

A Characterization of Brain Area Activation in Orienteers with Different Map-Recognition Memory Ability Task Levels—Based on fNIRS Evidence

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
2.3. Experimental Materials
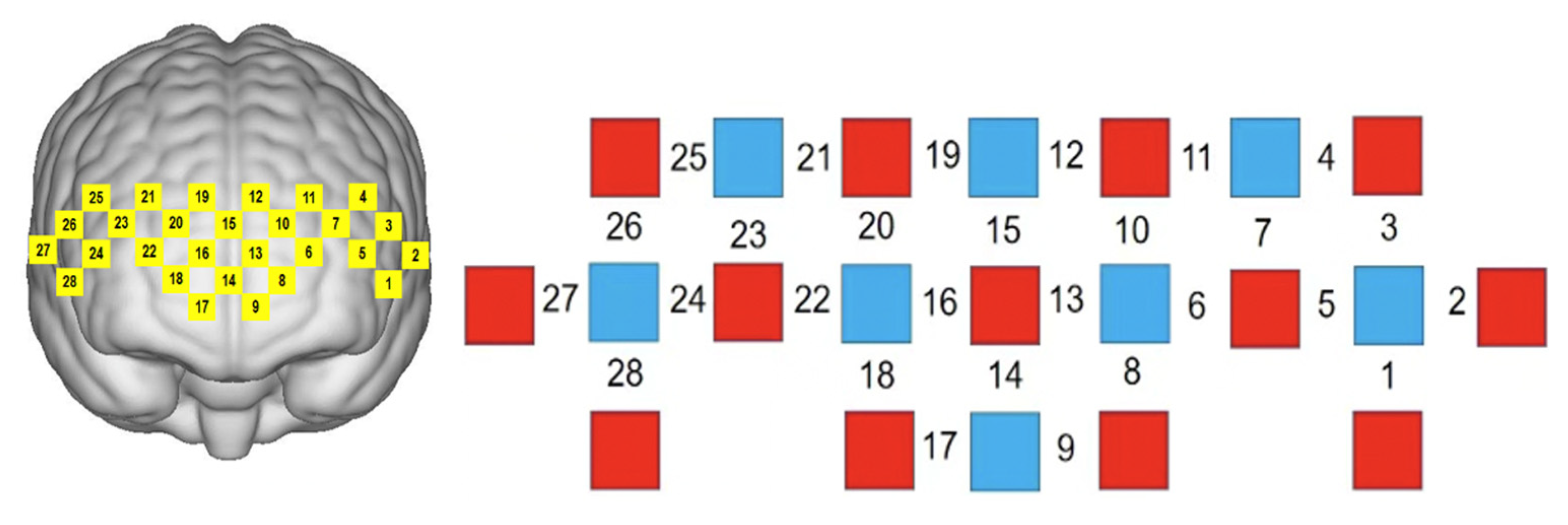
2.4. fNIRS Test Protocol
2.5. Experiment Procedures
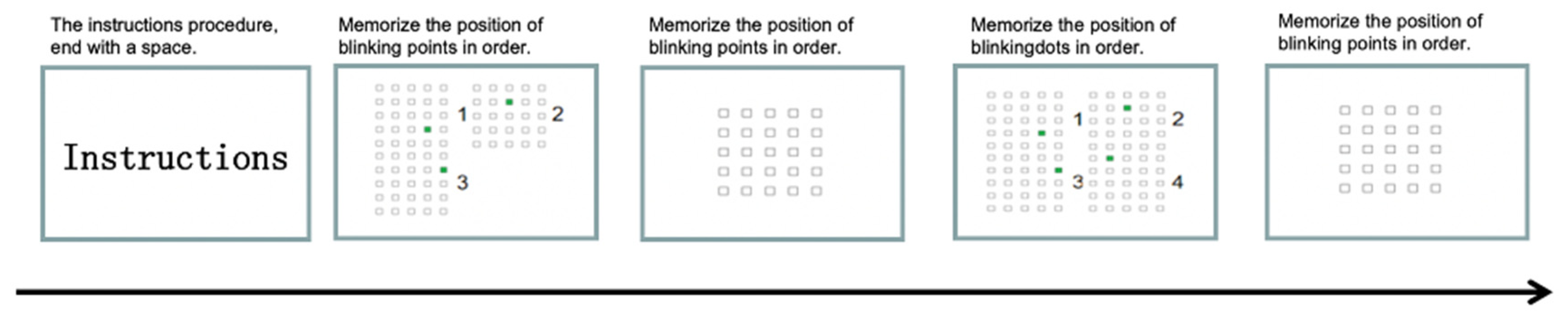
2.5.1. Non-Orienteering Scenario Experiment Flow
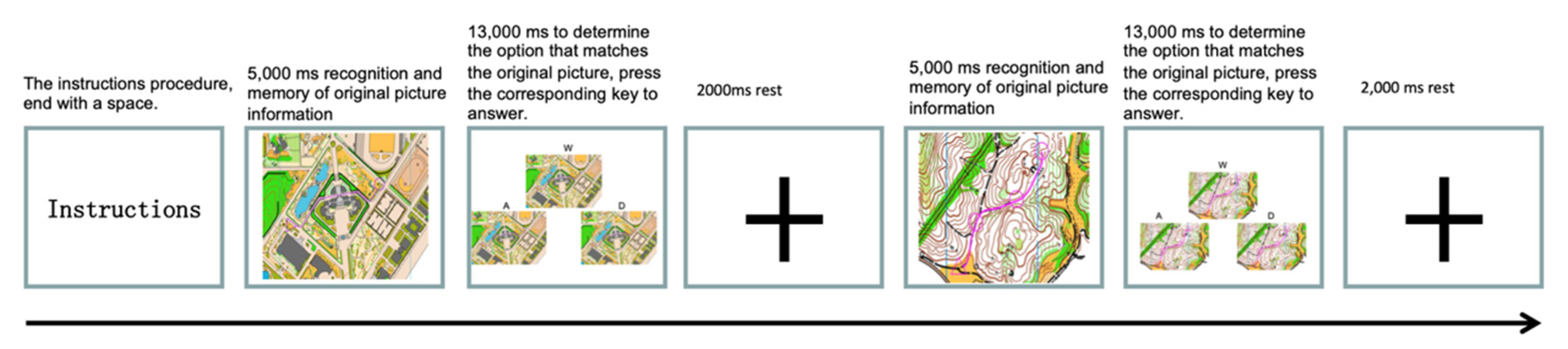
2.5.2. Orienteering Scenario Experiment Flow
2.6. Data Processing and Statistics
2.6.1. Behavioral Data Processing
2.6.2. fNIRS Data Processing
3. Results
3.1. Results of Spatial Memory Ability for Non-Orienteering Scenes
3.2. Results of Spatial Memory Ability for Orienteering Scenes
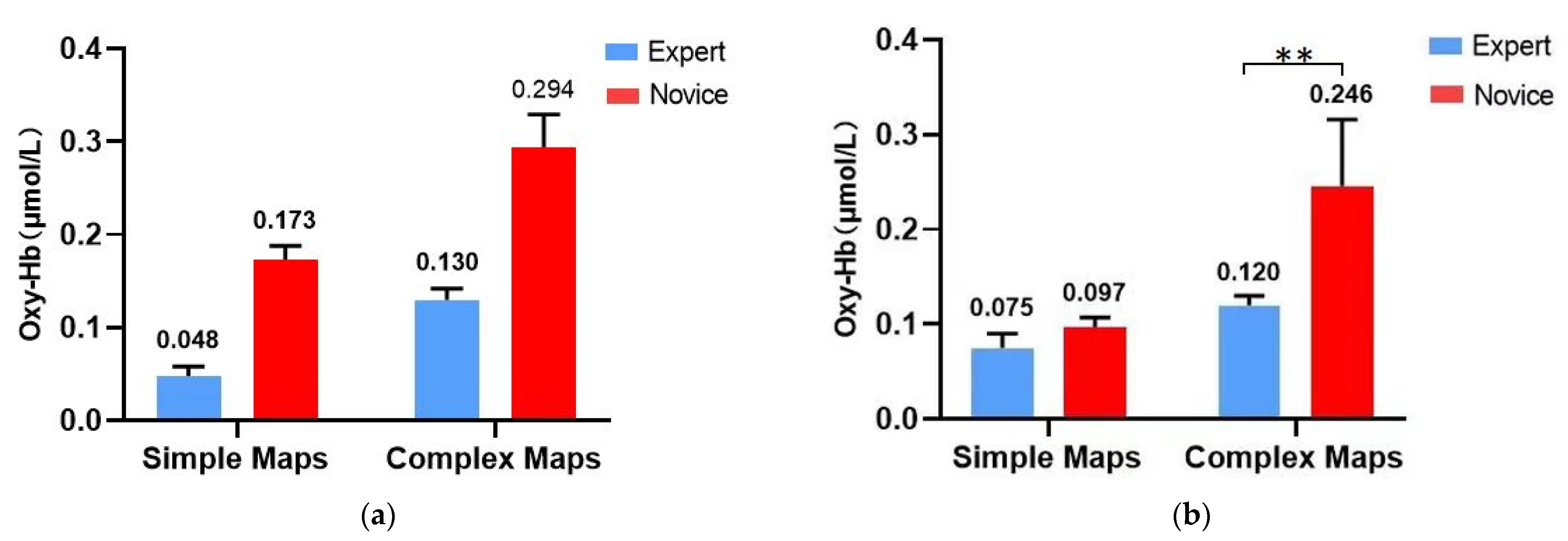
3.3. Results of fNIRS Data for the Orienteering Scene Spatial Memory Task
3.3.1. Left Ventral Lateral Prefrontal Lobe (L-VLPFC)
3.3.2. Left Dorsolateral Prefrontal Lobe (L-DLPFC)
3.3.3. Right Ventral Lateral Prefrontal (R-VLPFC)
3.3.4. Right Dorsolateral Prefrontal Lobe (R-DLPFC)
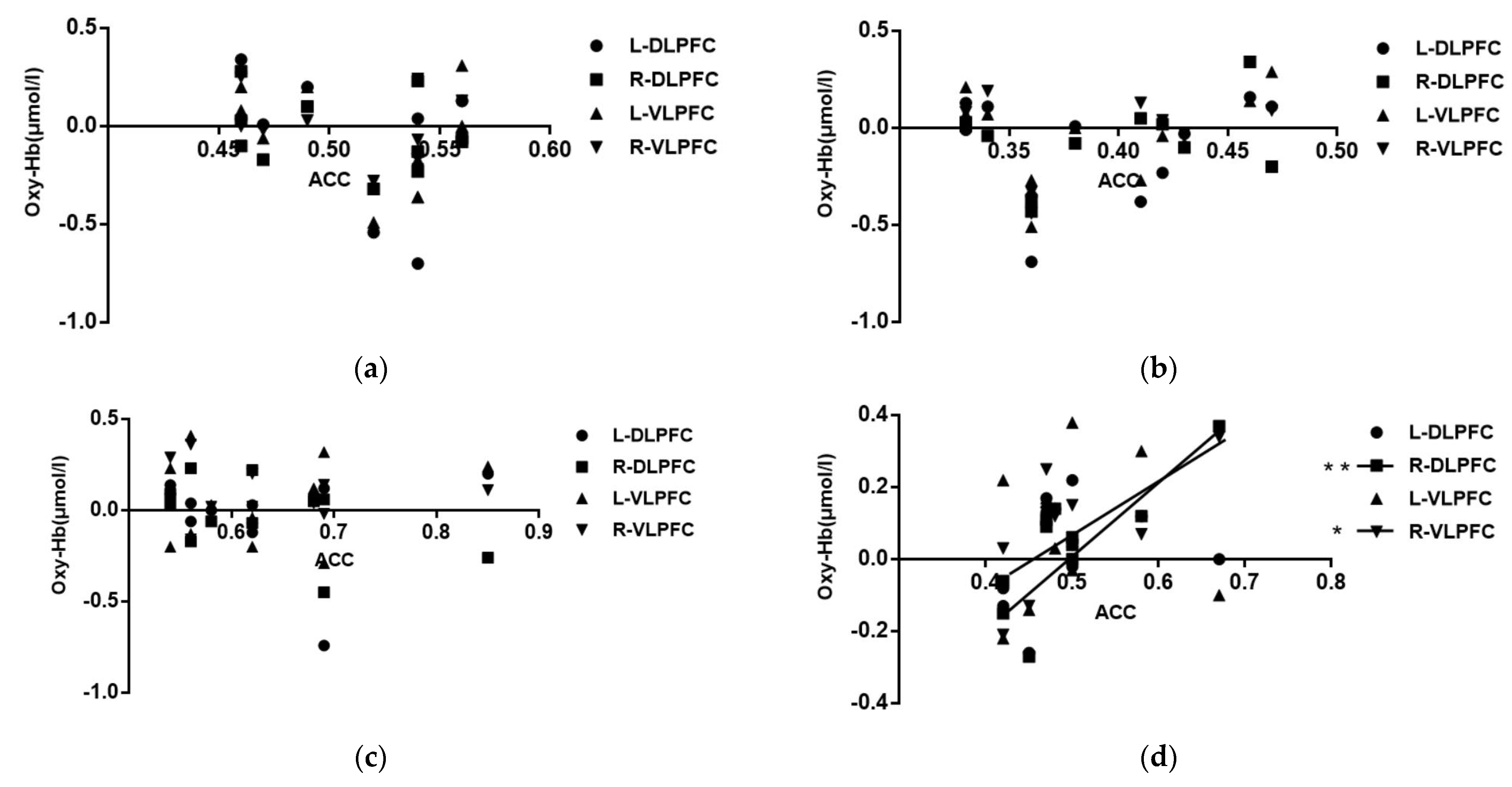
3.4. Correlation Analysis of the Correctness of Spatial Memory Tasks with Different Map Difficulty and the Intensity of Activation of Brain Interest Areas
4. Discussion
4.1. Spatial Memory Analysis of Non-Orienteering Scenes
4.2. Spatial Memory Analysis of Orienteering Scenes
4.2.1. Behavioral Data Analysis of Spatial Memory to Athletes
4.2.2. Brain Activation Analysis of Spatial Memory in Orienteering Athletes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macquet, A.C.; Eccles, D.W.; Barraux, E. What makes an orienteer an expert? A case study of a highly elite orienteer’s concerns in the course of competition. J. Sport. 2012, 30, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Sobotka, R.; Werner, R. A model of performance-determining components in orienteering. Sci. J. Orienteer. 1987, 3, 71–81. [Google Scholar]
