Abstract
Introduction: PTEN gene mutations are frequently found in the genetic landscape of high-grade gliomas since they influence cell proliferation, proangiogenetic pathways, and antitumoral immune response. The present bioinformatics analysis explores the PTEN gene expression profile in HGGs as a prognostic factor for survival, especially focusing on the related immune microenvironment. The effects of PTEN mutation on the susceptibility to conventional chemotherapy were also investigated. Methods: Clinical and genetic data of GBMs and normal tissue samples were acquired from The Cancer Genome Atlas (TCGA)-GBM and Genotype-Tissue Expression (GTEx) online databases, respectively. The genetic differential expressions were analyzed in both groups via the one-way ANOVA test. Kaplan–Meier survival curves were applied to estimate the overall survival (OS) and disease-free survival (DFS). The Genomics of Drug Sensitivity in Cancer platform was chosen to assess the response of PTEN-mutated GBMs to temozolomide (TMZ). p < 0.05 was fixed as statistically significant. On Tumor Immune Estimation Resource and Gene Expression Profiling Interactive Analysis databases, the linkage between immune cell recruitment and PTEN status was assessed through Spearman’s correlation analysis. Results: PTEN was found mutated in 22.2% of the 617 TCGA-GBMs patients, with a higher log2-transcriptome per million reads compared to the GTEx group (255 samples). Survival curves revealed a worse OS and DFS, albeit not significant, for the high-PTEN profile GBMs. Spearman’s analysis of immune cells demonstrated a strong positive correlation between the PTEN status and infiltration of Treg (ρ = 0.179) and M2 macrophages (ρ = 0.303). The half-maximal inhibitor concentration of TMZ was proven to be lower for PTEN-mutated GBMs compared with PTEN wild-types. Conclusions: PTEN gene mutations prevail in GBMs and are strongly related to poor prognosis and least survival. The infiltrating immune lymphocytes Treg and M2 macrophages populate the glioma microenvironment and control the mechanisms of tumor progression, immune escape, and sensitivity to standard chemotherapy. Broader studies are required to confirm these findings and turn them into new therapeutic perspectives.
1. Introduction
High-grade gliomas (HGGs) are common neoplasms of the central nervous system accounting for 70% of brain tumors [1,2,3,4,5]. Glioblastoma (GBM) is the most lethal and represents 60% of newly diagnosed gliomas in the adult population [6,7]. The current standard of care in the management of HGGs is maximum surgical resection, adjuvant chemoradiation, and six cycles of temozolomide [8].
Despite advances in surgical techniques, diagnostics, and target therapeutic strategies, the 5-year survival rate persists under 10% and the median overall survival (OS) still ranges between 14 and 16 months [9]. The poor prognosis and high mortality rate of GBM are attributable to aberrant angiogenesis, extreme mitotic activity, immune escape mechanisms, and intrinsic genome-wide heterogeneity [10,11,12,13,14,15,16,17]. In 2021, Louis and colleagues published the fifth edition of the WHO classification of brain cancers, which reflects the advances in translational medicine, taxonomy, and genetics in neuro-oncology.
They reported a novel tumors nomenclature aimed at integrating histological features, key diagnostic genes, and molecular characteristics underlying oncogenesis [18].
Phosphatase and tensin homolog (PTEN), a tumor suppressor gene, is closely involved in cell translation, proliferation, and tumorigenesis [19,20,21,22]. PTEN protein blocks the intracellular pathways of phosphatidylinositol 3-kinase/AKT/mammalian target of rapamycin (PI3K/AKT/mTOR) via dephosphorylation of phosphatidylinositol-3-triphosphate, resulting in inhibition of the cell cycle [23,24,25,26].
Moreover, the PTEN pathway has a close interaction with the Wnt/β-catenin signals involved in embryogenesis and the determination of neural patterning [22].
Lack/mutation of the PTEN gene, found in 40% of GBMs, influences neurogenesis, and gliogenesis and heightens the DNA damage repairing and the malignant progression of brain tumors [20,25,27].
The prognostic significance of the PTEN gene is also related to the maintenance of the immune microenvironment [28]. Recent transcriptomic pieces of evidence support the correlation between PTEN mutation and the amendment of immune infiltrating cells expression [29,30]. The immune suppression mechanisms intrude on the host antitumor responses and are liable for the failure of conventional therapeutic approaches [31,32,33,34,35]. The number of reported studies on the immunogenomics and immunosuppressive microenvironment of HGGS opened the way for tailored treatments against glioma resilience [36,37,38,39]. Based on these assumptions, we conducted a bioinformatics analysis to examine the PTEN gene expression profile in HGGs as a prognostic factor for survival. We investigated the cluster of the immune infiltrates within the PTEN-related microenvironment intending to identify the contribution of each subpopulation to the immune escape mechanisms. The effects of PTEN mutation on the sensitivity to standard chemotherapy were also explored.
2. Materials and Methods
2.1. Data Acquisition
Transcriptomes, genetic, and clinical features of HGGs-patients were extracted from The Cancer Genome Atlas (TCGA)-GBM project (https://portal.gdc.cancer.gov) (accessed on 31 October 2021) [40]. The genetic data of normal brain tissue samples were acquired from the Genotype-Tissue Expression (GTEx) online database (https://gtexportal.org) (accessed on 1 November 2021).
2.2. Statistical Bioinformatics Analysis
R (https://www.r-project.org) (accessed on 7 March 2022) and Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA) software were used for statistical analysis. Continuous and categorical variables were reported as mean and percentages, respectively.
Top mutation trends and gene patterns were estimated in the TCGA-GBM cohort, focusing on nucleotide variations of the PTEN gene. The differential expressions of PTEN mRNA levels were determined in TCGA-GBM and GTEx groups with the aim to assess the significance of the PTEN gene in the glioma genome compared to healthy brain tissue. A one-way ANOVA test was used for the analysis.
Kaplan–Meier survival curves were used to estimate the prognostic value of the PTEN gene mutations. The overall survival (OS), disease-free survival (DFS), and comparisons between the high- or low-PTEN mutation profile in TCGA-GBM patients were assessed using the log-rank test. Hazard ratios (HDs) were calculated with the Cox proportional risk regression model. The Genomics of Drug Sensitivity in Cancer (GDSC) database (https://www.cancerrxgene.org/) (accessed on 15 November 2021) was the source used for the appraisal of the chemotherapeutic response of PTEN-mutated HGGs [41]. The R software package “Prophetic” was used and the half-maximal inhibitor concentration (IC50) of temozolomide (TMZ) was assessed by one-way ANOVA analysis [41]. p-value was set at <0.05 for all the tests.
