
Impact of Partial Volume Correction on [18F]GE-180 PET Quantification in Subcortical Brain Regions of Patients with Corticobasal Syndrome

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Study Population and Clinical Assessment
2.2. TSPO PET Acquisition
2.3. Data Analysis and PVEC
2.4. rs6971 Single Nucleotide Polymorphism
2.5. Statistical Analysis
3. Results
3.1. Demographics and Clinical Data
3.2. Subcortical Brain Regions in CBS Patients Show Volume Loss When Compared to Controls
3.3. Region-Based PVEC in TSPO PET in Subcortical Brain Regions
3.4. Impact of PVEC on SUVr Differences and Effect Sizes for the Comparison of CBS and Controls
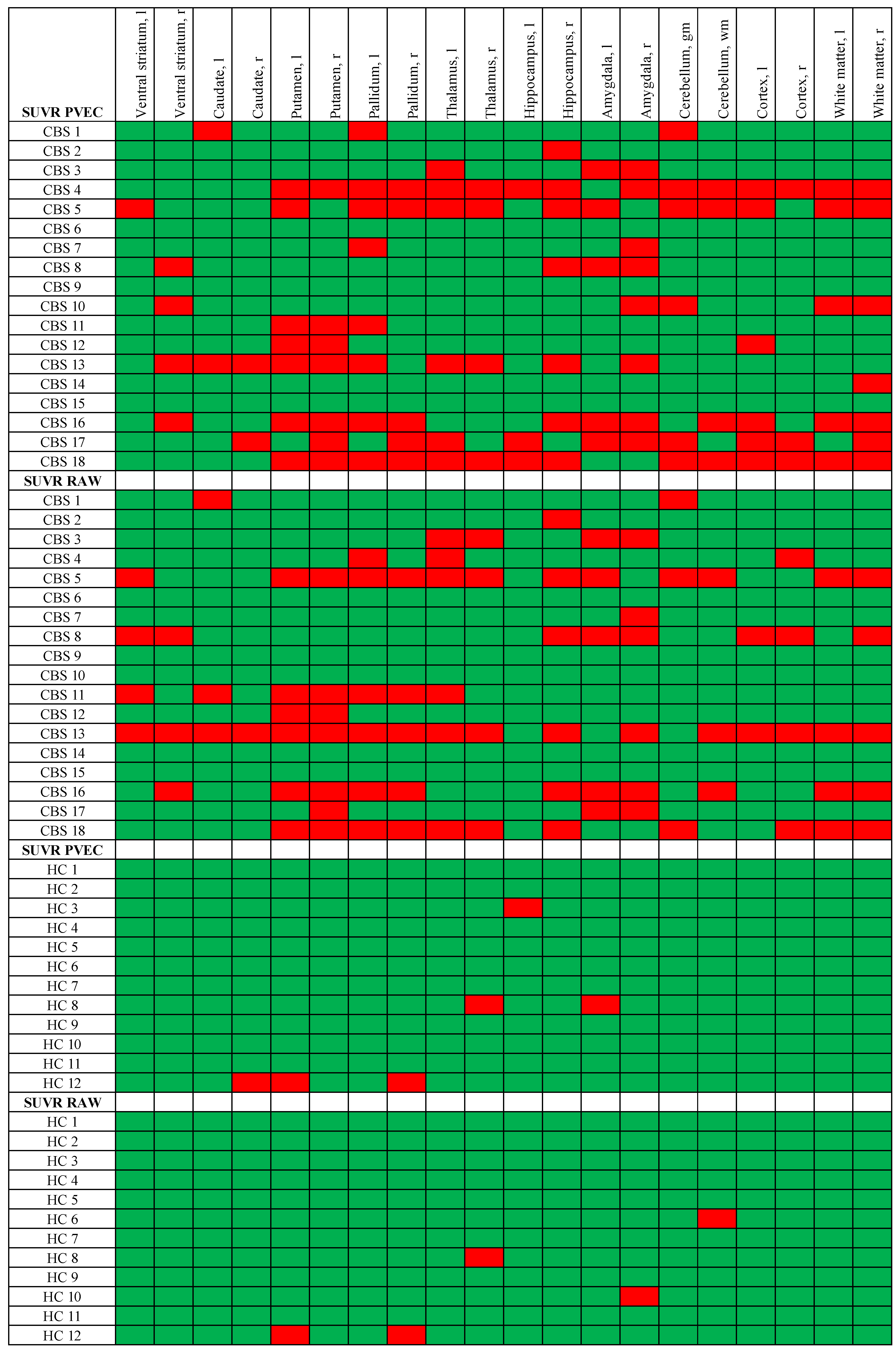
3.5. Single Region Positivity of TSPO PET before and after PVEC
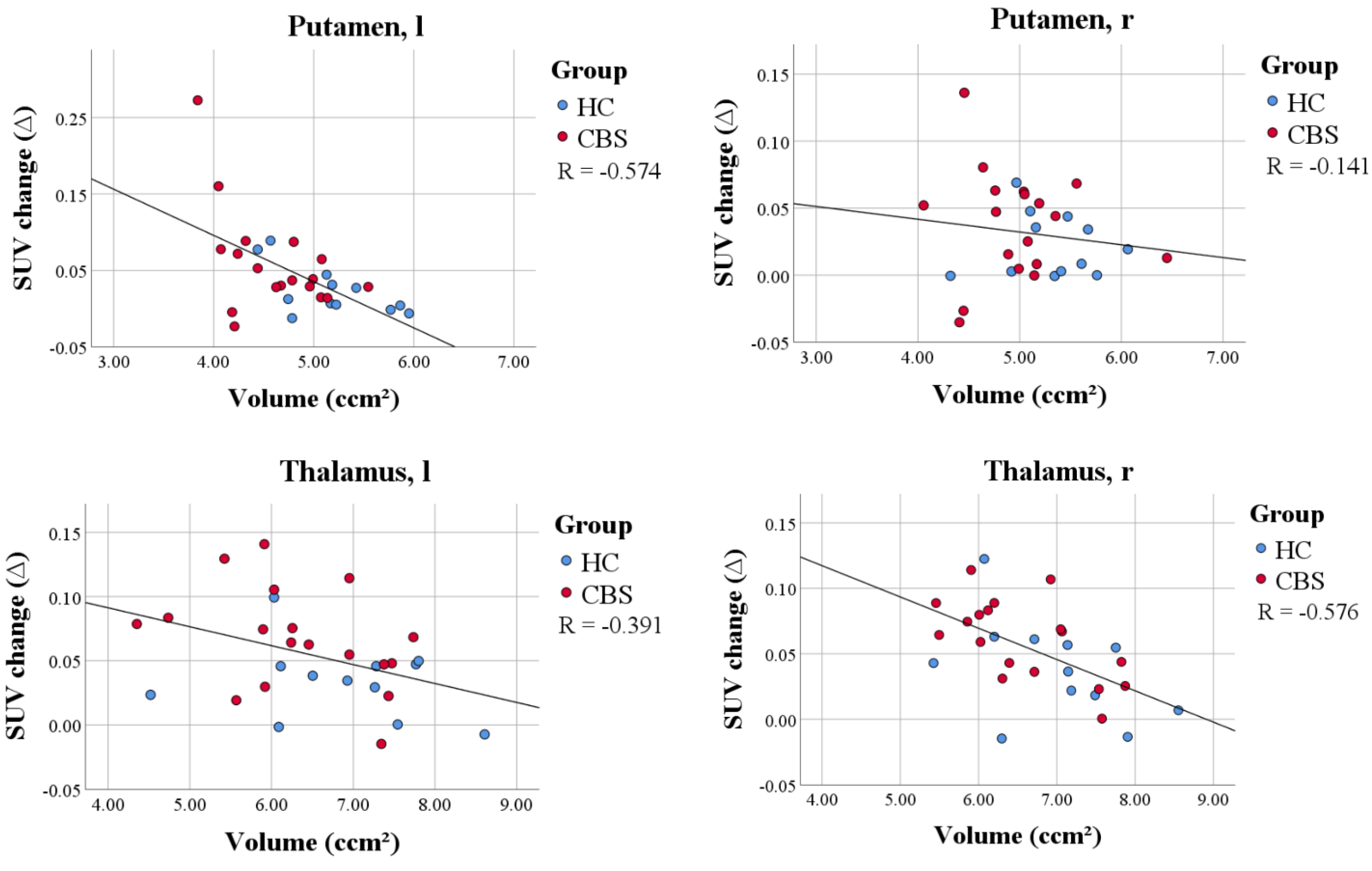
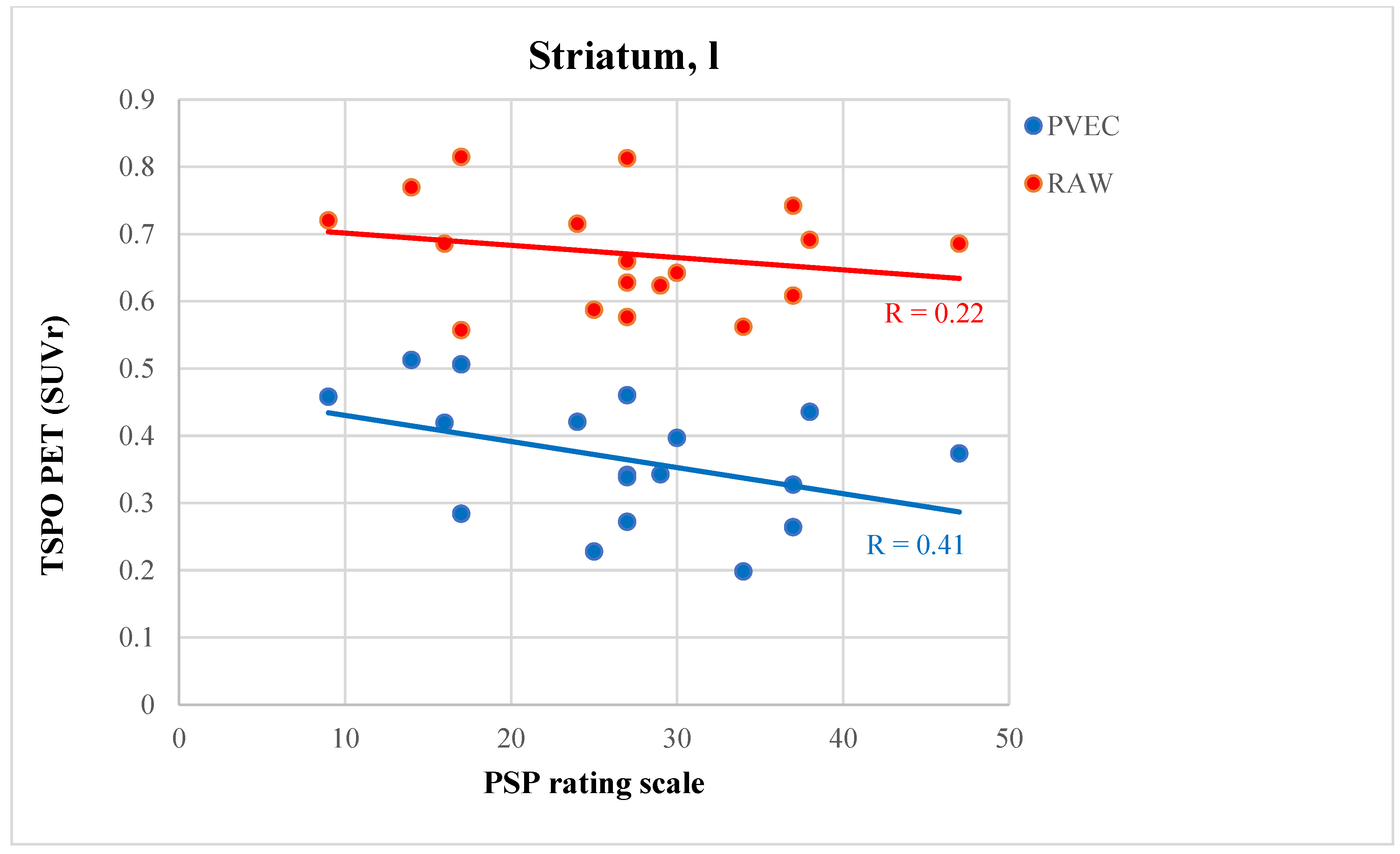
3.6. PVEC Influence on the Association of Disease Parameter with TSPO Labeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrer, I.; López-González, I.; Carmona, M.; Arregui, L.; Dalfó, E.; Torrejón-Escribano, B.; Diehl, R.; Kovacs, G.G. Glial and neuronal tau pathology in tauopathies: Characterization of disease-specific phenotypes and tau pathology progression. J. Neuropathol. Exp. Neurol. 2014, 73, 81–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chahine, L.M.; Rebeiz, T.; Rebeiz, J.J.; Grossman, M.; Gross, R.G. Corticobasal syndrome: Five new things. Neurol. Clin. Pract. 2014, 4, 304–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, M.J.; Litvan, I.; Lang, A.E.; Bak, T.H.; Bhatia, K.P.; Borroni, B.; Boxer, A.L.; Dickson, D.W.; Grossman, M.; Hallett, M. Criteria for the diagnosis of corticobasal degeneration. Neurology 2013, 80, 496–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouri, N.; Whitwell, J.L.; Josephs, K.A.; Rademakers, R.; Dickson, D.W. Corticobasal degeneration: A pathologically distinct 4R tauopathy. Nat. Rev. Neurol. 2011, 7, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Simón, D.; García-García, E.; Royo, F.; Falcón-Pérez, J.M.; Avila, J. Proteostasis of tau. Tau overexpression results in its secretion via membrane vesicles. FEBS Lett. 2012, 586, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Pooler, A.M.; Phillips, E.C.; Lau, D.H.; Noble, W.; Hanger, D.P. Physiological release of endogenous tau is stimulated by neuronal activity. EMBO Rep. 2013, 14, 389–394. [Google Scholar] [CrossRef]
- Gómez-Ramos, A.; Díaz-Hernández, M.; Cuadros, R.; Hernández, F.; Avila, J. Extracellular tau is toxic to neuronal cells. FEBS Lett. 2006, 580, 4842–4850. [Google Scholar] [CrossRef] [Green Version]
