
A Breakdown of Immune Tolerance in the Cerebellum



Abstract
:1. Immune Surveillance of the Central Nervous System (CNS)
1.1. Blood–CSF Barrier: Portal for Immune Surveillance during Physiological Conditions
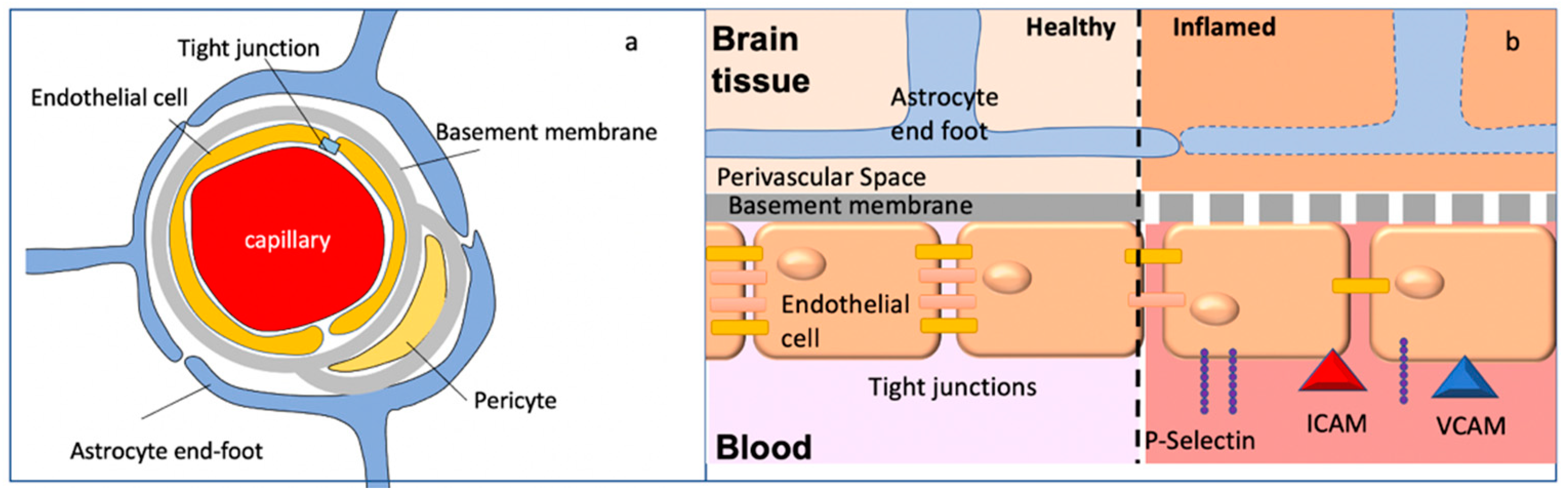
1.2. The Blood–Brain Barrier in Health and Disease
1.2.1. Inflammation and Breakdown of BBB Integrity
1.2.2. Region-Specific BBB Permeability
2. Autoimmunity Specific to the Cerebellum
2.1. Innate Immunity
2.2. Adaptive Immune Response
3. Diversity in Cerebellar Autoimmunity
3.1. Postinfectious Cerebellitis (PIC)
3.2. Paraneoplastic Cerebellar Degeneration (PCD)
3.2.1. Anti-Yo Autoantibody-Associated PCD
3.2.2. VGCC-Associated PCD
3.3. Gluten Ataxia (GA)
3.4. GAD65Ab-Associated Cerebellar Ataxia
3.5. mGluR1-Associated Cerebellar Ataxia
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bechmann, I.; Galea, I.; Perry, V.H. What is the blood-brain barrier (not)? Trends Immunol. 2007, 28, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ousman, S.S.; Kubes, P. Immune surveillance in the central nervous system. Nat. Neurosci. 2012, 15, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, B.; Vajkoczy, P.; Weller, R.O. The movers and shapers in immune privilege of the CNS. Nat. Immunol. 2017, 18, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Garden, G.A.; Möller, T. Microglia biology in health and disease. J. Neuroimmune Pharmacol. 2006, 1, 127–137. [Google Scholar] [CrossRef]
- Streit, W.J.; Mrak, R.E.; Griffin, W.S.T. Microglia and neuroinflammation: A pathological perspective. J. Neuroinflamm. 2004, 1, 14. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, B.; Ransohoff, R.M. The ins and outs of T-lymphocyte trafficking to the CNS: Anatomical sites and molecular mechanisms. Trends Immunol. 2005, 26, 485–495. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Kivisäkk, P.; Kidd, G. Three or more routes for leukocyte migration into the central nervous system. Nat. Rev. Immunol. 2003, 3, 569–581. [Google Scholar] [CrossRef]
- Ayub, M.; Jin, H.; Bae, J. The blood cerebrospinal fluid barrier orchestrates immunosurveillance, immunoprotection, and immunopathology in the central nervous system. BMB Rep. 2021, 54, 196–202. [Google Scholar] [CrossRef]
- Engelhardt, B.; Ransohoff, R.M. Capture, crawl, cross: The T cell code to breach the blood-brain barriers. Trends Immunol. 2012, 33, 579–589. [Google Scholar] [CrossRef]
- Meeker, R.B.; Williams, K.; Killebrew, D.A.; Hudson, L.C. Cell trafficking through the choroid plexus. Cell Adh. Migr. 2012, 6, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Kivisäkk, P.; Mahad, D.; Callahan, M.; Trebst, C.; Tucky, B.; Wei, T.; Wu, L.; Baekkevold, E.; Lassmann, H.; Staugaitis, S.; et al. Human cerebrospinal fluid central memory CD4+ T cells: Evidence for trafficking through choroid plexus and meninges via P-selectin. Proc. Natl. Acad. Sci. USA 2003, 100, 8389–8394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrithers, M.D.; Visintin, I.; Kang, S.J.; Janeway, C.A. Differential adhesion molecule requirements for immune surveillance and inflammatory recruitment. Brain 2000, 123, 1092–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumbholz, M.; Theil, D.; Steinmeyer, F.; Cepok, S.; Hemmer, B.; Hofbauer, M.; Farina, C.; Derfuss, T.; Junker, A.; Arzberger, T.; et al. CCL19 is constitutively expressed in the CNS, up-regulated in neuroinflammation, active and also inactive multiple sclerosis lesions. J. Neuroimmunol. 2007, 190, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Reboldi, A.; Coisne, C.; Baumjohann, D.; Benvenuto, F.; Bottinelli, D.; Lira, S.; Uccelli, A.; Lanzavecchia, A.; Engelhardt, B.; Sallusto, F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat. Immunol. 2009, 10, 514–523. [Google Scholar] [CrossRef]
- Johanson, C.; Stopa, E.; McMillan, P. The Blood–Cerebrospinal Fluid Barrier: Structure and Functional Significance. Methods Mol. Biol. 2011, 686, 101–131. [Google Scholar] [CrossRef]
- Wolburg, K.; Gerhardt, H.; Schulz, M.; Wolburg, H.; Engelhardt, B. Ultrastructural localization of adhesion molecules in the healthy and inflamed choroid plexus of the mouse. Cell Tissue Res. 1999, 296, 259–269. [Google Scholar] [CrossRef]
- Steffen, B.J.; Breier, G.; Butcher, E.C.; Schulz, M.; Engelhardt, B. ICAM-1, VCAM-1, and MAdCAM-1 are expressed on choroid plexus epithelium but not endothelium and mediate binding of lymphocytes in vitro. Am. J. Pathol. 1996, 148, 1819–1838. [Google Scholar]
- Erickson, M.A.; Banks, W.A. Neuroimmune axes of the blood-brain barriers and blood-brain interfaces: Bases for physiological regulation, disease states, and pharmacological interventions. Pharmacol. Rev. 2018, 70, 278–314. [Google Scholar] [CrossRef]
- De Graaf, M.T.; Sillevis Smitt, P.A.E.; Luitwieler, R.L.; Van Velzen, C.; Van Den Broek, P.D.M.; Kraan, J.; Gratama, J.W. Central memory CD4+ T cells dominate the normal cerebrospinal fluid. Cytom. Part B Clin. Cytom. 2011, 80B, 43–50. [Google Scholar] [CrossRef]
- Raper, D.; Louveau, A.; Kipnis, J. How do meningeal lymphatic vessels drain the CNS? Trends Neurosci 2016, 36, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Rustenhoven, J.; Drieu, A.; Mamuladze, T.; de Lima, K.; Dystra, T.; Wall, M.; Papadopoulos, Z.; Kanamori, M.; Salvador, A.; Baker, W.; et al. Functional characterization of the dural sinuses as a neuroimmune interface. Cell 2021, 184, 1000–1016. [Google Scholar] [CrossRef] [PubMed]
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Sherin, J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Kevin, L.; et al. Structural and functional features of central nervous system lymphatics. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Engelhardt, B. The anatomical and cellular basis of immune surveillance in the central nervous system. Nat. Rev. Immunol. 2012, 12, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef]
- Ren, H.; Han, R.; Chen, X.; Liu, X.; Wan, J.; Wang, L.; Yang, X.; Wang, J. Potential therapeutic targets for intracerebral hemorrhage-associated inflammation: An update. J. Cereb. Blood Flow Metab. 2020, 40, 1752–1768. [Google Scholar] [CrossRef] [PubMed]