2.3. Estimation of Immune Infiltrating Cells
On behalf of the glioma immune microenvironment, the T cells CD4+, CD8+, Treg, NK cells, monocytes/macrophages, and tumor-infiltrating endothelial cells were considered in the TCGA-GBM cohort. Tumor Immune Estimation Resource 2.0 (TIMER2.0) was used to identify the correlation between mRNA PTEN expression and the transcriptional profile of each tumor immune cell in the TCGA-GBM project.
Assuming the purity adjustment, Spearman’s correlation method was employed, where rho (ρ) > 0 and ρ < 0 denoted a positive and negative correlation between the variables, respectively. Based on the Gene Expression Profiling Interactive Analysis (GEPIA), the differential subexpression of mRNA PTEN in the immune subtypes, for both TCGA-GBM and GTEx samples, was clustered with the one-way ANOVA method, and p < 0.05 was assumed as statistical. The results were reported as boxplots.
3. Results
3.1. Demographics and Gene Mutation Profiles
The genetic and clinical data of 617 HGGs were collected by the TCGA-GBM project. The average patients’ age was 58.8 ± 14 years; males were 59.5%, and 96.3% were Caucasian. All the tumors were supratentorial glioblastoma. Chemotherapy was administered as adjuvant treatment in 52.2% of patients, while 45% were treated with concomitant radiotherapy. The average follow-up was 14.7 months, and 69.2% of patients were dead. Overall data about TCGA-GBM patients are summarized in Table 1.

Table 1.
Demographic, clinical, and histological data of TCGA-GBM patients.
PTEN was the most frequent gene by the transcriptome examination (22.2%). It was followed by TTN (20.75%), TP53 (20.10%), EGFR (17.18%), FLG (12.64%), MUC16 (11.51%), NF1 (8.27%), RYR2 (7.62%), PKHD1 (7.29%), HMCN1 (7.29%), SYNE1 (7.29%), SPTA1 (6.97%), PIK3R1 (6.97%), RB1 (6.81%), ATRX (6.65%), IDH (6.62%), PIK3CA (6.48%), OBSCN (6.48%), APOB (6.32%), FLG2 (6.32%), and LRP2 (6.16%) (Figure 1).
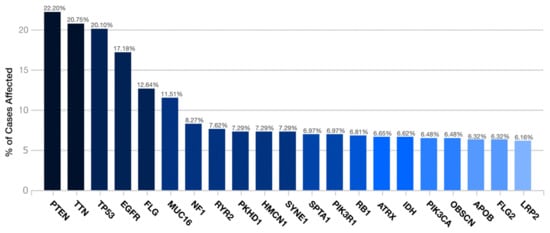
Figure 1.
Distribution of the most frequent mutated genes in the TCGA-GBM project.
Expression profiles and mutations of the top 50 genes in TCGA-GBM are shown in the Oncogrid (Figure 2).
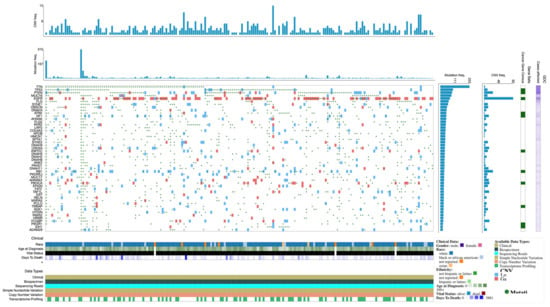
Figure 2.
OncoGrid of top 50 mutated genes with impact mutations on the TCGC-GBM cohort.
From the GTEx dataset, a total of 255 samples of normal brain tissue were included. The differential analysis revealed a higher expression of PTEN mRNA levels in the tumor than in normal tissue, albeit not significant, with a log2-transcriptome per million reads (TPM) +1 of 2.8–5.75 and 2.7–4.8 in the TCGA-GBM and GTEx datasets, respectively (Figure 3).
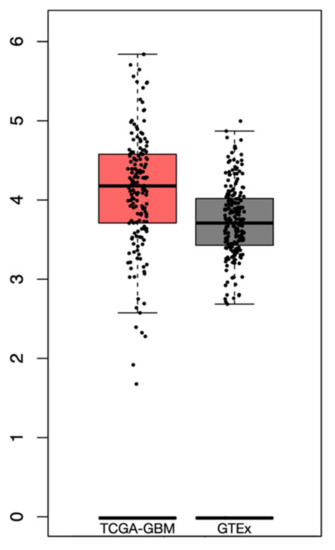
Figure 3.
Box plots revealing the differential PTEN mRNA expression levels in TCGA-GBM and GTEx datasets.
3.2. Survival Analysis
Kaplan–Meier curves showed a worse, though non statistic, OS (Log-rank p = 0.58, HR = 0.91, p = 0.6) and DFS (Log-rank p = 0.78, HR = 1, p = 0.82) in high-PTEN mutation profile (Figure 4).
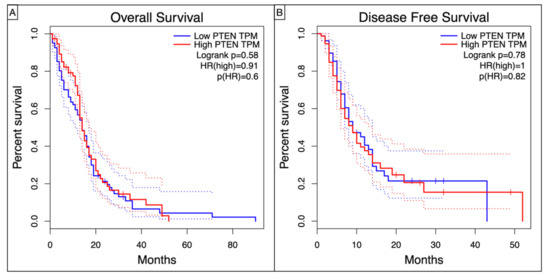
Figure 4.
Kaplan–Meyer curves showing the (A) Overall Survival and (B) Disease Free Survival in TCGA-GBM patients according to the level of PTEN TPM.
3.3. Immune Landscape in PTEN-Related Glioma Microenvironment
Spearman’s correlation analysis of immune subpopulations applied in the TCGA-GBM cohort revealed a negative correlation between mRNA PTEN mutations expression (log2 TPM) and infiltrating T cell CD4+ Th 1 (ρ = −0.245; p = 3.81 × 10−3), Th 2 (ρ = −0.225, p = 8.06 × 10−3), and NK cells (ρ = −0.163, p = 5.64 × 10−2) within the tumor microenvironment. Conversely, Treg (ρ = 0.179, p = 3.54 × 10−2), endothelial cells (ρ = 0.303, p = 2.97 × 10−4), and monocyte/macrophages (ρ = 0.368, p = 9.17 × 10−6) were predominant, with a polarization of M2 (ρ = 0.303, p = 2.97 × 10−4) against of the monocyte (ρ = −0.205, p = 1.6 × 10−2) (Figure 5 and Figure 6).
