approach of Surveillant and Reactive Microglia" Brain Sciences 11, no. 8: 1009. https://doi.org/10.3390/brainsci11081009
APA Styleda Silva Creão, L. S., Neto, J. B. T., de Lima, C. M., dos Reis, R. R., de Sousa, A. A., dos Santos, Z. A., Diniz, J. A. P., Diniz, D. G., & Diniz, C. W. P. (2021). Microglial Metamorphosis in Three Dimensions in Virus Limbic Encephalitis: An Unbiased Pictorial Representation Based on a Stereological Sampling Approach of Surveillant and Reactive Microglia. Brain Sciences, 11(8), 1009. https://doi.org/10.3390/brainsci11081009