- Calafate, S.; Buist, A.; Miskiewicz, K.; Vijayan, V.; Daneels, G.; de Strooper, B.; de Wit, J.; Verstreken, P.; Moechars, D. Synaptic Contacts Enhance Cell-to-Cell Tau Pathology Propagation. Cell Rep. 2015, 11, 1176–1183. [Google Scholar] [CrossRef] [Green Version]
- Ransohoff, R.M. How neuroinflammation contributes to neurodegeneration. Science 2016, 353, 777–783. [Google Scholar] [CrossRef]
- Leyns, C.E.G.; Holtzman, D.M. Glial contributions to neurodegeneration in tauopathies. Mol. Neurodegener. 2017, 12, 50. [Google Scholar] [CrossRef]
- Kettenmann, H.; Hanisch, U.K.; Noda, M.; Verkhratsky, A. Physiology of microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Kahlson, M.A.; Colodner, K.J. Glial Tau Pathology in Tauopathies: Functional Consequences. J. Exp. Neurosci. 2015, 9, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ising, C.; Venegas, C.; Zhang, S.; Scheiblich, H.; Schmidt, S.V.; Vieira-Saecker, A.; Schwartz, S.; Albasset, S.; McManus, R.M.; Tejera, D.; et al. NLRP3 inflammasome activation drives tau pathology. Nature 2019, 575, 669–673. [Google Scholar] [CrossRef]
- Asai, H.; Ikezu, S.; Tsunoda, S.; Medalla, M.; Luebke, J.; Haydar, T.; Wolozin, B.; Butovsky, O.; Kügler, S.; Ikezu, T. Depletion of microglia and inhibition of exosome synthesis halt tau propagation. Nat. Neurosci. 2015, 18, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- von Bernhardi, R.; Eugenín-von Bernhardi, L.; Eugenín, J. Microglial cell dysregulation in brain aging and neurodegeneration. Front. Aging Neurosci. 2015, 7, 124. [Google Scholar] [CrossRef] [Green Version]
- Stefaniak, J.; O’Brien, J. Imaging of neuroinflammation in dementia: A review. J. Neurol. Neurosurg. Psychiatry 2016, 87, 21–28. [Google Scholar] [CrossRef]
- Liu, B.; Le, K.X.; Park, M.A.; Wang, S.; Belanger, A.P.; Dubey, S.; Frost, J.L.; Holton, P.; Reiser, V.; Jones, P.A.; et al. In Vivo Detection of Age- and Disease-Related Increases in Neuroinflammation by 18F-GE180 TSPO MicroPET Imaging in Wild-Type and Alzheimer’s Transgenic Mice. J. Neurosci. 2015, 35, 15716–15730. [Google Scholar] [CrossRef] [Green Version]
- Parhizkar, S.; Arzberger, T.; Brendel, M.; Kleinberger, G.; Deussing, M.; Focke, C.; Nuscher, B.; Xiong, M.; Ghasemigharagoz, A.; Katzmarski, N. Loss of TREM2 function increases amyloid seeding but reduces plaque-associated ApoE. Nat. Neurosci. 2019, 22, 191–204. [Google Scholar] [CrossRef]