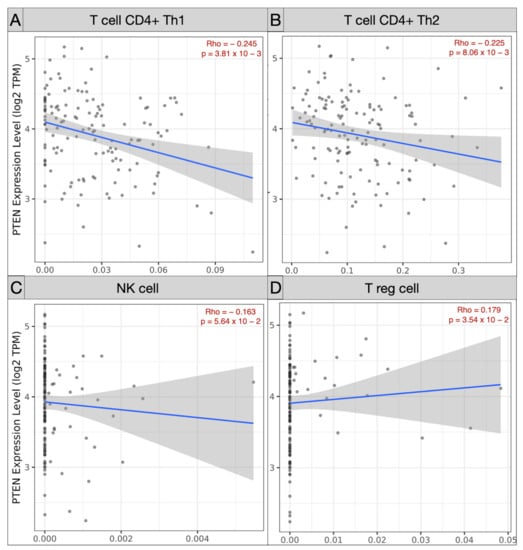
Figure 5.
Scatter plots picturing the correlation of mRNA PTEN expression and the immune infiltration of T cell CD4 Th1 (A), Th2 (B), NK cell (C), and Treg (D) in the TCGC-GBM project.
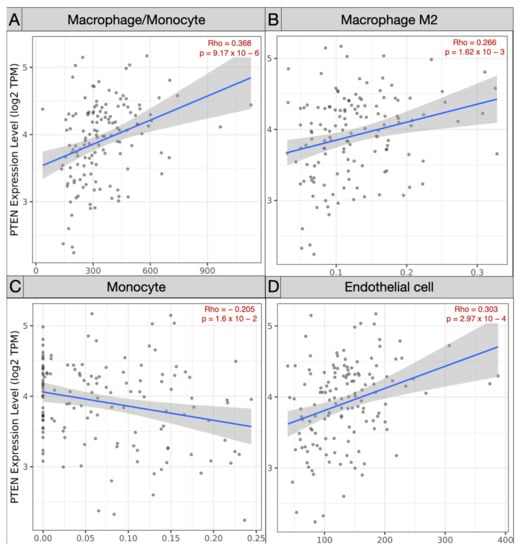
Figure 6.
Scatter plots exhibiting the correlation of mRNA PTEN expression and the immune infiltration of (A) macrophage/monocyte, (B) macrophage M2, (C) monocyte, and (D) endothelial cells in the TCGA-GBM project.
The GEPIA analysis of the mRNA PTEN mutations expression (logTPM +1) in the immune infiltrates of the TCGA-GBM cohort highlighted the decreased density of B cell naive (p < 1 × 10−15), T cell CD4+ naive (p < 1 × 10−15), T cell CD8+ (p = 0.08), and NK cell (p = 3.97 × 10−9) in the glioma microenvironment compared to GTEx samples. The Treg subtype was slightly more represented (p = 0.07).
Furthermore, the boxplots evidenced the monocytes (p < 1 × 10−15), endothelial cells (p < 1 × 10−15), macrophages M0 (p = 3.49 × 10−3), M1 (p < 1 × 10−15), and M2 (p < 1 × 10−15) as the main components of the immune profile at the tumor site in the TCGA-GBM cohort (Figure 7 and Figure 8).
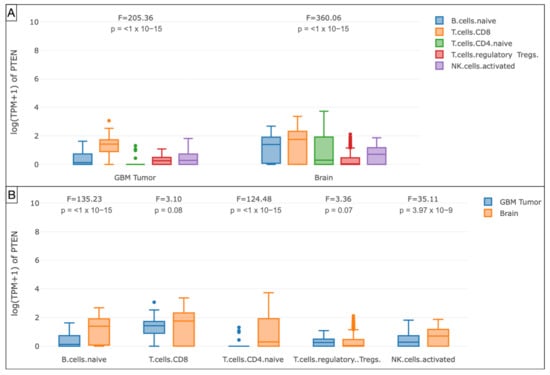
Figure 7.
Box plots showing the immune cell subexpression analysis in PTEN-mutated TCGA-GBMs and normal brain samples from GTEx. (A) Grouped by tissue. (B) Grouped by B cells naive, T cells CD8, CD4, Treg, and NK cells.
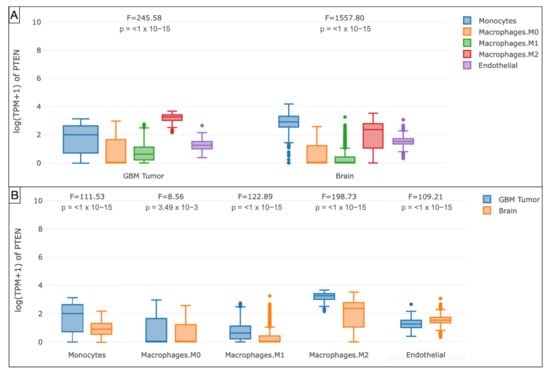
Figure 8.
Box plots illustrating the immune cell subexpression analysis in PTEN-mutated TCGA-GBMs and normal brain samples from GTEx. (A) Grouped by tissue. (B) Grouped by monocytes, macrophages M0, M1, and M2, and endothelial cells.
3.4. Prediction of Chemotherapeutic Response
Based on the GDSC pharmacogenomic database, lower differential targeted responses to TMZ were found in PTEN-mutated and PTEN wild-type samples, with a median IC50 of 531.13 and 701.55 μM, respectively (Figure 9).
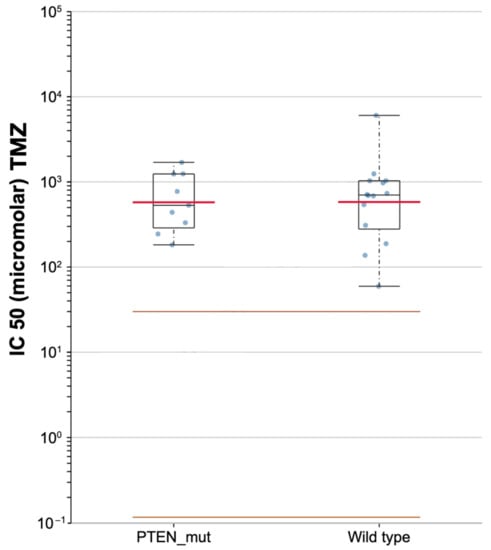
Figure 9.