- Liu, Y. Research on the Cognitive Processing Characteristics and Skill Training of Map Recognition in Orienteering Athletes; Northeast Normal University: Changchun, China, 2017. [Google Scholar]
- Rosofsky, I. Orienteering: The map sport. Faces People Places Cult. 2005, 22, 18. [Google Scholar]
- Baddeley, A.D. Is working memory still working? Am. Psychol. 2002, 56, 851. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Fu, S.; Cui, S.; Yin, X.; Sun, X.; Qi, X.; Cui, K.; Wang, J.; Ma, L.; Liu, F.Y.; et al. Development of a CRISPR-SaCas9 system for projection- and function-specific gene editing in the rat brain. Sci. Adv. 2020, 18, e6687. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Visual search characteristics of precise map reading by orienteers. PeerJ 2019, 7, e7592. [Google Scholar] [CrossRef]
- Lunze, J. Psychological Information Acceptance and Information Reproduction Abilities of Orienteer’s. Sci. J. Orienteer. 1987, 3, 52–63. [Google Scholar]
- Wang, H. A Study on the Cognitive Processing Characteristics of Badminton Practitioners’ Perceptual Movement Skills; Shanghai Institute of Physical Education: Shanghai, China, 2009. [Google Scholar]
- Shelton, A.L.; Pippitt, H.A. Motion in the mind’s eye: Comparing mental and visual rotation. Cogn. Affect. Behav. Neurosci. 2006, 6, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Dai, K.; Yan, B.; Yang, S. You X. A study of strategies for constructing dynamic spatial representations in pilots. Psychol. Sci. 2010, 33, 87–91. [Google Scholar]
- Liu, Y.; He, J. Visual memory characteristics and processing strategies of orienteering athletes in different task contexts. J. Sport. 2017, 24, 64–70. [Google Scholar]
- Zhu, Y.; Xu, C.; Wang, Y. Study on visual attention strategies of orienteers in different cognitive load scenarios. China Sport. Sci. Technol. 2011, 47, 82–89. [Google Scholar]