- Gerhard, A.; Trender-Gerhard, I.; Turkheimer, F.; Quinn, N.P.; Bhatia, K.P.; Brooks, D.J. In vivo imaging of microglial activation with [11C](R)-PK11195 PET in progressive supranuclear palsy. Mov. Disord. 2006, 21, 89–93. [Google Scholar] [CrossRef]
- Passamonti, L.; Rodríguez, P.V.; Hong, Y.T.; Allinson, K.S.J.; Bevan-Jones, W.R.; Williamson, D.; Jones, P.S.; Arnold, R.; Borchert, R.J.; Surendranathan, A.; et al. [(11)C]PK11195 binding in Alzheimer disease and progressive supranuclear palsy. Neurology 2018, 90, e1989–e1996. [Google Scholar] [CrossRef] [Green Version]
- Palleis, C.; Sauerbeck, J.; Beyer, L.; Harris, S.; Schmitt, J.; Morenas-Rodriguez, E.; Finze, A.; Nitschmann, A.; Ruch-Rubinstein, F.; Eckenweber, F.; et al. In Vivo Assessment of Neuroinflammation in 4-Repeat Tauopathies. Mov. Disord. 2021, 36, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.A.; Erlandsson, K.; Modat, M.; Thurfjell, L.; Vandenberghe, R.; Ourselin, S.; Hutton, B.F. The importance of appropriate partial volume correction for PET quantification in Alzheimer’s disease. Eur. J. Nucl. Med. Mol. Imaging 2011, 38, 1104–1119. [Google Scholar] [CrossRef] [PubMed]
- Brendel, M.; Delker, A.; Rötzer, C.; Böning, G.; Carlsen, J.; Cyran, C.; Mille, E.; Gildehaus, F.J.; Cumming, P.; Baumann, K.; et al. Impact of partial volume effect correction on cerebral β-amyloid imaging in APP-Swe mice using [(18)F]-florbetaben PET. Neuroimage 2014, 84, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Brendel, M.; Högenauer, M.; Delker, A.; Sauerbeck, J.; Bartenstein, P.; Seibyl, J.; Rominger, A. Improved longitudinal [(18)F]-AV45 amyloid PET by white matter reference and VOI-based partial volume effect correction. Neuroimage 2015, 108, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Brendel, M.; Reinisch, V.; Kalinowski, E.; Levin, J.; Delker, A.; Därr, S.; Pogarell, O.; Förster, S.; Bartenstein, P.; Rominger, A.; et al. Hypometabolism in Brain of Cognitively Normal Patients with Depressive Symptoms is Accompanied by Atrophy-Related Partial Volume Effects. Curr. Alzheimer Res. 2016, 13, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Rullmann, M.; McLeod, A.; Grothe, M.J.; Sabri, O.; Barthel, H. Reshaping the Amyloid Buildup Curve in Alzheimer Disease? Partial-Volume Effect Correction of Longitudinal Amyloid PET Data. J. Nucl. Med. 2020, 61, 1820–1824. [Google Scholar] [CrossRef]