Scatter plot for the sensitivity to temozolomide (TMZ) in PTEN mutated and wild-type of TCGA-GBMs.
4. Discussion
The present study analyzes mutation profiles and immune signatures of PTEN-associated microenvironment estimating GBM patients’ prognosis, survival, and chemotherapy response. The PTEN gene regulates the cell cycle and DNA repair mechanisms. Its expression modulates cell proliferation, neural development, and gliogenesis [21,27,42].
PTEN mutations are hallmarks of glioma malignancy and influence the patients’ survival [43,44,45,46,47,48,49]. As recently reported by Erira and his group, alterations of PTEN genes are related to glioma proliferation. PTEN mutation may induce post-translational changes in low-grade gliomas, leading to malignant progression [50].
The prognostic role of the PTEN status has been widely deepened in the literature. In 2001, Sasaki et al. analyzed the different median survival of glioma patients as it related to the PTEN expression. They reported a better OS for wild-type PTEN gliomas (123.4 months), compared to the mutated ones (14.8 months) [51]. In 2016, Han and colleagues explored the genetic linkage between PTEN expression and patients’ outcomes in a meta-analysis where a worse prognosis was revealed for PTEN-mutated gliomas [49]. Zhang and his group, 2021, conducted an online bioinformatics analysis about PTEN mutation as a prognostic signature for HGGs. They designed a tailored risk score based on the individual PTEN status, aiming to simplify HGGs diagnosis, prognosis, and treatment planning. They identified 14 independent prognostic genes in PTENwild-type tumors and 3 for the PTEN-mutated ones. These last proved to be related to the worse survival [52].
Even without any statistical significance, our Kaplan–Meier analysis confirmed dismal OS (Log-rank p = 0.58, HR = 0.91, p = 0.6) and DFS (Log-rank p = 0.78, HR = 1, p = 0.82) for the high-PTEN mutation profiles.
Amid the PTEN-related genetic mechanisms underlying the tumorigenesis, the maintenance of the glioma immune microenvironment is critical. Based on this rationale, our analysis also aimed at typifying the subpopulations in the glioma immune niche.
Our Spearman’s correlation tests of immune subpopulations reported a negative correlation (ρ < 0) between PTEN mutations and the expression of infiltrating T cell CD4+ Th 1 (ρ = −0.245), Th 2 (ρ = −0.225), and NK cells (ρ = −0.163) within the tumor microenvironment. The GEPIA one-way ANOVA analysis also revealed a lower density of B cell naive (p < 1 × 10−15), CD8+ (p = 0.08), CD4+ naive (p < 1 × 10−15), and NK cell (p = 3.97 × 10−9) concomitant with TCGA-GBM high-PTEN mutation profiles, compared to the GTEx samples.
In accordance with the evidence in the literature, these data denoted that brain tumor growth and progression are sustained by genetic mechanisms of immune tolerance and exhaustion. The suppression of T, B, and NK cells activity within the PTEN tumor microenvironment suggested immune-mediated biological processes are involved in pathways for glioma immune evasion and resistance to chemotherapies [53,54,55,56].
On the contrary, through Spearman’s analysis the Treg (ρ = 0.179), endothelial cells (ρ = 0.303), and monocyte/macrophages (ρ = 0.368) were found to be prevalent. The differential examination demonstrated the polarization of M2 (ρ = 0.303) within the glioma microenvironment, versus the monocytes (ρ = −0.205). The GEPIA Treg (p = 0.07) and macrophages (p = 3.49 × 10−3) were the most represented subtypes, with a prevalence of M2 in the TCGA-GBM group.
Tregs preserve immune homeostasis, contribute to the downregulation of T cell activity, and regulate innate and adaptive responses against self-antigens, allergens, and infectious agents [57,58,59,60,61]. Tregs also act as immunosuppressive within the tumor microenvironment, repressing the function of CD4+, CD8+, and NK cells [62]. The immune control is carried out by cytokines, extracellular vesicles, perforins, and cytolytic enzymes [63]. They repress antitumor immunity and facilitate immune escape mechanisms, resulting in glioma progression and relapse [64,65,66].
Our results also verified that the immunosuppressive anticancer microenvironment is sustained by the recruitment of monocytes, which in the glioma context are converted into macrophages, with an explicit M2 polarization. M2 macrophages are known to hold an immunosuppressive role [67,68]. M2 phenotypes induce the differential expression of receptors, cytokines, and chemokines. They produce IL-10, IL-1, and IL-6, stimulating tumorigenesis and negatively affecting the prognosis [69,70]. The M2 macrophages, detected in perivascular areas, enhance the VEGF and COX2 production resulting in increased and aberrant angiogenesis [71,72,73].
Therefore, Treg and M2 cells stimulate the glioma cell proliferation, invasion, and support immune escape mechanisms [74,75,76,77]. These data were confirmed by our bioinformatic analysis (ρ = 0.3.03).
Opposing our results, the latest study by Zhou and colleagues published in 2022, found the high expression of PTEN related to a better prognosis for HGGs patients [78]. They discovered via the transwell and flow cytometry that the PTEN gene may inhibit the M1/M2 polarization and M2 macrophages recruitment. These data suggest a potential positive role of the PTEN as an antitumoral immunoregulatory gene [78].
The discrepancy in our study can be explained by the distinct patients cohorts involved, such as the Chinese Glioma Genome Atlas (CGGA) database, and the different techniques applied for data analysis. However, above all, they explored the effects of different PTEN statuses, split into PTEN deletion, PTEN mutated, and PTEN wild-type. This distinction allowed us to assess the specific impact of each PTEN gene expression on glioma immunity. The study demonstrates that, despite advances in genomics, further research is needed to shed light on PTEN activity and its immunological role in tumor progression.
The composition of immune infiltrates explains the prognosis concomitant to the PTEN status and, above all, the ineffective response to standard therapies. TMZ is currently the first line of treatment for HGGs in combination with surgery and radiotherapy [79,80]. It is still debated whether the PTEN mutation may influence sensitivity to radiochemotherapy [81,82,83]. PTEN controls the Wnt/β-catenin and PI3K/Akt/mTOR signaling pathways and arrests the cell cycle at the G2/M phase. TMZ alkylates DNA at this stage, hence PTEN overexpression affects the complex biochemical mechanism of drug alkylation encouraging TMZ activity [84,85].