- Fu, J. Decision-Making Characteristics and Training Interventions of Badminton Players under Different Difficulty Tasks; Beijing Sport University: Beijing, China, 2015. [Google Scholar]
- Li, L.; Wang, H.; Luo, H.; Zhang, X.; Zhang, R.; Li, X. Interpersonal Neural Synchronization during Cooperative Behavior of Basketball Players: A fNIRS-Based Hyperscanning Study. Front. Hum. Neurosci. 2020, 14, 169. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, T.; Shao, M.; Yan, S.; Yue, G.H.; Jiang, C. Effects of Tai Chi Chuan on Inhibitory Control in Elderly Women: An fNIRS Study. Front. Hum. Neurosci. 2019, 22, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carius, D.; Kenville, R.; Maudrich, D.; Riechel, J.; Lenz, H.; Ragert, P. Cortical processing during table tennis-an fNIRS study in experts and novices. Eur. J. Sport Sci. 2020, 17, 21–26. [Google Scholar] [CrossRef]
- Park, I.; Kim, Y.; Kim, S.K. Athlete-Specific Neural Strategies under Pressure: A fNIRS Pilot Study. Int. J. Environ. Res. Public Health 2020, 17, 8464. [Google Scholar] [CrossRef]
- Mucke, M.; Ludyga, S.; Colledge, F.; Pühse, U.; Gerber, M. The Influence of an Acute Exercise Bout on Adolescents’ Stress Reactivity, Interference Control, and Brain Oxygenation Under Stress. Neurorehabil. Neural Repair 2020, 10, 581965–581978. [Google Scholar] [CrossRef]
- Tore, P.; Corona, F.; Sibilio, M. Orienteering: Spatial navigation strategies and cognitive processes. J. Hum. Sport Exerc. 2015, 10, S507–S514. [Google Scholar]
- Miller, E.K.; Cohen, J.D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 2001, 24, 167–202. [Google Scholar] [CrossRef] [Green Version]
- Simons, J.S.; Spiers, H.J. Prefrontal and medial temporal lobe interactions in long-term memory. Nat. Rev. Neurosci. 2003, 4, 637–648. [Google Scholar] [CrossRef]
- Robert, J.S.; Tirin, M. Selective Attention from Voluntary Control of Neurons in Prefrontal Cortex. Science 2011, 332, 1568–1571. [Google Scholar]