- Allison, D.J.; Ditor, D.S. Immune dysfunction and chronic inflammation following spinal cord injury. Spinal Cord 2015, 53, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Garden, G.A. Epigenetics and the Modulation of Neuroinflammation. Neurotherapeutics 2013, 10, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Banks, W.A. The Blood-brain Barrier in Neuroimmunology: Tales of Separation and Assimilation. Brain Behav. Immun. 2015, 44, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Liu, B. Resveratrol attenuates lipopolysaccharide-induced dysfunction of blood-brain barrier in endothelial cells via AMPK activation. Korean J. Physiol. Pharmacol. 2016, 20, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.L.; Kittel, Á.; Veszelka, S.; Palmela, I.; Tóth, A.; Brites, D.; Deli, M.A.; Brito, M.A. Exposure to lipopolysaccharide and/or unconjugated bilirubin impair the integrity and function of brain microvascular endothelial cells. PLoS ONE 2012, 7, e35919. [Google Scholar] [CrossRef]
- Biesmans, S.; Meert, T.; Bouwknecht, J.; Acton, P.; Davoodi, N.; De Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.; Ver Donck, L.; et al. Systemic Immune Activation Leads to Neuroinflammation and Sickness Behavior in Mice. Mediat. Inflamm. 2013, 2013, 271359. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M. Astrocyte barriers to neurotoxic inflammation Michael. Nat. Rev. Neurosci. 2015, 16, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Liebner, S.; Dijkhuizen, R.M.; Reiss, Y.; Plate, K.H.; Agalliu, D.; Constantin, G. Functional morphology of the blood–brain barrier in health and disease. Acta Neuropathol. 2018, 135, 311–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelly, D.T.; Hennessy, E.; Dansereau, M.A.; Cunningham, C. A Systematic Analysis of the Peripheral and CNS Effects of Systemic LPS, IL-1Β, TNF-α and IL-6 Challenges in C57BL/6 Mice. PLoS ONE 2013, 8, e69123. [Google Scholar] [CrossRef]
- Chui, R.; Dorovini-Zis, K. Regulation of CCL2 and CCL3 expression in human brain endothelial cells by cytokines and lipopolysaccharide. J. Neuroinflamm. 2010, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, J.B. The adhesion molecule ICAM-1 and its regulation in relation with the blood-brain barrier. J. Neuroimmunol. 2002, 128, 58–68. [Google Scholar] [CrossRef]
- Horng, S.; Therattil, A.; Moyon, S.; Gordon, A.; Kim, K.; Argaw, A.T.; Hara, Y.; Mariani, J.N.; Sawai, S.; Flodby, P.; et al. Astrocytic tight junctions control inflammatory CNS lesion pathogenesis. J. Clin. Investig. 2017, 127, 3136–3151. [Google Scholar] [CrossRef]
- Welcome, M.O.; Mastorakis, N.E. Stress-induced blood brain barrier disruption: Molecular mechanisms and signaling pathways. Pharmacol. Res. 2020, 157, 104769. [Google Scholar] [CrossRef]
- Galea, I. The blood–brain barrier in systemic infection and inflammation. Cell. Mol. Immunol. 2021, 18, 2489–2501. [Google Scholar] [CrossRef]
- Li, W.; Cao, F.; Takase, H.; Arai, K.; Lo, E.H.; Lok, J. Blood–Brain Barrier Mechanisms in Stroke and Trauma. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Cserr, H.F.; Knopf, P.M. Cervical lymphatics, the blood-brain barrier and the immunoreactivity of the brain: A new view. Immunol. Today 1992, 13, 507–512. [Google Scholar] [CrossRef]
- Knopf, P.M.; Harling-Berg, C.J.; Cserr, H.F.; Basu, D.; Sirulnick, E.J.; Nolan, S.C.; Park, J.T.; Keir, G.; Thompson, E.J.; Hickey, W.F. Antigen-dependent intrathecal antibody synthesis in the normal rat brain: Tissue entry and local retention of antigen-specific B cells. J. Immunol. 1998, 161, 692–701. [Google Scholar] [PubMed]
- Ruano-Salguero, J.S.; Lee, K.H. Antibody transcytosis across brain endothelial-like cells occurs nonspecifically and independent of FcRn. Sci. Rep. 2020, 10, 3685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villabona-Rueda, A.; Erice, C.; Pardo, C.A.; Stins, M.F. The Evolving Concept of the Blood Brain Barrier (BBB): From a Single Static Barrier to a Heterogeneous and Dynamic Relay Center. Front. Cell. Neurosci. 2019, 13, 405. [Google Scholar] [CrossRef]
- Batiuk, M.Y.; Martirosyan, A.; Voet, T.; Ponting, C.P.; Belgard, T.G.; Holt, M.G. Molecularly distinct astrocyte subpopulations spatially pattern the adult mouse brain. bioRxiv 2018. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K.; et al. Pericytes regulate the blood-brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, I.; Nyúl-Tóth, Á.; Suciu, M.; Hermenean, A.; Krizbai, I.A. Heterogeneity of the blood-brain barrier. Tissue Barriers 2016, 4, e1143544. [Google Scholar] [CrossRef] [PubMed]
- Phares, T.W.; Kean, R.B.; Mikheeva, T.; Hooper, D.C. Regional Differences in Blood-Brain Barrier Permeability Changes and Inflammation in the Apathogenic Clearance of Virus from the Central Nervous System. J. Immunol. 2006, 176, 7666–7675. [Google Scholar] [CrossRef]
- Fabis, M.J.; Phares, T.W.; Kean, R.B.; Koprowski, H.; Hooper, D.C. Blood-brain barrier changes and cell invasion differ between therapeutic immune clearance of neurotrophic virus and CNS autoimmunity. Proc. Natl. Acad. Sci. USA 2008, 105, 15511–15516. [Google Scholar] [CrossRef] [Green Version]
- Silwedel, C.; Förster, C. Differential susceptibility of cerebral and cerebellar murine brain microvascular endothelial cells to loss of barrier properties in response to inflammatory stimuli. J. Neuroimmunol. 2006, 179, 37–45. [Google Scholar] [CrossRef]
- Garza, M.; Piquet, A.L. Update in Autoimmune Movement Disorders: Newly Described Antigen Targets in Autoimmune and Paraneoplastic Cerebellar Ataxia. Front. Neurol. 2021, 12, e683048. [Google Scholar] [CrossRef] [PubMed]
- Jarius, S.; Wildemann, B. “Medusa-head ataxia”: The expanding spectrum of Purkinje cell antibodies in autoimmune cerebellar ataxia. Part 1: Anti-mGluR1, anti-Homer-3, anti-Sj/ITPR1 and anti-CARP VIII. J. Neuroinflamm. 2015, 12, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loehrer, P.A.; Zieger, L.; Simon, O.J. Update on paraneoplastic cerebellar degeneration. Brain Sci. 2021, 11, 1414. [Google Scholar] [CrossRef]
- Mitoma, H.; Manto, M.; Hampe, C.S. Immune-mediated Cerebellar Ataxias: Practical Guidelines and Therapeutic Challenges. Curr. Neuropharmacol. 2018, 17, 33–58. [Google Scholar] [CrossRef] [PubMed]
- Staszewski, O.; Hagemeyer, N. Unique microglia expression profile in developing white matter. BMC Res. Notes 2019, 12, 367. [Google Scholar] [CrossRef]