In 2012 Carico and colleagues conducted a clinical study involving newly diagnosed GBM treated with TMZ. They reported greater effectiveness of TMZ in GBMs with PTEN loss. Inaba et al. investigated the effects of TMZ related to PTEN status founding an increased efficacy in cases of PTEN mutation [83]. Similarly, our analysis of the GDSC database revealed a lower IC50 for PTEN-mutated GBMs (531.13 μM) in comparison with wild-types (701.55 μM).
Apart from conventional chemotherapy, the identification of immune phenotypes and molecular interactions within the tumor microenvironment has been recognized as crucial to widening the spectrum of tailored strategies against the immune escape mechanisms [86,87,88].
Limitations of the Study
The present study has several undeniable limitations, among which are the relatively limited number of patients and short follow-up (average 14.7 months). Other potential biases were the different patients’ ethnicity, limited data about the histological classification, and heterogeneity of radiochemotherapy regimens.
5. Conclusions
PTEN mutations frequently occur in malignant brain tumors, contributing to their progression, reduced OS, and DFS. Within the glioblastoma microenvironment, the PTEN-related immune landscape mainly consists of Treg and M2 macrophages. They repress the antitumor immune activation and are responsible for the triggering of the glioma cell growth, invasion, and aberrant vasculogenesis.
PTEN expression and related glioma microenvironment also influence the sensitivity to conventional radiochemotherapy.
Prospective and randomized trials are necessary to validate these data and to develop novel target treatments.
Author Contributions
Writing—Original Draft, Conceptualization, Writing—Original Draft, A.G.L.; Writing—Review & Editing, Supervision, Validation, S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Rete Italiana delle Neuroscienze with the Network Research Project (#RCR-2019-23669119-002).
Institutional Review Board Statement
All procedures performed in the study were in accordance with the ethical standards of the institution and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Data Availability Statement
All data are included in the main text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Rouse, C.; Chen, Y.; Dowling, J.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J. CBTRUS statistical report: Primary brain and central nervous system tumors diagnosed in the United States in 2007–2011. Neuro Oncol. 2014, 16 (Suppl. S4), iv1–iv63. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Steliarova-Foucher, E.; Lortet-Tieulent, J.; Rosso, S.; Coebergh, J.W.; Comber, H.; Forman, D.; Bray, F. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries in 2012. Eur. J. Cancer 2013, 49, 1374–1403. [Google Scholar] [CrossRef] [PubMed]
- Soerjomataram, I.; Lortet-Tieulent, J.; Parkin, D.M.; Ferlay, J.; Mathers, C.; Forman, D.; Bray, F. Global burden of cancer in 2008: A systematic analysis of disability-adjusted life-years in 12 world regions. Lancet 2012, 380, 1840–1850. [Google Scholar] [CrossRef]
- Ladomersky, E.; Genet, M.; Zhai, L.; Gritsina, G.; Lauing, K.L.; Lulla, R.R.; Fangusaro, J.; Lenzen, A.; Kumthekar, P.; Raizer, J.J.; et al. Improving vaccine efficacy against malignant glioma. Oncoimmunology 2016, 5, e1196311. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Stetson, L.; Virk, S.M.; Barnholtz-Sloan, J.S. Epidemiology of gliomas. Cancer Treat. Res. 2015, 163, 1–14. [Google Scholar] [CrossRef]
- Wrensch, M.; Minn, Y.; Chew, T.; Bondy, M.; Berger, M.S. Epidemiology of primary brain tumors: Current concepts and review of the literature. Neuro Oncol. 2002, 4, 278–299. [Google Scholar] [CrossRef]
- Leenstra, S.; Oskam, N.T.; Bijleveld, E.H.; Bosch, D.A.; Troost, D.; Hulsebos, T.J. Genetic sub-types of human malignant astrocytoma correlate with survival. Int. J. Cancer 1998, 79, 159–165. [Google Scholar] [CrossRef]
- Attarian, F.; Taghizadeh-Hesary, F.; Fanipakdel, A.; Javadinia, S.A.; Porouhan, P.; PeyroShabany, B.; Fazilat-Panah, D. A Systematic Review and Meta-Analysis on the Number of Adjuvant Temozolomide Cycles in Newly Diagnosed Glioblastoma. Front. Oncol. 2021, 11, 1–6. [Google Scholar] [CrossRef]
- Johnson, D.R.; O’Neill, B.P. Glioblastoma survival in the United States before and during the temozolomide era. J. Neurooncol. 2012, 107, 359–364. [Google Scholar] [CrossRef]
- Eagan, R.T.; Scott, M. Evaluation of prognostic factors in chemotherapy of recurrent brain tumors. J. Clin. Oncol. 1983, 1, 38–44. [Google Scholar] [CrossRef]
- Wen, P.Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [CrossRef]
- Perry, J.R.; Laperriere, N.; O’Callaghan, C.J.; Brandes, A.A.; Menten, J.; Phillips, C.; Fay, M.; Nishikawa, R.; Cairncross, J.G.; Roa, W.; et al. Short-Course Radiation plus Temozolomide in Elderly Patients with Glioblastoma. N. Engl. J. Med. 2017, 376, 1027–1037. [Google Scholar] [CrossRef]
- Cancer Genome Atlas (TCGA) Research Network. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.M.; Gallia, G.L.; et al. An integrated genomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef]