- Balaguer, J.; Spiers, H.; Hassabis, D.; Summerfield, C. Neural mechanisms of hierarchical planning in a virtual subway network. Neuron 2016, 90, 893–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, R.A.; Patai, E.Z.; Julian, J.B.; Spiers, H.J. The cognitive map in humans: Spatial navigation and beyond. Nat. Neurosci. 2017, 20, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Julian, J.B.; Keinath, A.T.; Marchette, S.A.; Epstein, R.A. The neurocognitive basis of spatial reorientation. Curr. Biol. 2018, 28, 1059–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessels, R.P.; van Zandvoort, M.J.; Postma, A.; Kappelle, L.J.; de Haan, E.H. The Corsi Block-Tapping Task: Standardization and normative data. Appl. Neuropsychol. 2000, 7, 252–258. [Google Scholar] [CrossRef]
- Cope, M.; Delpy, D.T. System for long-term measurement of cerebral blood and tissue oxygenation on newborn infants by near infra-red transillumination. Med. Biol. Eng. Comput. 1988, 26, 289–294. [Google Scholar] [CrossRef]
- Müller, M.M.; Ruppert, S.; Schaffner, W.; Matthias, P. A cloned octamer transcription factor stimulates transcription from lymphoid specific promoters in non B cells. Nature 1988, 336, 544–551. [Google Scholar] [CrossRef]
- Barker, T.W.; Horel, J.D. Quasi-Stationary Regimes in the Northern Hemisphere of the NCAR Community Climate Model. J. Clim. 1988, 1, 406–417. [Google Scholar] [CrossRef]
- Abernethy, B.; Baker, J.; Cote, J. Transfer of pattern recall skills may contribute to the development of sport expertise. Appl. Cogn. Psychol. 2005, 9, 705–718. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J. The effect of textual information on badminton players’ pre-set performance. J. Tianjin Sport. Inst. 2007, 6, 487–490. [Google Scholar]
- Ericsson, K.A.; Kintsch, W. Long-term working memory. Psychol. Rev. 1995, 102, 211–245. [Google Scholar] [CrossRef]