- Hunter, C. A Rare Disease in Two Brothers. Proc. R. Soc. Med. 1917, 10, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Grabert, K.; Michoel, T.; Karavolos, M.H.; Clohisey, S.; Kenneth Baillie, J.; Stevens, M.P.; Freeman, T.C.; Summers, K.M.; McColl, B.W. Microglial brain region-dependent diversity and selective regional sensitivities to aging. Nat. Neurosci. 2016, 19, 504–516. [Google Scholar] [CrossRef] [Green Version]
- Ferro, A.; Sheeler, C.; Rosa, J.G.; Cvetanovic, M. Role of Microglia in Ataxias. J. Mol. Biol. 2019, 431, 1792–1804. [Google Scholar] [CrossRef]
- Aikawa, T.; Mogushi, K.; Iijima-Tsutsui, K.; Ishikawa, K.; Sakurai, M.; Tanaka, H.; Mizusawa, H.; Watase, K. Loss of MyD88 alters neuroinflammatory response and attenuates early Purkinje cell loss in a spinocerebellar ataxia type 6 mouse model. Hum. Mol. Genet. 2015, 24, 4780–4791. [Google Scholar] [CrossRef]
- Cvetanovic, M. Decreased Expression of Glutamate Transporter GLAST in Bergmann Glia Is Associated with the Loss of Purkinje Neurons in the Spinocerebellar Ataxia Type 1. Cerebellum 2015, 14, 8–11. [Google Scholar] [CrossRef]
- Shen, Y.; McMackin, M.Z.; Shan, Y.; Raetz, A.; David, S.; Cortopassi, G. Frataxin deficiency promotes excess microglial DNA damage and inflammation that is rescued by PJ34. PLoS ONE 2016, 11, e0151026. [Google Scholar] [CrossRef] [PubMed]
- Stoessel, M.B.; Majewska, A.K. Little cells of the little brain: Microglia in cerebellar development and function. Trends Neurosci. 2021, 44, 564–578. [Google Scholar] [CrossRef]
- Bellamy, T. Interactions between Purkinje neurones and Bergmann glia. Cerebellum 2006, 5, 116–126. [Google Scholar] [CrossRef]
- Bergles, D.E.; Dzubay, J.A.; Jahr, C.E. Glutamate transporter currents in Bergmann glial cells follow the time course of extrasynaptic glutamate. Proc. Natl. Acad. Sci. USA 1997, 94, 14821–14825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuvaev, A.N.; Belozor, O.S.; Mozhei, O.; Yakovleva, D.A.; Potapenko, I.V.; Shuvaev, A.N.; Smolnikova, M.V.; Salmin, V.V.; Salmina, A.B.; Hirai, H.; et al. Chronic optogenetic stimulation of Bergman glia leads to dysfunction of EAAT1 and Purkinje cell death, mimicking the events caused by expression of pathogenic ataxin-1. Neurobiol. Dis. 2021, 154, 105340. [Google Scholar] [CrossRef] [PubMed]
- Hadjivassiliou, M.; Graus, F.; Honnorat, J.; Jarius, S.; Titulaer, M.; Manto, M.; Hoggard, N.; Sarrigiannis, P.; Mitoma, H. Diagnostic Criteria for Primary Autoimmune Cerebellar Ataxia—Guidelines from an International Task Force on Immune-Mediated Cerebellar Ataxias. Cerebellum 2020, 19, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Mitoma, H.; Manto, M.; Hadjivassiliou, M. Immune-mediated cerebellar ataxias: Clinical diagnosis and treatment based on immunological and physiological mechanisms. J. Mov. Disord. 2021, 14, 10–28. [Google Scholar] [CrossRef]
- Aw, D.; Silva, A.B.; Palmer, D.B. Immunosenescence: Emerging challenges for an ageing population. Immunology 2007, 120, 435–446. [Google Scholar] [CrossRef]
- Shaw, A.C.; Goldstein, D.R.; Montgomery, R.R. Age-dependent dysregulation of innate immunity. Nat. Rev. Immunol. 2013, 13, 875–887. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Hussain, B.; Fang, C.; Chang, J. Blood–Brain Barrier Breakdown: An Emerging Biomarker of Cognitive Impairment in Normal Aging and Dementia. Front. Neurosci. 2021, 15, e688090. [Google Scholar] [CrossRef] [PubMed]
- Bors, L.; Tóth, K.; Tóth, E.Z.; Bajza, Á.; Csorba, A.; Szigeti, K.; Máthé, D.; Perlaki, G.; Orsi, G.; Tóth, G.K.; et al. Age-dependent changes at the blood-brain barrier. A Comparative structural and functional study in young adult and middle aged rats. Brain Res. Bull. 2018, 139, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-Brain barrier breakdown in the aging human hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Erickson, M.A.; Banks, W.A. Age-associated changes in the immune system and blood–brain barrier functions. Int. J. Mol. Sci. 2019, 20, 1632. [Google Scholar] [CrossRef] [Green Version]
- Palus, M.; Vancova, M.; Sirmarova, J.; Elsterova, J.; Perner, J.; Ruzek, D. Tick-borne encephalitis virus infects human brain microvascular endothelial cells without compromising blood-brain barrier integrity. Virology 2017, 507, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Burdo, T.H.; Weiffenbach, A.; Woods, S.P.; Letendre, S.; Ellis, R.J.; Williams, K.C. Elevated sCD163 in plasma but not cerebrospinal fluid is a marker of neurocognitive impairment in HIV infection. Aids 2013, 27, 1387–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menicucci, A.R.; Versteeg, K.; Woolsey, C.; Mire, C.E.; Geisbert, J.B.; Cross, R.W.; Agans, K.N.; Jankeel, A.; Geisbert, T.W.; Messaoudi, I. Transcriptome analysis of circulating immune cell subsets highlight the role of monocytes in Zaire Ebola Virus Makona pathogenesis. Front. Immunol. 2017, 8, e1372. [Google Scholar] [CrossRef] [Green Version]
- Chaves, A.J.; Vergara-Alert, J.; Busquets, N.; Valle, R.; Rivas, R.; Ramis, A.; Darji, A.; Majó, N. Neuroinvasion of the highly pathogenic influenza virus H7N1 is caused by disruption of the blood brain barrier in an avian model. PLoS ONE 2014, 9, e115138. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Racaniello, V.R. Poliovirus Spreads from Muscle to the Central Nervous System by Neural Pathways. J. Infect. Dis. 1992, 166, 747–752. [Google Scholar] [CrossRef]
- Muehlenbachs, A.; Bhatnagar, J.; Zaki, S.R. Tissue tropism, pathology and pathogenesis of enterovirus infection†. J. Pathol. 2015, 235, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Sips, G.J.; Wilschut, J.; Smit, J.M. Neuroinvasive flavivirus infections. Rev. Med. Virol. 2012, 22, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.; Diadori, P.; Schoenroth, L.; Fritzler, M. Autoantibodies in childhood post-varicella acute cerebellar ataxia. Can. J. Neurol. Sci. 2000, 27, 316–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzler, M.J.; Zhang, M.; Stinton, L.M.; Rattner, J.B. Spectrum of centrosome autoantibodies in childhood varicella and post-varicella acute cerebellar ataxia. BMC Pediatr. 2003, 3, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchibori, A.; Sakuta, M.; Kusunoki, S.; Chiba, A. Autoantibodies in postinfectious acute cerebellar ataxia. Neurology 2005, 65, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Cimolai, N.; Mah, D.; Roland, E. Anticentriolar autoantibodies in children with central nervous system manifestations of Mycoplasma pneumoniae infection. J. Neurol. Neurosurg. Psychiatry 1994, 57, 638–639. [Google Scholar] [CrossRef]