- Palumbo, P.; Lombardi, F.; Siragusa, G.; Dehcordi, S.R.; Luzzi, S.; Cimini, A.; Cifone, M.G.; Cinque, B. Involvement of NOS2 Activity on Human Glioma Cell Growth, Clonogenic Potential, and Neurosphere Generation. Int. J. Mol. Sci. 2018, 19, 2801. [Google Scholar] [CrossRef]
- Palumbo, P.; Lombardi, F.; Augello, F.R.; Giusti, I.; Luzzi, S.; Dolo, V.; Cifone, M.G.; Cinque, B. NOS2 inhibitor 1400W Induces Autophagic Flux and Influences Extracellular Vesicle Profile in Human Glioblastoma U87MG Cell Line. Int. J. Mol. Sci. 2019, 20, 3010. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- James, C.D.; Galanis, E.; Frederick, L.; Kimmel, D.W.; Cunningham, J.M.; Atherton-Skaff, P.J.; O’Fallon, J.R.; Jenkins, R.B.; Buckner, J.C.; Hunter, S.B.; et al. Tumor suppressor gene alterations in malignant gliomas: Histopathological associations and prognostic evaluation. Int. J. Oncol. 1999, 15, 547–553. [Google Scholar] [CrossRef]
- Rasheed, B.K.; Stenzel, T.T.; McLendon, R.E.; Parsons, R.; Friedman, A.H.; Friedman, H.S.; Bigner, D.D.; Bigner, S.H. PTEN gene mutations are seen in high-grade but not in low-grade gliomas. Cancer Res. 1997, 57, 4187–4190. [Google Scholar]
- Yang, Y.; Shao, N.; Luo, G.; Li, L.; Zheng, L.; Nilsson-Ehle, P.; Xu, N. Mutations of PTEN gene in gliomas correlate to tumor differentiation and short-term survival rate. Anticancer Res. 2010, 30, 981–985. [Google Scholar] [PubMed]
- Javadinia, S.A.; Shahidsales, S.; Fanipakdel, A.; Joudi-Mashhad, M.; Mehramiz, M.; Talebian, S.; Maftouh, M.; Mardani, R.; Hassanian, S.M.; Khazaei, M.; et al. Therapeutic potential of targeting the Wnt/β-catenin pathway in the treatment of pancreatic cancer. J. Cell. Biochem. 2018, 120, 6833–6840. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lesche, R.; Li, D.M.; Liliental, J.; Zhang, H.; Gao, J.; Gavrilova, N.; Mueller, B.; Liu, X.; Wu, H. PTEN modulates cell cycle progression and cell survival by regulating phosphatidylinositol 3,4,5,-trisphosphate and Akt/protein kinase B signaling pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6199–6204. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yen, C.; Liaw, D.; Podsypanina, K.; Bose, S.; Wang, S.I.; Puc, J.; Miliaresis, C.; Rodgers, L.; McCombie, R.; et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science 1997, 275, 1943–1947. [Google Scholar] [CrossRef]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef]
- Javadinia, S.A.; Shahidsales, S.; Fanipakdel, A.; Mostafapour, A.; Joudi-Mashhad, M.; Ferns, G.A.; Avan, A. The Esophageal Cancer and the PI3K/AKT/mTOR Signaling Regulatory microRNAs: A Novel Marker for Prognosis, and a Possible Target for Immunotherapy. Curr. Pharm. Des. 2018, 24, 4646–4651. [Google Scholar] [CrossRef]
- Ming, M.; He, Y.Y. PTEN in DNA damage repair. Cancer Lett. 2012, 319, 125–129. [Google Scholar] [CrossRef]
- Yang, Y.; Shao, N.; Luo, G.; Li, L.; Nilsson-Ehle, P.; Xu, N. Relationship between PTEN gene expression and differentiation of human glioma. Scand. J. Clin. Lab. Investig. 2006, 66, 469–475. [Google Scholar] [CrossRef]
- Kitange, G.J.; Templeton, K.L.; Jenkins, R.B. Recent advances in the molecular genetics of primary gliomas. Curr. Opin. Oncol. 2003, 15, 197–203. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, X.; Chen, Z.; Liu, N. Establishment and validation of an immune-based prognostic score model in glioblastoma. Int. Immunopharmacol. 2020, 85, 106636. [Google Scholar] [CrossRef]
- Gieryng, A.; Pszczolkowska, D.; Walentynowicz, K.A.; Rajan, W.D.; Kaminska, B. Immune microenvironment of gliomas. Lab. Investig. 2017, 97, 498–518. [Google Scholar] [CrossRef]
- Wilcox, J.A.; Ramakrishna, R.; Magge, R. Immunotherapy in Glioblastoma. World Neurosurg. 2018, 116, 518–528. [Google Scholar] [CrossRef]
- Grabowski, M.M.; Sankey, E.W.; Ryan, K.J.; Chongsathidkiet, P.; Lorrey, S.J.; Wilkinson, D.S.; Fecci, P.E. Immune suppression in gliomas. J. Neurooncol. 2021, 151, 3–12. [Google Scholar] [CrossRef]
- Hanaei, S.; Afshari, K.; Hirbod-Mobarakeh, A.; Mohajer, B.; Amir Dastmalchi, D.; Rezaei, N. Therapeutic efficacy of specific immunotherapy for glioma: A systematic review and meta-analysis. Rev. Neurosci. 2018, 29, 443–461. [Google Scholar] [CrossRef]
- Giotta Lucifero, A.; Luzzi, S.; Brambilla, I.; Trabatti, C.; Mosconi, M.; Savasta, S.; Foiadelli, T. Innovative therapies for malignant brain tumors: The road to a tailored cure. Acta Biomed. 2020, 91, 5–17. [Google Scholar] [CrossRef]
- Luzzi, S.; Crovace, A.M.; Del Maestro, M.; Giotta Lucifero, A.; Elbabaa, S.K.; Cinque, B.; Palumbo, P.; Lombardi, F.; Cimini, A.; Cifone, M.G.; et al. The cell-based approach in neurosurgery: Ongoing trends and future perspectives. Heliyon 2019, 5, e02818. [Google Scholar] [CrossRef]
- Giotta Lucifero, A.; Luzzi, S. Against the Resilience of High-Grade Gliomas: The Immunotherapeutic Approach (Part I). Brain Sci. 2021, 11, 386. [Google Scholar] [CrossRef]
- Giotta Lucifero, A.; Luzzi, S. Against the Resilience of High-Grade Gliomas: Gene Therapies (Part II). Brain Sci. 2021, 11, 976. [Google Scholar] [CrossRef]
