
Frequency-Specific Analysis of the Dynamic Reconfiguration of the Brain in Patients with Schizophrenia

 ,
,  , ,
, ,
Abstract
:1. Introduction
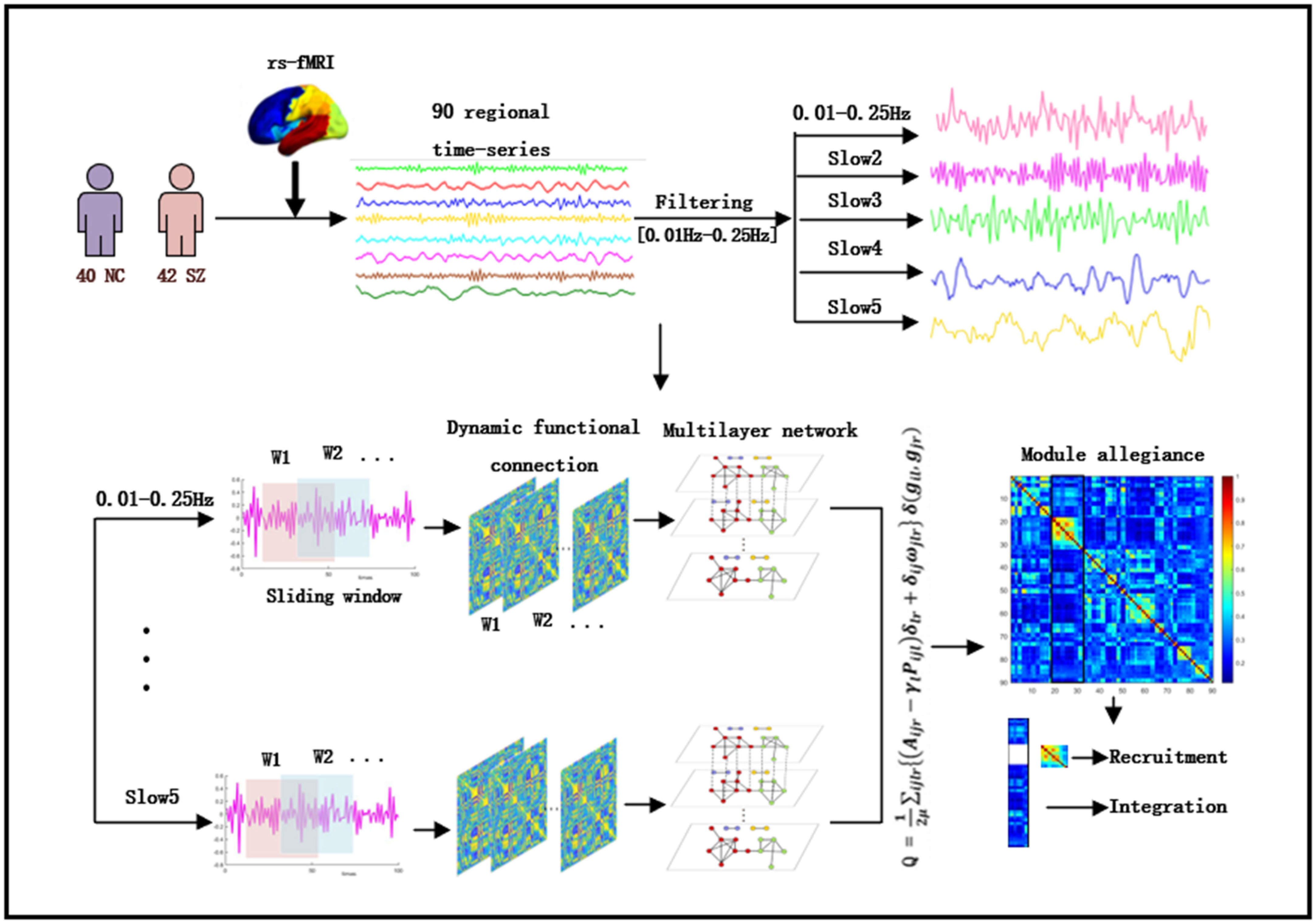
2. Materials and Methods
2.1. Participants
2.2. Imaging Acquisition and Preprocessing
2.3. Formatting of Mathematical Components