- Pruitt, A.A. Infections of the cerebellum. Neurol. Clin. 2014, 32, 1117–1131. [Google Scholar] [CrossRef]
- Desai, J.; Mitchell, W.G. Acute cerebellar ataxia, acute cerebellitis, and opsoclonus-myoclonus syndrome. J. Child. Neurol. 2012, 27, 1482–1488. [Google Scholar] [CrossRef]
- D’Ambrosio, E.; Khalighinejad, F.; Ionete, C. Intravenous immunoglobulins in an adult case of post-EBV cerebellitis. BMJ Case Rep. 2020, 13, 2019–2021. [Google Scholar] [CrossRef]
- Pillai, S.C.; Hacohen, Y.; Tantsis, E.; Prelog, K.; Merheb, V.; Kesson, A.; Barnes, E.; Gill, D.; Webster, R.; Menezes, M.; et al. Infectious and autoantibody-associated encephalitis: Clinical features and long-term outcome. Pediatrics 2015, 135, e974–e984. [Google Scholar] [CrossRef] [Green Version]
- D’Alonzo, R.; Mencaroni, E.; Di Genova, L.; Laino, D.; Principi, N.; Esposito, S. Pathogenesis and treatment of neurologic diseases associated with mycoplasma pneumoniae infection. Front. Microbiol. 2018, 9, 2751. [Google Scholar] [CrossRef]
- Schmucker, R.D.; Ehret, A.; Marshall, G.S. Cerebellitis and acute obstructive hydrocephalus associated with mycoplasma pneumoniae infection. Pediatr. Infect. Dis. J. 2014, 33, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Meyer Sauteur, P.M.; De Bruijn, A.C.J.M.; Graça, C.; Tio-Gillen, A.P.; Estevão, S.C.; Hoogenboezem, T.; Hendriks, R.W.; Berger, C.; Jacobs, B.C.; Van Rossum, A.M.C.; et al. Antibodies to protein but not glycolipid structures are important for host defense against mycoplasma pneumoniae. Infect. Immun. 2019, 87, e00663-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusunoki, S.; Shiina, M.; Kanazawa, I. Anti-Gal-C antibodies in GBS subsequent to mycoplasma infection: Evidence of molecular mimicry. Neurology 2001, 57, 736–738. [Google Scholar] [CrossRef] [PubMed]
- Koga, M.; Gilbert, M.; Li, J.; Koike, S.; Takahashi, M.; Furukawa, K.; Hirata, K.; Yuki, N. Antecedent infections in Fisher syndrome. Neurology 2005, 64, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Chiba, A.; Kusunoki, S.; Shimizu, T.; Kanazawa, I. Serum IgG antibody to ganglioside GQ1b is a possible marker of Miller Fisher syndrome. Ann. Neurol. 1992, 31, 677–679. [Google Scholar] [CrossRef]
- Willison, H.J.; Veitch, J.; Paterson, G.; Kennedy, P.G.E. Miller Fisher syndrome is associated with serum antibodies to GQ1b ganglioside. J. Neurol. Neurosurg. Psychiatry 1993, 56, 204–206. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, S.; Taboada, J. A Case of Miller Fisher Syndrome and Literature Review. Cureus 2017, 9, e1048. [Google Scholar] [CrossRef] [Green Version]
- Kimoto, K.; Koga, M.; Odaka, M.; Hirata, K.; Takahashi, M.; Li, J.; Gilbert, M.; Yuki, N. Relationship of bacterial strains to clinical syndromes of Campylobacter-associated neuropathies. Neurology 2006, 67, 1837–1843. [Google Scholar] [CrossRef]
- Houliston, R.S.; Koga, M.; Li, J.; Jarrell, H.C.; Richards, J.C.; Vitiazeva, V.; Schweda, E.K.H.; Yuki, N.; Gilbert, M. A Haemophilus influenzae strain associated with fisher syndrome expresses a novel disialylated ganglioside mimic. Biochemistry 2007, 46, 8164–8171. [Google Scholar] [CrossRef]
- Ito, M.; Kuwabara, S.; Odaka, M.; Misawa, S.; Koga, M.; Hirata, K.; Yuki, N. Bickerstaff’s brainstem encephalitis and Fisher syndrome form a continuous spectrum: Clinical analysis of 581 cases. J. Neurol. 2008, 255, 674–682. [Google Scholar] [CrossRef]
- Ropper, A.H.; Shahani, B. Proposed Mechanism of Ataxia in Fisher’s Syndrome. Arch. Neurol. 1983, 40, 537–538. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.X.; Willison, H.J.; Pedrosa-Domellöf, F. Immunolocalization of GQ1b and related gangliosides in human extraocular neuromuscular junctions and muscle spindles. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3226–3232. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Kuwabara, S.; Fukutake, T.; Yuki, N.; Hattori, T. Clinical features and prognosis of Miller Fisher syndrome. Neurology 2001, 56, 1104–1106. [Google Scholar] [CrossRef] [PubMed]
- Sarigecili, E.; Arslan, I.; Ucar, H.K.; Celik, U. Pediatric anti-NMDA receptor encephalitis associated with COVID-19. Child’s Nerv. Syst. 2021, 37, 3919–3922. [Google Scholar] [CrossRef]
- Gaughan, M.; Connolly, S.; O’Riordan, S.; Tubridy, N.; McGuigan, C.; Kinsella, J. Pediatric Parainfectious Encephalitis Associated With COVID-19. Neurology 2021, 96, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Oosthuizen, K.; Steyn, E.; Tucker, L.; Ncube, I.; Hardie, D.; Marais, S. SARS-CoV-2 Encephalitis Presenting as a Clinical Cerebellar Syndrome: A Case Report. Neurology 2021, 97, 27–29. [Google Scholar] [CrossRef]
- Ahsan, N.; Jafarpour, S.; Santoro, J.D. Myelin oligodendrocyte glycoprotein antibody encephalitis following severe acute respiratory syndrome coronavirus 2 in a pediatric patient. Clin. Exp. Pediatr. 2021, 64, 310–312. [Google Scholar] [CrossRef]
- Guilmot, A.; Maldonado Slootjes, S.; Sellimi, A.; Bronchain, M.; Hanseeuw, B.; Belkhir, L.; Yombi, J.C.; De Greef, J.; Pothen, L.; Yildiz, H.; et al. Immune-mediated neurological syndromes in SARS-CoV-2-infected patients. J. Neurol. 2021, 268, 751–757. [Google Scholar] [CrossRef]
- Connolly, A.M.; Dodson, W.E.; Prensky, A.L.; Rust, R.S. Course and outcome of acute cerebellar ataxia. Ann. Neurol. 1994, 35, 673–679. [Google Scholar] [CrossRef]
- Blumkin, L.; Pranzatelli, M.R. Acquired ataxias, infectious and para-infectious. Handb Clin. Neurol. 2012, 103, 137–146. [Google Scholar] [CrossRef]
- Schmahmann, J.D. Plasmapheresis improves outcome in postinfectious cerebellitis induced by Epstein-Barr virus. Neurology 2004, 62, 1443. [Google Scholar] [CrossRef] [PubMed]
- Giannantoni, N.M.; Rigamonti, E.; Rampolli, F.I.; Grazioli-Gauthier, L.; Allali, G.; Vanini, G. Myoclonus and Cerebellar Ataxia Associated with SARS-CoV-2 Infection: Case Report and Review of the Literature. Eur. J. Case Rep. Intern. Med. 2021, 8, e002531. [Google Scholar] [CrossRef]
- Chan, J.L.; Murphy, K.A.; Sarna, J.R. Myoclonus and cerebellar ataxia associated with COVID-19: A case report and systematic review. J. Neurol. 2021, 268, 3517–3548. [Google Scholar] [CrossRef] [PubMed]