- Krause, D.; Richert, B.; Weigelt, M. Neurophysiology of Embodied Mental Rotation-Event-Related Potentials in a Mental Rotation Task with Human Bodies as Compared to Alphanumeric Stimuli. Eur. J. Neurosci. 2021, 54, 5384–5403. [Google Scholar] [CrossRef] [PubMed]
- Bethell-Fox, C.E.; Shepard, R.N. Mental rotation: Effects of stimulus complexity and familiarity. J. Exp. Psychol. Hum. Percept. Perform. 1988, 14, 12–23. [Google Scholar] [CrossRef]
- Lin, Y. Study on the Role of Dorsolateral Prefrontal Lobe in Motor Sequence Learning; Shanghai Institute of Physical Education: Shanghai, China, 2021. [Google Scholar]
- Peng, Y.; Ren, J. Study on the potential relationship between athletic performance and brain EEG characteristics in precision class items. J. Tianjin Sport. Inst. 2021, 36, 332–338. [Google Scholar]
- Wu, Z. Innovation of geometry teaching strategies in junior high school from the perspective of educational neuroscience. J. Educ. Biol. 2021, 9, 249–256. [Google Scholar]
- Plechawska-Wójcik, M.; Tokovarov, M.; Kaczorowska, M.; Zapała, D. A Three-Class Classification of Cognitive Workload Based on EEG Spectral Data. Appl. Sci. 2019, 9, 5340. [Google Scholar] [CrossRef] [Green Version]
- Anna, M.P.; Elisenda, B.; Noelia, V.C.; María-Jesús, F.C.; Maria, A.P.; César, A. Long-term brain effects of N-back training: An fMRI study. Brain Imaging Behav. 2019, 13, 1115–1127. [Google Scholar]
- Ma, Y. An Experimental Study of Physical Motor Function Training to Promote Cognitive Executive Function in Older Adults; Capital Institute of Physical Education and Sports: Beijing, China, 2021. [Google Scholar]
- Xu, Z.; Niu, Y.; Wen, X.; Zhou, Y.; Wang, P.; Wu, X.; Zhang, D. A review of cognitive load assessment based on EEG signals. Aerosp. Med. Med. Eng. 2021, 34, 339–348. [Google Scholar]
- Babiloni, C.; Vecchio, F.; Infarinato, F.; Buffo, P.; Marzano, N.; Spada, D.; Rossi, S.; Bruni, I.; Rossini, P.M.; Perani, D. Simultaneous recording of electroencephalographic data in musicians playing in ensemble. Cortex 2011, 47, 1082–1090. [Google Scholar] [CrossRef]