- Luzzi, S.; Giotta Lucifero, A.; Brambilla, I.; Trabatti, C.; Mosconi, M.; Savasta, S.; Foiadelli, T. The impact of stem cells in neuro-oncology: Applications, evidence, limitations and challenges. Acta Biomed. 2020, 91, 51–60. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Yang, W.; Soares, J.; Greninger, P.; Edelman, E.J.; Lightfoot, H.; Forbes, S.; Bindal, N.; Beare, D.; Smith, J.A.; Thompson, I.R.; et al. Genomics of Drug Sensitivity in Cancer (GDSC): A resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 2012, 41, D955–D961. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Molina, A.; Serrano, M. PTEN in cancer, metabolism, and aging. Trends Endocrinol. Metab. 2013, 24, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.S.; Tachibana, I.; Passe, S.M.; Huntley, B.K.; Borell, T.J.; Iturria, N.; O’Fallon, J.R.; Schaefer, P.L.; Scheithauer, B.W.; James, C.D.; et al. PTEN mutation, EGFR amplification, and outcome in patients with anaplastic astrocytoma and glioblastoma multiforme. J. Natl. Cancer Inst. 2001, 93, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Srividya, M.R.; Thota, B.; Shailaja, B.C.; Arivazhagan, A.; Thennarasu, K.; Chandramouli, B.A.; Hegde, A.S.; Santosh, V. Homozygous 10q23/PTEN deletion and its impact on outcome in glioblastoma: A prospective translational study on a uniformly treated cohort of adult patients. Neuropathology 2011, 31, 376–383. [Google Scholar] [CrossRef] [PubMed]
- Koul, D. PTEN signaling pathways in glioblastoma. Cancer Biol. Ther. 2008, 7, 1321–1325. [Google Scholar] [CrossRef] [PubMed]
- D’Urso, O.F.; D’Urso, P.I.; Marsigliante, S.; Storelli, C.; Luzi, G.; Gianfreda, C.D.; Montinaro, A.; Distante, A.; Ciappetta, P. Correlative analysis of gene expression profile and prognosis in patients with gliomatosis cerebri. Cancer 2009, 115, 3749–3757. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, J.M.; Farizan, A.; Asmarina, K.; Zainuddin, N.; Ghazali, M.M.; Jaafar, H.; Isa, M.N.; Naing, N.N. Association of loss of heterozygosity and PTEN gene abnormalities with paraclinical, clinical modalities and survival time of glioma patients in Malaysia. Asian J. Surg. 2006, 29, 274–282. [Google Scholar] [CrossRef][Green Version]
- Xu, J.; Li, Z.; Wang, J.; Chen, H.; Fang, J.Y. Combined PTEN Mutation and Protein Expression Associate with Overall and Disease-Free Survival of Glioblastoma Patients. Transl. Oncol. 2014, 7, 196–205.e191. [Google Scholar] [CrossRef]
- Han, F.; Hu, R.; Yang, H.; Liu, J.; Sui, J.; Xiang, X.; Wang, F.; Chu, L.; Song, S. PTEN gene mutations correlate to poor prognosis in glioma patients: A meta-analysis. Onco Targets Ther. 2016, 9, 3485–3492. [Google Scholar] [CrossRef]
- Erira, A.; Velandia, F.; Penagos, J.; Zubieta, C.; Arboleda, G. Differential Regulation of the EGFR/PI3K/AKT/PTEN Pathway between Low- and High-Grade Gliomas. Brain Sci. 2021, 11, 1655. [Google Scholar] [CrossRef]
- Sasaki, H.; Zlatescu, M.C.; Betensky, R.A.; Ino, Y.; Cairncross, J.G.; Louis, D.N. PTEN is a target of chromosome 10q loss in anaplastic oligodendrogliomas and PTEN alterations are associated with poor prognosis. Am. J. Pathol. 2001, 159, 359–367. [Google Scholar] [CrossRef]
- Zhang, P.; Meng, X.; Liu, L.; Li, S.; Li, Y.; Ali, S.; Li, S.; Xiong, J.; Liu, X.; Li, S.; et al. Identification of the Prognostic Signatures of Glioma With Different PTEN Status. Front. Oncol. 2021, 11, 633357. [Google Scholar] [CrossRef]
- Cordell, E.C.; Alghamri, M.S.; Castro, M.G.; Gutmann, D.H. T lymphocytes as dynamic regulators of glioma pathobiology. Neuro Oncol. 2022, noac055. [Google Scholar] [CrossRef]
- Codrici, E.; Popescu, I.D.; Tanase, C.; Enciu, A.M. Friends with Benefits: Chemokines, Glioblastoma-Associated Microglia/Macrophages, and Tumor Microenvironment. Int. J. Mol. Sci. 2022, 23, 2509. [Google Scholar] [CrossRef]
- DeCordova, S.; Shastri, A.; Tsolaki, A.G.; Yasmin, H.; Klein, L.; Singh, S.K.; Kishore, U. Molecular Heterogeneity and Immunosuppressive Microenvironment in Glioblastoma. Front. Immunol. 2020, 11, 1402. [Google Scholar] [CrossRef]
- Ma, Q.; Long, W.; Xing, C.; Chu, J.; Luo, M.; Wang, H.Y.; Liu, Q.; Wang, R.F. Cancer Stem Cells and Immunosuppressive Microenvironment in Glioma. Front. Immunol. 2018, 9, 2924. [Google Scholar] [CrossRef]
- Grover, P.; Goel, P.N.; Greene, M.I. Regulatory T Cells: Regulation of Identity and Function. Front. Immunol. 2021, 12, 750542. [Google Scholar] [CrossRef]
- Strauss, L.; Bergmann, C.; Whiteside, T.L. Human circulating CD4+CD25highFoxp3+ regulatory T cells kill autologous CD8+ but not CD4+ responder cells by Fas-mediated apoptosis. J. Immunol. 2009, 182, 1469–1480. [Google Scholar] [CrossRef]
- Boer, M.C.; Joosten, S.A.; Ottenhoff, T.H. Regulatory T-Cells at the Interface between Human Host and Pathogens in Infectious Diseases and Vaccination. Front. Immunol. 2015, 6, 217. [Google Scholar] [CrossRef]
- Sakaguchi, S. Regulatory T cells: Key controllers of immunologic self-tolerance. Cell 2000, 101, 455–458. [Google Scholar] [CrossRef]
- Scheinecker, C.; Göschl, L.; Bonelli, M. Treg cells in health and autoimmune diseases: New insights from single cell analysis. J. Autoimmun. 2020, 110, 102376. [Google Scholar] [CrossRef] [PubMed]