- Peterson, K.; Rosenblum, M.; Kotanides, H.; Posner, J. Paraneoplastic cerebellar degeneration. Neurology 1992, 42, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Shams’Ili, S.; Grefkens, J.; De Leeuw, B.; Van den Bent, M.; Hooijkaas, H.; Van der Holt, B.; Vecht, C.; Sillevis Smitt, P. Paraneoplastic cerebellar degeneration associated with antineuronal antibodies: Analysis of 50 patients. Brain 2003, 126, 1409–1418. [Google Scholar] [CrossRef] [Green Version]
- Bruhnding, A.; Notch, D.; Beard, A. Anti-Yo positive paraneoplastic cerebellar degeneration in the setting of cholangiocarcinoma. J. Clin. Neurosci. 2017, 36, 71–72. [Google Scholar] [CrossRef]
- Mitoma, H.; Adhikari, K.; Aeschlimann, D.; Chattopadhyay, P.; Hadjivassiliou, M.; Hampe, C.S.; Honnorat, J.; Joubert, B.; Kakei, S.; Lee, J.; et al. Consensus Paper: Neuroimmune Mechanisms of Cerebellar Ataxias. Cerebellum 2016, 15, 213–232. [Google Scholar] [CrossRef] [Green Version]
- Vernino, S. Paraneoplastic cerebellar degeneration. Handb. Clin. Neurol. 2012, 103, 215–223. [Google Scholar] [CrossRef]
- Drlicek, M.; Bianchi, G.; Bogliun, G.; Casati, B.; Grisold, W.; Kolig, C.; Liszka-Setinek, U.; Marzorati, L.; Wondrusch, E.; Cavaletti, G. Antibodies of the anti-Yo and anti-Ri type in the absence of paraneoplastic neurological syndromes: A long-term survey of ovarian cancer patients. J. Neurol. 1997, 244, 85–89. [Google Scholar] [CrossRef]
- Eichler, T.W.; Totland, C.; Haugen, M.; Qvale, T.H.; Mazengia, K.; Storstein, A.; Haukanes, B.I.; Vedeler, C.A. CDR2L Antibodies: A New Player in Paraneoplastic Cerebellar Degeneration. PLoS ONE 2013, 8, e66002. [Google Scholar] [CrossRef]
- Mason, W.P.; Graus, F.; Lang, B.; Honnorat, J.; Delattre, J.Y.; Valldeoriola, F.; Antoine, J.C.; Rosenblum, M.K.; Rosenfeld, M.R.; Newsom-Davis, J.; et al. Small-cell lung cancer, paraneoplastic cerebellar degeneration and the Lambert-Eaton myasthenic syndrome. Brain 1997, 120, 1279–1300. [Google Scholar] [CrossRef]
- Rana, A.Q.; Rana, A.N.; Adlul, A. Acute ataxia due to anti-Yo antibody paraneoplastic cerebellar degeneration 4 months prior to diagnosis of uterine carcinoma. Acta Neurol. Belg. 2012, 112, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Hasadsri, L.; Lee, J.; Wang, B.H.; Yekkirala, L.; Wang, M. Anti-Yo Associated Paraneoplastic Cerebellar Degeneration in a Man with Large Cell Cancer of the Lung. Case Rep. Neurol. Med. 2013, 2013, 725936. [Google Scholar] [CrossRef] [PubMed]
- Kråkenes, T.; Herdlevær, I.; Raspotnig, M.; Haugen, M.; Schubert, M.; Vedeler, C.A. CDR2L Is the Major Yo Antibody Target in Paraneoplastic Cerebellar Degeneration. Ann. Neurol. 2019, 86, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Corradi, J.P.; Yang, C.; Darnell, J.C.; Dalmau, J.; Darnell, R.B. A post-transcriptional regulatory mechanism restricts expression of the paraneoplastic cerebellar degeneration antigen cdr2 to immune privileged tissues. J. Neurosci. 1997, 17, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.K.; Darnell, R.B. Neuroimmunology of the paraneoplastic neurological degenerations. Curr. Opin. Immunol. 2004, 16, 616–622. [Google Scholar] [CrossRef]
- Herdlevær, I.; Kråkenes, T.; Schubert, M.; Vedeler, C.A. Localization of CDR2L and CDR2 in paraneoplastic cerebellar degeneration. Ann. Clin. Transl. Neurol. 2020, 7, 2231–2242. [Google Scholar] [CrossRef]
- Raspotnig, M.; Haugen, M.; Thorsteinsdottir, M.; Stefansson, I.; Salvesen, H.B.; Storstein, A.; Vedeler, C.A. Cerebellar degeneration-related proteins 2 and 2-like are present in ovarian cancer in patients with and without Yo antibodies. Cancer Immunol. Immunother. 2017, 66, 1463–1471. [Google Scholar] [CrossRef]
- Diamond, B.; Huerta, P.; Mina-Osorio, P.; Kowal, C.; Volpe, B. Losing your nerves? Maybe it’s the antibodies. Nat. Rev. Immunol 2009, 9, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, E.L.; Vance, B.A.; Klein, R.S.; Voloschin, A.; Dalmau, J.; Vonderheide, R.H. Functional analysis of CD8+ T cell responses to the onconeural self protein cdr2 in patients with paraneoplastic cerebellar degeneration. J. Neuroimmunol. 2008, 193, 173–182. [Google Scholar] [CrossRef]
- Small, M.; Treilleux, I.; Couillault, C.; Pissaloux, D.; Picard, G.; Paindavoine, S.; Attignon, V.; Wang, Q.; Rogemond, V.; Lay, S.; et al. Genetic alterations and tumor immune attack in Yo paraneoplastic cerebellar degeneration. Acta Neuropathol. 2018, 135, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Dalmau, J.; Rosenfeld, M.R. Paraneoplastic syndromes of the CNS. Lancet Neurol. 2008, 7, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Verschnuren, J.; Chuang, L.; Rosenblum, M.K.; Lieberman, F.; Pryor, A.; Posner, J.B.; Dalmau, J. Inflammatory infiltrates and complete absence of Purkinje cells in anti-Yo-associated paraneoplastic cerebellar degeneration. Acta Neuropathol. 1996, 91, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.L.; Darnell, J.C.; Bender, A.; Francisco, L.M.; Bhardwaj, N.; Darnell, R.B. Tumor-specific killer cells in paraneoplastic cerebellar degeneration. Nat. Med. 1998, 4, 1321–1324. [Google Scholar] [CrossRef] [Green Version]
- Greenlee, J.E.; Clawson, S.A.; Hill, K.E.; Wood, B.L.; Tsunoda, I.; Carlson, N.G. Purkinje cell death after uptake of anti-yo antibodies in cerebellar slice cultures. J. Neuropathol. Exp. Neurol. 2010, 69, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Graus, F.; Illa, I.; Agusti, M.; Ribalta, T.; Cruz-Sanchez, F.; Juarez, C. Effect of intraventricular injection of an anti-Purkinje cell antibody (anti-Yo) in a guinea pig model. J. Neurol. Sci. 1991, 106, 82–87. [Google Scholar] [CrossRef]
- Greenlee, J.E.; Burns, J.B.; Rose, J.W.; Jaeckle, K.A.; Clawson, S. Uptake of systemically administered human anticerebellar antibody by rat Purkinje cells following blood-brain barrier disruption. Acta Neuropathol. 1995, 89, 341–345. [Google Scholar] [CrossRef]
- Hill, K.E.; Clawson, S.A.; Rose, J.W.; Carlson, N.G.; Greenlee, J.E. Cerebellar Purkinje cells incorporate immunoglobulins and immunotoxins in vitro: Implications for human neurological disease and immunotherapeutics. J. Neuroinflamm. 2009, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Manto, M.; Honnorat, J.; Hampe, C.S.; Guerra-Narbona, R.; López-Ramos, J.C.; Delgado-García, J.M.; Saitow, F.; Suzuki, H.; Yanagawa, Y.; Mizusawa, H.; et al. Disease-specific monoclonal antibodies targeting glutamate decarboxylase impair GABAergic neurotransmission and affect motor learning and behavioral functions. Front. Behav. Neurosci. 2015, 9, 78. [Google Scholar] [CrossRef] [Green Version]