- Rullmann, M.; Dukart, J.; Hoffmann, K.T.; Luthardt, J.; Tiepolt, S.; Patt, M.; Gertz, H.J.; Schroeter, M.L.; Seibyl, J.; Schulz-Schaeffer, W.J.; et al. Partial-Volume Effect Correction Improves Quantitative Analysis of 18F-Florbetaben β-Amyloid PET Scans. J. Nucl. Med. 2016, 57, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Wickstrøm, T.; Clarke, A.; Gausemel, I.; Horn, E.; Jørgensen, K.; Khan, I.; Mantzilas, D.; Rajanayagam, T.; In’t Veld, D.J.; Trigg, W. The development of an automated and GMP compliant FASTlab™ Synthesis of [18F] GE-180; a radiotracer for imaging translocator protein (TSPO). J. Label. Compd. Radiopharm. 2014, 57, 42–48. [Google Scholar] [CrossRef]
- Hammers, A.; Allom, R.; Koepp, M.J.; Free, S.L.; Myers, R.; Lemieux, L.; Mitchell, T.N.; Brooks, D.J.; Duncan, J.S. Three-dimensional maximum probability atlas of the human brain, with particular reference to the temporal lobe. Hum. Brain Mapp. 2003, 19, 224–247. [Google Scholar] [CrossRef]
- Rousset, O.G.; Ma, Y.; Evans, A.C. Correction for partial volume effects in PET: Principle and validation. J. Nucl. Med. 1998, 39, 904–911. [Google Scholar]
- Albert, N.L.; Unterrainer, M.; Fleischmann, D.; Lindner, S.; Vettermann, F.; Brunegraf, A.; Vomacka, L.; Brendel, M.; Wenter, V.; Wetzel, C. TSPO PET for glioma imaging using the novel ligand 18 F-GE-180: First results in patients with glioblastoma. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 2230–2238. [Google Scholar] [CrossRef] [PubMed]
- Vettermann, F.J.; Harris, S.; Schmitt, J.; Unterrainer, M.; Lindner, S.; Rauchmann, B.S.; Palleis, C.; Weidinger, E.; Beyer, L.; Eckenweber, F.; et al. Impact of TSPO Receptor Polymorphism on [(18)F]GE-180 Binding in Healthy Brain and Pseudo-Reference Regions of Neurooncological and Neurodegenerative Disorders. Life 2021, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Rohrer, J.D.; Lashley, T.; Schott, J.M.; Warren, J.E.; Mead, S.; Isaacs, A.M.; Beck, J.; Hardy, J.; de Silva, R.; Warrington, E.; et al. Clinical and neuroanatomical signatures of tissue pathology in frontotemporal lobar degeneration. Brain 2011, 134, 2565–2581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, M.; Brendel, M.; Rizk-Jackson, A.; Rominger, A.; Bartenstein, P.; Schuff, N.; Weiner, M.W. Reduced FDG-PET brain metabolism and executive function predict clinical progression in elderly healthy subjects. Neuroimage Clin. 2014, 4, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Schöll, M.; Lockhart, S.N.; Schonhaut, D.R.; O’Neil, J.P.; Janabi, M.; Ossenkoppele, R.; Baker, S.L.; Vogel, J.W.; Faria, J.; Schwimmer, H.D.; et