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression—Implications for anticancer therapy. Nat. Rev. Clin. Oncol. 2019, 16, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Okeke, E.B.; Uzonna, J.E. The Pivotal Role of Regulatory T Cells in the Regulation of Innate Immune Cells. Front. Immunol. 2019, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, H.; Sakaguchi, S. Regulatory T cells in tumor immunity. Int. J. Cancer 2010, 127, 759–767. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Nishikawa, H. Roles of regulatory T cells in cancer immunity. Int. Immunol. 2016, 28, 401–409. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, B.S.; Lee, S.K. Regulatory T Cells in Tumor Microenvironment and Approach for Anticancer Immunotherapy. Immune Netw. 2020, 20, e4. [Google Scholar] [CrossRef]
- Gajewski, T.F. Identifying and Overcoming Immune Resistance Mechanisms in the Melanoma Tumor Microenvironment. Clin. Cancer Res. 2006, 12, 2326s–2330s. [Google Scholar] [CrossRef]
- Shevach, E.M. CD4+CD25+ suppressor T cells: More questions than answers. Nat. Rev. Immunol. 2002, 2, 389–400. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Mignogna, C.; Signorelli, F.; Vismara, M.F.; Zeppa, P.; Camastra, C.; Barni, T.; Donato, G.; Di Vito, A. A reappraisal of macrophage polarization in glioblastoma: Histopathological and immunohistochemical findings and review of the literature. Pathol. Res. Pract. 2016, 212, 491–499. [Google Scholar] [CrossRef]
- Hughes, R.; Qian, B.Z.; Rowan, C.; Muthana, M.; Keklikoglou, I.; Olson, O.C.; Tazzyman, S.; Danson, S.; Addison, C.; Clemons, M.; et al. Perivascular M2 Macrophages Stimulate Tumor Relapse after Chemotherapy. Cancer Res. 2015, 75, 3479–3491. [Google Scholar] [CrossRef]
- Fan, Y.; Ye, J.; Shen, F.; Zhu, Y.; Yeghiazarians, Y.; Zhu, W.; Chen, Y.; Lawton, M.T.; Young, W.L.; Yang, G.Y. Interleukin-6 stimulates circulating blood-derived endothelial progenitor cell angiogenesis in vitro. J. Cereb. Blood Flow Metab. 2008, 28, 90–98. [Google Scholar] [CrossRef]
- Chen, Y.; Song, Y.; Du, W.; Gong, L.; Chang, H.; Zou, Z. Tumor-associated macrophages: An accomplice in solid tumor progression. J. Biomed. Sci. 2019, 26, 78. [Google Scholar] [CrossRef]
- Hussain, S.F.; Yang, D.; Suki, D.; Aldape, K.; Grimm, E.; Heimberger, A.B. The role of human glioma-infiltrating microglia/macrophages in mediating antitumor immune responses. Neuro Oncol. 2006, 8, 261–279. [Google Scholar] [CrossRef]
- Zhu, C.; Kros, J.M.; Cheng, C.; Mustafa, D. The contribution of tumor-associated macrophages in glioma neo-angiogenesis and implications for anti-angiogenic strategies. Neuro Oncol. 2017, 19, 1435–1446. [Google Scholar] [CrossRef]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef]
- Zhou, F.; Shi, Q.; Fan, X.; Yu, R.; Wu, Z.; Wang, B.; Tian, W.; Yu, T.; Pan, M.; You, Y.; et al. Diverse Macrophages Constituted the Glioma Microenvironment and Influenced by PTEN Status. Front. Immunol. 2022, 13, 841404. [Google Scholar] [CrossRef]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Sano, T.; Lin, H.; Chen, X.; Langford, L.A.; Koul, D.; Bondy, M.L.; Hess, K.R.; Myers, J.N.; Hong, Y.K.; Yung, W.K.; et al. Differential expression of MMAC/PTEN in glioblastoma multiforme: Relationship to localization and prognosis. Cancer Res. 1999, 59, 1820–1824. [Google Scholar] [PubMed]
- McEllin, B.; Camacho, C.V.; Mukherjee, B.; Hahm, B.; Tomimatsu, N.; Bachoo, R.M.; Burma, S. PTEN loss compromises homologous recombination repair in astrocytes: Implications for glioblastoma therapy with temozolomide or poly(ADP-ribose) polymerase inhibitors. Cancer Res. 2010, 70, 5457–5464. [Google Scholar] [CrossRef] [PubMed]
- Inaba, N.; Kimura, M.; Fujioka, K.; Ikeda, K.; Somura, H.; Akiyoshi, K.; Inoue, Y.; Nomura, M.; Saito, Y.; Saito, H.; et al. The effect of PTEN on proliferation and drug-, and radiosensitivity in malignant glioma cells. Anticancer Res. 2011, 31, 1653–1658. [Google Scholar] [PubMed]
- Zając, A.; Sumorek-Wiadro, J.; Langner, E.; Wertel, I.; Maciejczyk, A.; Pawlikowska-Pawlęga, B.; Pawelec, J.; Wasiak, M.; Hułas-Stasiak, M.; Bądziul, D.; et al. Involvement of PI3K Pathway in Glioma Cell Resistance to Temozolomide Treatment. Int. J. Mol. Sci. 2021, 22, 5155. [Google Scholar] [CrossRef]
- Carico, C.; Nuño, M.; Mukherjee, D.; Elramsisy, A.; Dantis, J.; Hu, J.; Rudnick, J.; Yu, J.S.; Black, K.L.; Bannykh, S.I.; et al. Loss of PTEN is not associated with poor survival in newly diagnosed glioblastoma patients of the temozolomide era. PLoS ONE 2012, 7, e33684. [Google Scholar] [CrossRef]
- Giotta Lucifero, A.; Luzzi, S.; Brambilla, I.; Schena, L.; Mosconi, M.; Foiadelli, T.; Savasta, S. Potential roads for reaching the summit: An overview on target therapies for high-grade gliomas. Acta Biomed. 2020, 91, 61–78. [Google Scholar] [CrossRef]
- Giotta Lucifero, A.; Luzzi, S.; Brambilla, I.; Guarracino, C.; Mosconi, M.; Foiadelli, T.; Savasta, S. Gene therapies for high-grade gliomas: From the bench to the bedside. Acta Biomed. 2020, 91, 32–50. [Google Scholar] [CrossRef]
- Luzzi, S.; Giotta Lucifero, A.; Brambilla, I.; Magistrali, M.; Mosconi, M.; Savasta, S.; Foiadelli, T. Adoptive immunotherapies in neuro-oncology: Classification, recent advances, and translational challenges. Acta Biomed. 2020, 91, 18–31. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).