- Jarius, S.; Wildemann, B. “Medusa head ataxia”: The expanding spectrum of Purkinje cell antibodies in autoimmune cerebellar ataxia. Part 2: Anti-PKC-gamma, anti-GluR-delta2, anti-Ca/ARHGAP26 and anti-VGCC. J. Neuroinflamm. 2015, 12, 167. [Google Scholar] [CrossRef] [Green Version]
- Jarius, S.; Wildemann, B. “Medusa head ataxia”: The expanding spectrum of Purkinje cell antibodies in autoimmune cerebellar ataxia. Part 3: Anti-Yo/CDR2, anti-Nb/AP3B2, PCA-2, anti-Tr/DNER, other antibodies, diagnostic pitfalls, summary and outlook. J. Neuroinflamm. 2015, 12, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panja, D.; Vedeler, C.A.; Schubert, M. Paraneoplastic cerebellar degeneration: Yo antibody alters mitochondrial calcium buffering capacity. Neuropathol. Appl. Neurobiol. 2019, 45, 141–156. [Google Scholar] [CrossRef]
- Schubert, M.; Panja, D.; Haugen, M.; Bramham, C.R.; Vedeler, C.A. Paraneoplastic CDR2 and CDR2L antibodies affect Purkinje cell calcium homeostasis. Acta Neuropathol. 2014, 128, 835–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laure-Kamionowska, M.; Málińska, D. Calbindin positive Purkinje cells in the pathology of human cerebellum occurring at the time of its development. Folia Neuropathol. 2009, 47, 300–305. [Google Scholar] [PubMed]
- Hwang, J.Y.; Lee, J.; Oh, C.K.; Kang, H.W.; Hwang, I.Y.; Um, J.W.; Park, H.C.; Kim, S.; Shin, J.H.; Park, W.Y.; et al. Proteolytic degradation and potential role of onconeural protein cdr2 in neurodegeneration. Cell Death Dis. 2016, 7, e2240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palop, J.J.; Jones, B.; Kekonius, L.; Chin, J.; Yu, G.Q.; Raber, J.; Masliah, E.; Mucke, L. Neuronal depletion of calcium-dependent proteins in the dentate gyrus is tightly linked to Alzheimer’s disease-related cognitive deficits. Proc. Natl. Acad. Sci. USA 2003, 100, 9572–9577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, H.J.; Park, W.Y.; Corradi, J.P.; Darnell, R.B. The cytoplasmic Purkinje onconeural antigen cdr2 down-regulates c-Myc function: Implications for neuronal and tumor cell survival. Genes Dev. 1999, 13, 2087–2097. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Tanaka, K.; Onodera, O.; Tsuji, S. Trial to establish an animal model of paraneoplastic cerebellar degeneration with anti-Yo antibody. 1. Mouse strains bearing different MHC molecules produce antibodies on immunization with recombinant Yo protein, but do not cause Purkinje cell loss. Clin. Neurol. Neurosurg. 1995, 97, 95–100. [Google Scholar] [CrossRef]
- Tanaka, M.; Tanaka, K.; Tsuji, S.; Kawata, A.; Kojima, S.; Kurokawa, T.; Kira, J.; Takiguchi, M. Cytotoxic T cell activity against the peptide, AYRARALEL, from Yo protein of patients with the HLA A24 or B27 supertype and paraneoplastic cerebellar degeneration. J. Neurol. Sci. 2001, 188, 1–65. [Google Scholar] [CrossRef]
- Sutton, I.J.; Steele, J.; Savage, C.O.; Winer, J.B.; Young, L.S. An interferon-γ ELISPOT and immunohistochemical investigation of cytotoxic T lymphocyte-mediated tumour immunity in patients with paraneoplastic cerebellar degeneration and anti-Yo antibodies. J. Neuroimmunol. 2004, 150, 98–106. [Google Scholar] [CrossRef]
- Albert, M.; Austin, L.; Danress, R.B. Detection and treatment of activated T cells in the cerebrospinal fluid of patients with paraneoplastic cerebellar degeneration. Ann. Neurol. 2000, 47, 9–17. [Google Scholar] [CrossRef]
- Aye, M.M.; Kasai, T.; Tashiro, Y.; Xing, H.Q.; Shirahama, H.; Mitsuda, M.; Suetsugu, T.; Tanaka, K.; Osame, M.; Izumo, S. CD8 positive T-cell infiltration in the dentate nucleus of paraneoplastic cerebellar degeneration. J. Neuroimmunol. 2009, 208, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Stich, O.; Jarius, S.; Kleer, B.; Rasiah, C.; Voltz, R.; Rauer, S. Specific antibody index in cerebrospinal fluid from patients with central and peripheral paraneoplastic neurological syndromes. J. Neuroimmunol. 2007, 183, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Stich, O.; Graus, F.; Rasiah, C.; Rauer, S. Qualitative evidence of anti-Yo-specific intrathecal antibody synthesis in patients with paraneoplastic cerebellar degeneration. J. Neuroimmunol. 2003, 141, 165–169. [Google Scholar] [CrossRef]
- Lennon, V.A.; Kryzer, T.J.; Griesmann, G.E.; O’Suilleabhain, P.E.; Windebank, A.J.; Woppmann, A.; Miljanich, G.P.; Lambert, E.H. Calcium-Channel Antibodies in the Lambert–Eaton Syndrome and Other Paraneoplastic Syndromes. N. Engl. J. Med. 1995, 332, 1467–1475. [Google Scholar] [CrossRef]
- Graus, F.; Lang, B.; Pozo-Rosich, P.; Saiz, A.; Casamitjana, R.; Vincent, A. P/Q type calcium- channel antibodies in paraneoplastic cerebellar degeneration with lung cancer. Neurology 2002, 59, 764–766. [Google Scholar] [CrossRef]
- Fukuda, T.; Motomura, M.; Nakao, Y.; Shiraishi, H.; Yoshimura, T.; Iwanaga, K.; Tsujihata, M.; Eguchi, K. Reduction of P/Q-type calcium channels in the postmortem cerebellum of paraneoplastic cerebellar degeneration with Lambert-Eaton myasthenic syndrome. Ann. Neurol. 2003, 53, 21–28. [Google Scholar] [CrossRef]
- Bürk, K.; Wick, M.; Roth, G.; Decker, P.; Voltz, R. Antineuronal antibodies in sporadic late-onset cerebellar ataxia. J. Neurol. 2010, 257, 59–62. [Google Scholar] [CrossRef]
- Liao, Y.J.; Safa, P.; Chen, Y.R.; Sobel, R.A.; Boyden, E.S.; Tsien, R.W. Anti-Ca2+ channel antibody attenuates Ca2+ currents and mimics cerebellar ataxia in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 2705–2710. [Google Scholar] [CrossRef] [Green Version]
- Martín-García, E.; Mannara, F.; Gutiérrez-Cuesta, J.; Sabater, L.; Dalmau, J.; Maldonado, R.; Graus, F. Intrathecal injection of P/Q type voltage-gated calcium channel antibodies from paraneoplastic cerebellar degeneration cause ataxia in mice. J. Neuroimmunol. 2013, 261, 53–59. [Google Scholar] [CrossRef] [Green Version]