al. PET Imaging of Tau Deposition in the Aging Human Brain. Neuron 2016, 89, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Benzinger, T.L.; Su, Y.; Christensen, J.; Friedrichsen, K.; Aldea, P.; McConathy, J.; Cairns, N.J.; Fagan, A.M.; Morris, J.C.; et al. Evaluation of Tau Imaging in Staging Alzheimer Disease and Revealing Interactions Between β-Amyloid and Tauopathy. JAMA Neurol. 2016, 73, 1070–1077. [Google Scholar] [CrossRef]
- Parmera, J.B.; de Almeida, I.J.; de Oliveira, M.C.B.; Silagi, M.L.; de Godoi Carneiro, C.; Studart-Neto, A.; Ono, C.R.; Reis Barbosa, E.; Nitrini, R.; Buchpiguel, C.A.; et al. Metabolic and Structural Signatures of Speech and Language Impairment in Corticobasal Syndrome: A Multimodal PET/MRI Study. Front. Neurol. 2021, 12, 702052. [Google Scholar] [CrossRef]
- Boxer, A.L.; Geschwind, M.D.; Belfor, N.; Gorno-Tempini, M.L.; Schauer, G.F.; Miller, B.L.; Weiner, M.W.; Rosen, H.J. Patterns of brain atrophy that differentiate corticobasal degeneration syndrome from progressive supranuclear palsy. Arch. Neurol. 2006, 63, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Tokumaru, A.M.; O’Uchi, T.; Kuru, Y.; Maki, T.; Murayama, S.; Horichi, Y. Corticobasal degeneration: MR with histopathologic comparison. AJNR Am. J. Neuroradiol. 1996, 17, 1849–1852. [Google Scholar]
- Franceschi, A.M.; Clifton, M.; Naser-Tavakolian, K.; Ahmed, O.; Cruciata, G.; Bangiyev, L.; Clouston, S.; Franceschi, D. ((18)F)-Fluorodeoxyglucose positron emission tomography/magnetic resonance imaging assessment of hypometabolism patterns in clinical phenotypes of suspected corticobasal degeneration. World J. Nucl. Med. 2021, 20, 176–184. [Google Scholar] [CrossRef]
- Tournier, B.B.; Tsartsalis, S.; Ceyzériat, K.; Garibotto, V.; Millet, P. In Vivo TSPO Signal and Neuroinflammation in Alzheimer’s Disease. Cells 2020, 9, 1941. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, J.; Sarazin, M.; Bottlaender, M. In vivo PET imaging of neuroinflammation in Alzheimer’s disease. J. Neural Transm. 2018, 125, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Schain, M.; Kreisl, W.C. Neuroinflammation in Neurodegenerative Disorders-a Review. Curr. Neurol. Neurosci. Rep. 2017, 17, 25. [Google Scholar] [CrossRef] [PubMed]
- Sacher, C.; Blume, T.; Beyer, L.; Peters, F.; Eckenweber, F.; Sgobio, C.; Deussing, M.; Albert, N.L.; Unterrainer, M.; Lindner, S.; et al. Longitudinal PET Monitoring of Amyloidosis and Microglial Activation in a Second-Generation Amyloid-β Mouse Model. J. Nucl. Med. 2019, 60, 1787–1793. [Google Scholar] [CrossRef]