- Jin, F.; Flombaum, J.I.; Mccandliss, B.D.; Thomas, K.M.; Posner, M.I. Cognitive and Brain Consequences of Conflic. NeuroImage 2003, 18, 42–57. [Google Scholar]
- Straten, A.; Huyser, C.; Wolters, L.; Denys, D.; van Wingen, G. Long-Term Effects of Cognitive Behavioral Therapy on Planning and Prefrontal Cortex Function in Pediatric Obsessive-Compulsive Disorder. Eur. Neuropsychopharmacol. 2018, 28, 67–68. [Google Scholar]
- Wilson, C.R.E.; Gaffan, D.; Mitchell, A.S.; Baxter, M.G. Neurotoxic lesions of ventrolateral prefrontal cortex impair object-in-place scene memory. Eur. J. Neurosci. 2007, 25, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Marika, C.; Stefania, L.; Alessia, B.; Marco, F.; Laura, P.; Valentina, Q. Does ventrolateral prefrontal cortex help in searching for the lost key? Evidence from an fNIRS study. Brain Imaging Behav. 2017, 12, 785–797. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Height | Weight | BMI | Age |
|---|---|---|---|---|
| Novice Group | 170.4 ± 6.60 | 65.2 ± 11.93 | 22.31 ± 2.80 | 22.33 ± 1.63 |
| Expert Group | 170.3 ± 8.00 | 62.3 ± 14.01 | 21.24 ± 3.01 | 21.40 ± 2.06 |
| Brain Regions | Corresponding Channels |
|---|---|
| Left ventral lateral prefrontal cortical area (L-VLPFC) | Ch1, Ch2, Ch3, Ch4, Ch5, Ch7 |
| Left dorsolateral prefrontal cortical area (L-DLPFC) | Ch6, Ch8, Ch9, Ch10, Ch11, Ch12, Ch13 |
| Right ventral lateral prefrontal cortical area (R-VLPFC) | Ch23, Ch24, Ch25, Ch26, Ch27, Ch28 |
| Right dorsolateral prefrontal cortical area (R-DLPFC) | Ch16, Ch17, Ch18, Ch19, Ch20, Ch21, Ch22 |
| Expert Group | Novice Group | |
|---|---|---|
| Spatial memory breadth score | 5.97 ± 0.09 | 5.65 ± 0.08 |
| Novice Group | Expert Group | |||
|---|---|---|---|---|
| Map Difficulty | Simple Map | Complex Map | Simple Map | COMPLEX MAP |