- McKasson, M.; Clardy, S.L.; Clawson, S.A.; Hill, K.E.; Wood, B.; Carlson, N.; Bromberg, M.; Greenlee, J.E. Voltage-gated calcium channel autoimmune cerebellar degeneration. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3, e222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellkofer, H.L.; Voltz, R.; Kuempfel, T. Favorable response to rituximab in a patient with anti-VGCC-positive lambert-eaton myasthenic syndrome and cerebellar dysfunction. Muscle Nerve 2009, 40, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Rigamonti, A.; Lauria, G.; Stanzani, L.; Mantero, V.; Andreetta, F.; Salmaggi, A. Non-paraneoplastic voltage-gated calcium channels antibody-mediated cerebellar ataxia responsive to IVIG treatment. J. Neurol. Sci. 2014, 336, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, H.; Manto, M.; Hampe, C.S. Immune-mediated cerebellar ataxias: From bench to bedside. Cerebellum Ataxias 2017, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancaster, E.; Dalmau, J. Neuronal autoantigens-pathogenesis, associated disorders and antibody testing. Nat. Rev. Neurol. 2012, 8, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Ma, Y.; Zekeridou, A.; Lennon, V.A. Case Report: Innate Immune System Challenge Unleashes Paraneoplastic Neurological Autoimmunity. Front. Neurol. 2020, 11, 598894. [Google Scholar] [CrossRef]
- Bürk, K.; Farecki, M.L.; Lamprecht, G.; Roth, G.; Decker, P.; Weller, M.; Rammensee, H.G.; Oertel, W. Neurological symptoms in patients with biopsy proven celiac disease. Mov. Disord. 2009, 24, 2358–2362. [Google Scholar] [CrossRef] [Green Version]
- Hadjivassiliou, M.; Grünewald, R.A.; Chattopadhyay, A.K.; Davies-Jones, G.A.B.; Gibson, A.; Jarratt, J.A.; Kandler, R.H.; Lobo, A.; Powell, T.; Smith, C.M.L. Clinical, radiological, neurophysiological, and neuropathological characteristics of gluten ataxia. Lancet 1998, 352, 1582–1585. [Google Scholar] [CrossRef]
- Mearns, E.S.; Taylor, A.; Thomas Craig, K.J.; Puglielli, S.; Cichewicz, A.B.; Leffler, D.A.; Sanders, D.S.; Lebwohl, B.; Hadjivassiliou, M. Neurological manifestations of neuropathy and ataxia in celiac disease: A systematic review. Nutrients 2019, 11, 380. [Google Scholar] [CrossRef] [Green Version]
- Hadjivassiliou, M.; Rao, D.G.; Grìnewald, R.A.; Aeschlimann, D.P.; Sarrigiannis, P.G.; Hoggard, N.; Aeschlimann, P.; Mooney, P.D.; Sanders, D.S. Neurological dysfunction in coeliac disease and non-coeliac gluten sensitivity. Am. J. Gastroenterol. 2016, 111, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Hadjivassiliou, M.; Williamson, C.A.; Woodroofe, N. The immunology of gluten sensitivity: Beyond the gut. Trends Immunol. 2004, 25, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Rouvroye, M.D.; Zis, P.; Van Dam, A.M.; Rozemuller, A.J.M.; Bouma, G.; Hadjivassiliou, M. The neuropathology of gluten-related neurological disorders: A systematic review. Nutrients 2020, 12, 822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittelbronn, M.; Schittenhelm, J.; Bakos, G.; De Vos, R.A.; Wehrmann, M.; Meyermann, R.; Bürk, K. CD8+/perforin+/granzyme B+ effector cells infiltrating cerebellum and inferior olives in gluten ataxia. Neuropathology 2010, 30, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Zis, P.; Rao, D.G.; Sarrigiannis, P.G.; Aeschlimann, P.; Aeschlimann, D.P.; Sanders, D.; Grünewald, R.A.; Hadjivassiliou, M. Transglutaminase 6 antibodies in gluten neuropathy. Dig. Liver Dis. 2017, 49, 1196–1200. [Google Scholar] [CrossRef]
- Lin, C.; Wang, M.; Tse, W.; Pinotti, R.; Alaedini, A.; Green, P.; Kuo, S. Serum antigliadin antibodies in cerebellar ataxias: A systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2018, 89, 1174–1180. [Google Scholar] [CrossRef]
- Hadjivassiliou, M.; Sanders, D.S.; Grünewald, R.A.; Woodroofe, N.; Boscolo, S.; Aeschlimann, D. Gluten sensitivity: From gut to brain. Lancet Neurol. 2010, 9, 318–330. [Google Scholar] [CrossRef]
- Liu, Y.T.; Tang, B.S.; Lan, W.; Song, N.N.; Huang, Y.; Zhang, L.; Guan, W.J.; Shi, Y.T.; Shen, L.; Jiang, H.; et al. Distribution of transglutaminase 6 in the central nervous system of adult mice. Anat. Rec. 2013, 296, 1576–1587. [Google Scholar] [CrossRef]
- Hadjivassiliou, M.; Aeschlimann, P.; Strigun, A.; Sanders, D.S.; Woodroofe, N.; Aeschlimann, D. Autoantibodies in gluten ataxia recognize a novel neuronal transglutaminase. Ann. Neurol. 2008, 64, 332–343. [Google Scholar] [CrossRef]
- Boscolo, S.; Lorenzon, A.; Sblattero, D.; Florian, F.; Stebel, M.; Marzari, R.; Not, T.; Aeschlimann, D.; Ventura, A.; Hadjivassiliou, M.; et al. Anti transglutaminase antibodies cause ataxia in mice. PLoS ONE 2010, 5, e9698. [Google Scholar] [CrossRef]
- Hadjivassiliou, M.; Grünewald, R.A.; Sanders, D.S.; Shanmugarajah, P.; Hoggard, N. Effect of gluten-free diet on cerebellar MR spectroscopy in gluten ataxia. Neurology 2017, 89, 705–709. [Google Scholar] [CrossRef]
- Diaconu, G.; Burlea, M.; Grigore, I.; Anton, D.; Trandafir, L. Celiac disease with neurologic manifestations in children. Rev. Med. Chir. Soc. Med. Nat. Iasi 2013, 117, 88–94. [Google Scholar] [PubMed]
- Honnorat, J.; Saiz, A.; Giometto, B.; Vincent, A.; Brieva, L.; de Andres, C.; Maestre, J.; Fabien, N.; Vighetto, A.; Casamitjana, R.; et al. Cerebellar Ataxia With Anti–Glutamic Acid Decarboxylase Antibodies. Arch. Neurol. 2001, 58, 225–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baizabal-Carvallo, J.F.; Jankovic, J. Stiff-person syndrome: Insights into a complex autoimmune disorder. J. Neurol. Neurosurg. Psychiatry 2015, 86, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, H.; Manto, M.; Hampe, C.S. Pathogenic Roles of Glutamic Acid Decarboxylase 65 Autoantibodies in Cerebellar Ataxias. J. Immunol. Res. 2017, 2017, e2913297. [Google Scholar] [CrossRef]
- Mitoma, H.; Ishida, K.; Shizuka-Ikeda, M.; Mizusawa, H. Dual impairment of GABAA- and GABAB-receptor-mediated synaptic responses by autoantibodies to glutamic acid decarboxylase. J. Neurol. Sci. 2003, 208, 51–56. [Google Scholar] [CrossRef]
- Ishida, K.; Mitoma, H.; Song, S.Y.; Uchihara, T.; Inaba, A.; Eguchi, S.; Kobayashi, T.; Mizusawa, H. Selective suppression of cerebellar GABAergic transmission by an autoantibody to glutamic acid decarboxylase. Ann. Neurol. 1999, 46, 263–267. [Google Scholar] [CrossRef]
- Ishida, K.; Mitoma, H.; Mizusawa, H. Reversibility of cerebellar GABAergic synapse impairment induced by anti-glutamic acid decarboxylase autoantibodies. J. Neurol. Sci. 2008, 271, 186–190. [Google Scholar] [CrossRef]
- Tian, N.; Petersen, C.; Kash, S.; Baekkeskov, S.; Copenhagen, D.; Nicoll, R. The role of the synthetic enzyme GAD65 in the control of neuronal γ-aminobutyric acid release. Proc. Natl. Acad. Sci. USA 1999, 96, 12911–12916. [Google Scholar] [CrossRef] [Green Version]