- Brendel, M.; Kleinberger, G.; Probst, F.; Jaworska, A.; Overhoff, F.; Blume, T.; Albert, N.L.; Carlsen, J.; Lindner, S.; Gildehaus, F.J.; et al. Increase of TREM2 during Aging of an Alzheimer’s Disease Mouse Model Is Paralleled by Microglial Activation and Amyloidosis. Front. Aging Neurosci. 2017, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Deussing, M.; Blume, T.; Vomacka, L.; Mahler, C.; Focke, C.; Todica, A.; Unterrainer, M.; Albert, N.L.; Lindner, S.; von Ungern-Sternberg, B.; et al. Data on specificity of [(18)F]GE180 uptake for TSPO expression in rodent brain and myocardium. Data Brief 2018, 19, 331–336. [Google Scholar] [CrossRef]
- Sridharan, S.; Raffel, J.; Nandoskar, A.; Record, C.; Brooks, D.J.; Owen, D.; Sharp, D.; Muraro, P.A.; Gunn, R.; Nicholas, R. Confirmation of specific binding of the 18-kDa translocator protein (TSPO) radioligand [18F] GE-180: A blocking study using XBD173 in multiple sclerosis normal appearing white and grey matter. Mol. Imaging Biol. 2019, 21, 935–944. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| CBS | HC | |
|---|---|---|
| n | 18 | 12 |
| Age (y) | 66.00 ± 7.46 | 70.42 ± 7.45 |
| Sex | ♀ 11/♂ 7 | ♀ 6/♂ 6 |
| rs6971 | HAB: 14/MAB: 4 | HAB: 4/MAB: 8 |
| PSP rating scale | 26.78 ± 9.45 | n.a. |
| Disease duration (m) | 30.83 ± 19.47 | n.a. |
| MoCA | 23.39 ± 4.36 | 29.00 ± 1.00 |
| SEADL | 63.89 ± 13.80 | n.a. |
| Region | Mean vol. CBS (ccm ± SD) | Mean vol. HC (ccm ± SD) | p-Value |
|---|---|---|---|
| Ventral striatum, l | 0.47 ± 0.06 | 0.48 ± 0.05 | 0.498 |
| Ventral striatum, r | 0.46 ± 0.04 | 0.49 ± 0.06 | 0.041 |
| Caudate, l | 4.11 ± 0.64 | 4.35 ± 0.69 | 0.374 |
| Caudate, r | 4.29 ± 0.60 | 4.77 ± 0.60 | 0.086 |
| Putamen, l | 4.61 ± 0.47 | 5.19 ± 0.48 | 0.005 |
| Putamen, r | 4.97 ± 0.53 | 5.32 ± 0.44 | 0.038 |
| Pallidum, l | 1.52 ± 0.22 | 1.79 ± 0.27 | 0.015 |
| Pallidum, r | 1.65 ± 0.15 | 1.81 ± 0.21 | 0.065 |
| Thalamus, l | 6.34 ± 0.97 | 6.87 ± 1.04 | 0.133 |
| Thalamus, r | 6.57 ± 0.78 | 6.99 ± 0.85 | 0.175 |
| Hippocampus, l | 3.95 ± 0.49 | 3.89 ± 0.32 | 0.703 |
| Hippocampus, r | 3.86 ± 0.35 | 3.97 ± 0.36 | 0.397 |
| Amygdala, l | 1.66 ± 0.19 | 1.71 ± 0.20 | 0.611 |
| Amygdala, r | 1.78 ± 0.23 | 1.75 ± 0.27 | 0.882 |
| Cerebellum, gm | 102.50 ± 23.27 | 101.76 ± 27.23 | 0.866 |
| Cerebellum, wm | 36.24 ± 8.47 | 37.42 ± 10.83 | 0.204 |
| Cortex, l | 298.70 ± 22.08 | 325.59 ± 36.22 | 0.057 |
| Cortex, r | 302.01 ± 25.64 | 318.41 ± 17.91 | 0.047 |