| Correctness rate | 0.50 ± 0.03 ** | 0.39 ± 0.03 ** | 0.61 ± 0.03 ** | 0.48 ± 0.03 ** |
| Reaction time | 10,832.85 ± 252.97 ** | 11,470.71 ± 252.05 ** | 9218.46 ± 431.94 ** | 9605.31 ± 312.54 ** |
| Novice Group | Expert Group | |||
|---|---|---|---|---|
| ROI | Simple Map | Complex Map | Simple Map | Complex Map |
| L-VLPFC | 0.173 ± 0.016 | 0.294 ± 0.031 | 0.048 ± 0.011 | 0.130 ± 0.013 |
| L-DLPFC | 0.097 ± 0.011 | 0.246 ± 0.074 | 0.075 ± 0.017 | 0.120 ± 0.014 |
| R-VLPFC | 0.198 ± 0.011 | 0.417 ± 0.0348 | 0.054 ± 0.023 | 0.114 ± 0.033 |
| R-DLPFC | 0.177 ± 0.011 | 0.317 ± 0.020 | 0.025 ± 0.010 | 0.134 ± 0.018 |
| Area of Interest | Group | Type of Map | |
|---|---|---|---|
| Simple Map | Complex Map | ||
| L-DLPFC | Expert Group | −0.032 | 0.271 |
| Novice Group | 0.259 | −0.296 | |
| R-DLPFC | Expert Group | −0.483 | 0.792 ** |
| Novice Group | 0.252 | −0.071 | |
| L-VLPFC | Expert Group | 0.178 | 0.047 |
| Novice Group | 0.266 | −0.396 | |
| R-VLPFC | Expert Group | −0.145 | 0.657 * |
| Novice Group | 0.317 | 0.059 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Lu, S.; Liu, J.; Zhao, M.; Chao, Y.; Kang, P. A Characterization of Brain Area Activation in Orienteers with Different Map-Recognition Memory Ability Task Levels—Based on fNIRS Evidence. Brain Sci. 2022, 12, 1561. https://doi.org/10.3390/brainsci12111561
Liu Y, Lu S, Liu J, Zhao M, Chao Y, Kang P. A Characterization of Brain Area Activation in Orienteers with Different Map-Recognition Memory Ability Task Levels—Based on fNIRS Evidence. Brain Sciences. 2022; 12(11):1561. https://doi.org/10.3390/brainsci12111561
Chicago/Turabian StyleLiu, Yang, Su Lu, Jingru Liu, Mingsheng Zhao, Yue Chao, and Pengyang Kang. 2022. "A Characterization of Brain Area Activation in Orienteers with Different Map-Recognition Memory Ability Task Levels—Based on fNIRS Evidence" Brain Sciences 12, no. 11: 1561. https://doi.org/10.3390/brainsci12111561