- Asada, H.; Kawamura, Y.; Maruyama, K.; Kume, H.; Ding, R.G.; Ji, F.Y.; Kanbara, N.; Kuzume, H.; Sanbo, M.; Yagi, T.; et al. Mice lacking the 65 kDa isoform of glutamic acid decarboxylase (GAD65) maintain normal levels of GAD67 and GABA in their brains but are susceptible to seizures. Biochem. Biophys. Res. Commun. 1996, 229, 891–895. [Google Scholar] [CrossRef]
- Dinkel, K.; Meinck, H.M.; Jury, K.M.; Karges, W.; Richter, W. Inhibition of γ-aminobutyric acid synthesis by glutamic acid decarboxylase autoantibodies in stiff-man syndrome. Ann. Neurol. 1998, 44, 194–201. [Google Scholar] [CrossRef]
- Massie, A.; Boillee, S.; Hewett, S.; Knackstedt, L.; Lewerenz, J. Main path and byways: Non-vesicular glutamate release by system xc—As an important modifier of glutamatergic neurotransmission. J. Neurochem 2015, 135, 1062–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.T.; Wang, M.; Li, W. Regulation of microglia by ionotropic glutamatergic and GABAergic neurotransmission. Neuron Glia Biol. 2012, 7, 41–46. [Google Scholar] [CrossRef]
- Ishida, K.; Mitoma, H.; Wada, Y.; Oka, T.; Shibahara, J.; Saito, Y.; Murayama, S.; Mizusawa, H. Selective loss of Purkinje cells in a patient with anti-glutamic acid decarboxylase antibody-associated cerebellar ataxia. J. Neurol. Neurosurg. Psychiatry 2007, 78, 190–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampe, C.S.; Petrosini, L.; De Bartolo, P.; Caporali, P.; Cutuli, D.; Laricchiuta, D.; Foti, F.; Radtke, J.R.; Vidova, V.; Honnorat, J.; et al. Monoclonal antibodies to 65kDa glutamate decarboxylase induce epitope specific effects on motor and cognitive functions in rats. Orphanet J. Rare Dis. 2013, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Vega-Flores, G.; Rubio, S.E.; Jurado-Parras, M.T.; Gómez-Climent, M.Á.; Hampe, C.S.; Manto, M.; Soriano, E.; Pascual, M.; Gruart, A.; Delgado-García, J.M. The GABAergic septohippocampal pathway is directly involved in internal processes related to operant reward learning. Cereb. Cortex 2014, 24, 2093–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, A.R.; Baquet, Z.; Eisenbarth, G.S.; Tisch, R.; Workman, C.J.; Vignali, D.A.A. CNS Destruction Mediated by GAD-Specific CD4+ T Cells. J. Immunol. 2011, 184, 4863–4870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arino, H.; Hoftberger, R.; Gresa-Arribas, N.; Martinez-Hernandez, E.; Armangue, T.; Kruer, M.C.; Arpa, J.; Domingo, J.; Rojc, B.; Bataller, L.; et al. Paraneoplastic Neurological Syndromes and Glutamic Acid Decarboxylase Antibodies. JAMA Neurol. 2015, 72, 874–881. [Google Scholar] [CrossRef]
- Hildebrand, M.E.; Isope, P.; Miyazaki, T.; Nakaya, T.; Garcia, E.; Feltz, A.; Schneider, T.; Hescheler, J.; Kano, M.; Sakimura, K.; et al. Functional coupling between mGluR1 and Cav3.1 T-type calcium channels contributes to parallel fiber-induced fast calcium signaling within Purkinje cell dendritic spines. J. Neurosci. 2009, 29, 9668–9682. [Google Scholar] [CrossRef] [Green Version]
- Sillevis Smitt, P.; Kinoshita, A.; De Leeuw, B.; Moll, W.; Coesmans, M.; Jaarsma, D.; Henzen-Logmans, S.; Vecht, C.; De Zeeuw, C.; Sekiyama, N.; et al. Paraneoplastic cerebellar ataxia due to autoantibodies against a glutamate receptor. N. Engl. J. Med. 2000, 342, 21–27. [Google Scholar] [CrossRef]
- Iorio, R.; Damato, V.; Mirabella, M.; Vita, M.G.; Hulsenboom, E.; Plantone, D.; Bizzarro, A.; Del Grande, A.; Sillevis Smitt, P.A.E. Cerebellar degeneration associated with mGluR1 autoantibodies as a paraneoplastic manifestation of prostate adenocarcinoma. J. Neuroimmunol. 2013, 263, 155–158. [Google Scholar] [CrossRef]
- Marignier, R.; Chenevier, F.; Rogemond, V.; Smitt, P.S.; Renoux, C.; Cavillon, G.; Androdias, G.; Vukusic, S.; Graus, F.; Honnorat, J.; et al. Metabotropic glutamate receptor type 1 autoantibody-associated cerebellitis: A primary autoimmune disease? Arch. Neurol. 2010, 67, 627–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coesmans, M.; Sillevis Smitt, P.A.; Linden, D.J.; Shigemoto, R.; Hirano, T.; Yamakawa, Y.; Van Alphen, A.M.; Luo, C.; Van der Geest, J.N.; Kros, J.M.; et al. Mechanisms underlying cerebellar motor deficits due to mGluR1-autoantibodies. Ann. Neurol. 2003, 53, 325–336. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Autoantigen | Clinical Presentation | Association with Cancer |
|---|---|---|
| AP3B2 | CA, peripheral neuropathy | Unclear |
| Ca/ARHGAP26 | CA | Rare |
| CARP VIII | CA | Breast cancer, ovarian cancer, colorectal cancer, SCLC |
| Caspr2 | CA, LE, Morvan syndrome | SCLC, thymoma |
| CV2/CRMP5 | CA | SCLC, thymoma |
| GAD65 | LE, CA | SCLC, neuroendocrine tumors, thymoma |
| GFAP | CA | Thymoma, ovarian, prostate, breast cancer |
| GluRdelta 2 | CA, LE | Unclear |
| GlyR | CA, encephalitis | Thymoma, breast cancer, Hodgkin’s lymphoma, SCLC |
| Homer3 | CA, encephalitis | SCLC |
| Hu | CA, LE | SCLC |
| IgLON5 | CA | None |
| ITPR1 | CA, encephalitis, | Breast cancer |
| Yo | CA | Breast, ovarian cancer |
| KLHL11 | Brainstem syndrome, CA | Testicular cancer |
| LGl1 | CA, encephalitis | Thymoma, neuroendocrine tumors |
| Ma2 | LE, CA, brainstem encephalitis | Testis and lung cancer |
| mGluR1 | CA | Hodgkin’s lymphoma |
| mGluR2 | CA | unclear |
| Neuro chondrin | CA | None |
| PCA-2 | Limbic/brainstem encephalitis, LEMS, CA | SCLC |
| Ri | CA | Breast cancer |
| Septin 5 | CA | None |
| SEZ6L2 | CA | Unclear |
| TG 2, 6 | Gluten ataxia | None |
| Tr/DNER | CA, LE | Hodgkin’s lymphoma |
| TRIM 46, 9 & 67 | CA | SCLC |
| VGCC | CA, LEMS | SCLC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hampe, C.S.; Mitoma, H. A Breakdown of Immune Tolerance in the Cerebellum. Brain Sci. 2022, 12, 328. https://doi.org/10.3390/brainsci12030328
Hampe CS, Mitoma H. A Breakdown of Immune Tolerance in the Cerebellum. Brain Sciences. 2022; 12(3):328. https://doi.org/10.3390/brainsci12030328
Chicago/Turabian StyleHampe, Christiane S., and Hiroshi Mitoma. 2022. "A Breakdown of Immune Tolerance in the Cerebellum" Brain Sciences 12, no. 3: 328. https://doi.org/10.3390/brainsci12030328
APA StyleHampe, C. S., & Mitoma, H. (2022). A Breakdown of Immune Tolerance in the Cerebellum. Brain Sciences, 12(3), 328. https://doi.org/10.3390/brainsci12030328