| White matter, l | 237.06 ± 23.70 | 248.52 ± 25.66 | 0.374 |
| White matter, r | 239.64 ± 20.07 | 248.18 ± 22.34 | 0.290 |
| Anterior temporal lobe, l | 4.06 ± 1.53 | 3.92 ± 1.54 | 0.398 |
| Anterior temporal lobe, r | 4.14 ± 1.54 | 3.99 ± 1.53 | 0.353 |
| CBS | HC | |||||
|---|---|---|---|---|---|---|
| Region | Mean SUV before PVEC ± SD | Mean SUV after PVEC ± SD | p | Mean SUV before PVEC ± SD | Mean SUV after PVEC ± SD | p |
| Ventral striatum, l | 0.888 ± 0.20 | 1.022 ± 0.34 | 0.177 | 0.904 ± 0.21 | 1.112 ± 0.31 | 0.002 |
| Ventral striatum, r | 0.919 ± 0.20 | 1.107 ± 0.37 | 0.149 | 0.948 ± 0.22 | 1.185 ± 0.35 | 0.002 |
| Caudate, l | 0.594 ± 0.12 | 0.445 ± 0.12 | 0.001 | 0.582 ± 0.13 | 0.387 ± 0.12 | 0.002 |
| Caudate, r | 0.584 ± 0.13 | 0.432 ± 0.18 | 0.017 | 0.619 ± 0.16 | 0.465 ± 0.16 | 0.002 |
| Putamen, l | 0.875 ± 0.19 | 0.934 ± 0.23 | 0.463 | 0.805 ± 0.19 | 0.828 ± 0.20 | 0.028 |
| Putamen, r | 0.849 ± 0.17 | 0.886 ± 0.20 | 0.287 | 0.810 ± 0.20 | 0.832 ± 0.21 | 0.010 |
| Pallidum, l | 0.890 ± 0.18 | 0.979 ± 0.21 | 0.084 | 0.807 ± 0.20 | 0.862 ± 0.23 | 0.002 |
| Pallidum, r | 0.878 ± 0.17 | 0.969 ± 0.20 | 0.068 | 0.818 ± 0.19 | 0.883 ± 0.21 | 0.002 |
| Thalamus, l | 0.955 ± 0.19 | 1.021 ± 0.22 | 0.149 | 0.907 ± 0.23 | 0.940 ± 0.25 | 0.008 |
| Thalamus, r | 0.938 ± 0.17 | 0.999 ± 0.19 | 0.149 | 0.916 ± 0.25 | 0.954 ± 0.28 | 0.008 |
| Hippocampus, l | 0.834 ± 0.16 | 0.852 ± 0.19 | 0.619 | 0.895 ± 0.23 | 0.937 ± 0.27 | 0.010 |
| Hippocampus, r | 0.833 ± 0.16 | 0.855 ± 0.19 | 0.723 | 0.845 ± 0.19 | 0.848 ± 0.20 | 0.875 |
| Amygdala, l | 0.835 ± 0.18 | 0.815 ± 0.22 | 0.831 | 0.858 ± 0.22 | 0.811 ± 0.23 | 0.006 |
| Amygdala, r | 0.846 ± 0.17 | 0.834 ± 0.21 | 0.943 | 0.848 ± 0.21 | 0.785 ± 0.21 | 0.002 |
| Cerebellum, gm | 0.880 ± 0.15 | 1.002 ± 0.19 | 0.015 | 0.921 ± 0.24 | 1.033 ± 0.27 | 0.002 |
| Cerebellum, wm | 0.826 ± 0.16 | 0.800 ± 0.17 | 0.653 | 0.852 ± 0.20 | 0.817 ± 0.19 | 0.008 |
| Cortex, l | 0.860 ± 0.16 | 1.049 ± 0.21 | 0.006 | 0.903 ± 0.22 | 1.083 ± 0.27 | 0.002 |
| Cortex, r | 0.871 ± 0.16 | 1.073 ± 0.21 | 0.006 | 0.927 ± 0.23 | 1.129 ± 0.28 | 0.002 |
| White matter, l | 0.760 ± 0.14 | 0.664 ± 0.13 | 0.062 | 0.765 ± 0.17 | 0.648 ± 0.14 | 0.002 |
| White matter, r | 0.759 ± 0.14 | 0.659 ± 0.13 | 0.084 | 0.771 ± 0.18 | 0.643 ± 0.14 | 0.002 |
| Anterior temporal lobe, l | 0.906 ± 0.16 | 1.261 ± 0.21 | 0.001 | 0.998 ± 0.24 | 1.460 ± 0.36 | 0.002 |
| Anterior temporal lobe, r | 0.870 ± 0.16 | 1.193 ± 0.23 | 0.001 | 0.938 ± 0.24 | 1.320 ± 0.41 | 0.002 |
| Regions | Before PVEC—CBS vs. HC (%) | r | After PVEC—CBS vs. HC (%) | r | ∆ (%) |
|---|---|---|---|---|---|
| Ventral striatum, l | 5.88 | 0.2 | 0.72 | 0.1 | −5.16 |
| Ventral striatum, r | 5.72 | 0.2 | 5.73 | 0.1 | 0.01 |
| Caudate nucl., l | 10.44 | 0.3 | 27.32 | 0.4 | 16.88 |
| Caudate nucl., r | 2.67 | 0.1 | 5.56 | 0.1 | 2.89 |
| Putamen, l | 18.4 | 0.5 | 27.25 | 0.5 | 8.85 |
| Putamen, r | 14.14 | 0.5 | 19.99 | 0.5 | 5.85 |
| Pallidum, l | 20.75 | 0.6 | 29.38 | 0.5 | 8.63 |
| Pallidum, r | 16.57 | 0.5 | 22.8 | 0.5 | 6.23 |
| Thalamus, l | 14.81 | 0.6 | 22.8 | 0.5 | 7.99 |
| Thalamus, r | 12.37 | 0.5 | 19.1 | 0.5 | 6.73 |
| Hippocampus, l | 1.6 | 0.1 | 3.26 | 0.2 | 1.66 |
| Hippocampus, r | 6.83 | 0.4 | 13.01 | 0.5 | 6.18 |
| Amygdala, l | 6.03 | 0.2 | 13.08 | 0.2 | 7.05 |
| Amygdala, r | 8.44 | 0.4 | 19.09 | 0.5 | 10.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuster, S.; Beyer, L.; Palleis, C.; Harris, S.; Schmitt, J.; Weidinger, E.; Prix, C.; Bötzel, K.; Danek, A.; Rauchmann, B.-S.; et al. Impact of Partial Volume Correction on [18F]GE-180 PET Quantification in Subcortical Brain Regions of Patients with Corticobasal Syndrome. Brain Sci. 2022, 12, 204. https://doi.org/10.3390/brainsci12020204
Schuster S, Beyer L, Palleis C, Harris S, Schmitt J, Weidinger E, Prix C, Bötzel K, Danek A, Rauchmann B-S, et al. Impact of Partial Volume Correction on [18F]GE-180 PET Quantification in Subcortical Brain Regions of Patients with Corticobasal Syndrome. Brain Sciences. 2022; 12(2):204. https://doi.org/10.3390/brainsci12020204
Chicago/Turabian StyleSchuster, Sebastian, Leonie Beyer, Carla Palleis, Stefanie Harris, Julia Schmitt, Endy Weidinger, Catharina Prix, Kai Bötzel, Adrian Danek, Boris-Stephan Rauchmann, and et al. 2022. "Impact of Partial Volume Correction on [18F]GE-180 PET Quantification in Subcortical Brain Regions of Patients with Corticobasal Syndrome" Brain Sciences 12, no. 2: 204. https://doi.org/10.3390/brainsci12020204
APA StyleSchuster, S., Beyer, L., Palleis, C., Harris, S., Schmitt, J., Weidinger, E., Prix, C., Bötzel, K., Danek, A., Rauchmann, B.-S., Stöcklein, S., Lindner, S., Unterrainer, M., Albert, N. L., Mittlmeier, L. M., Wetzel, C., Rupprecht, R., Rominger, A., Bartenstein, P., ... Dekorsy, F. J. (2022). Impact of Partial Volume Correction on [18F]GE-180 PET Quantification in Subcortical Brain Regions of Patients with Corticobasal Syndrome. Brain Sciences, 12(2), 204. https://doi.org/10.3390/brainsci12020204