2.4. Multilayer Community Detection
2.5. Dynamic Network Statistics
2.5.1. Module Allegiance
2.5.2. Recruitment and Integration
2.6. Statistical Analysis
3. Results
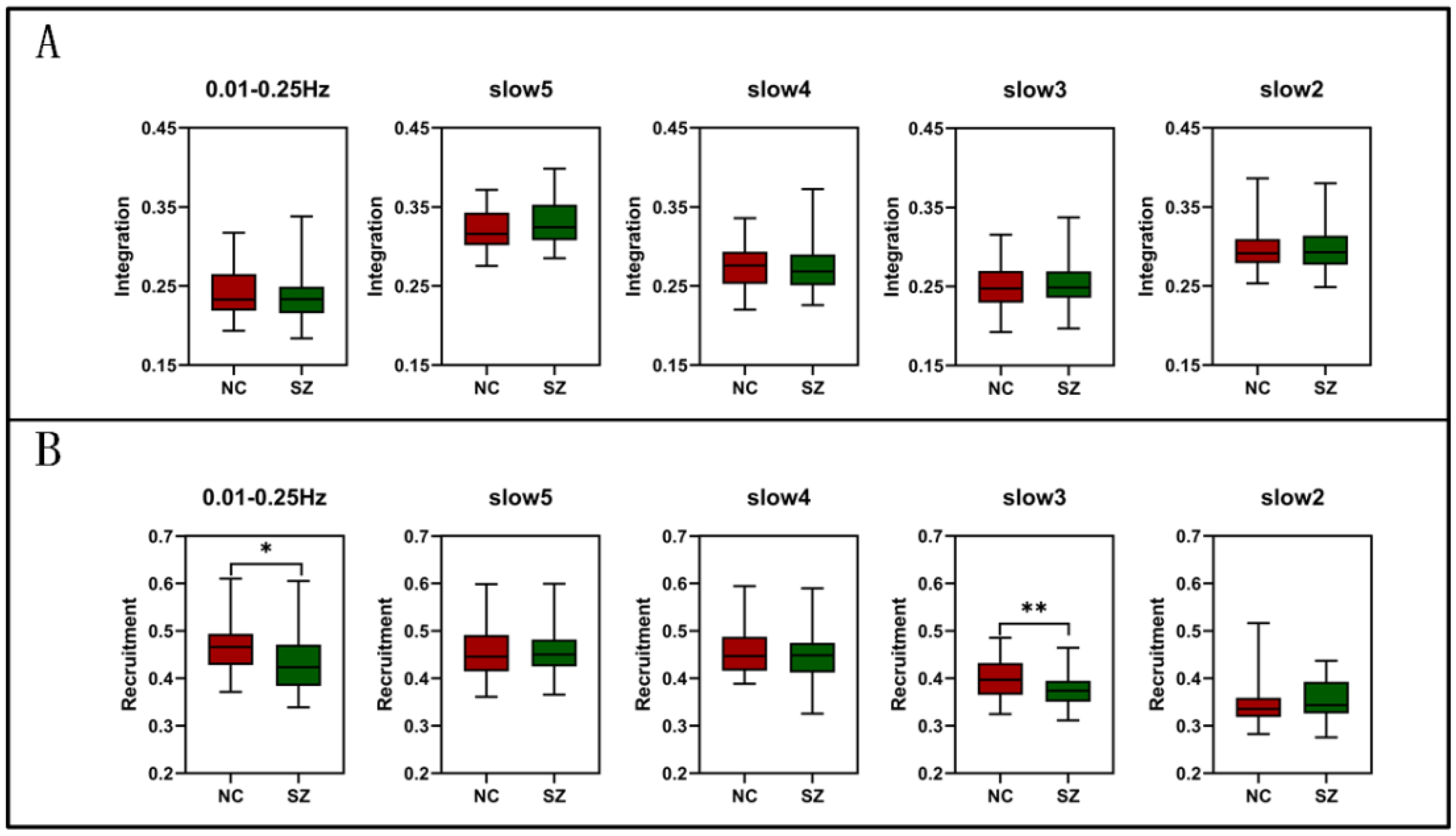
3.1. Group Comparisons of the Whole-Brain Level at Different Frequencies
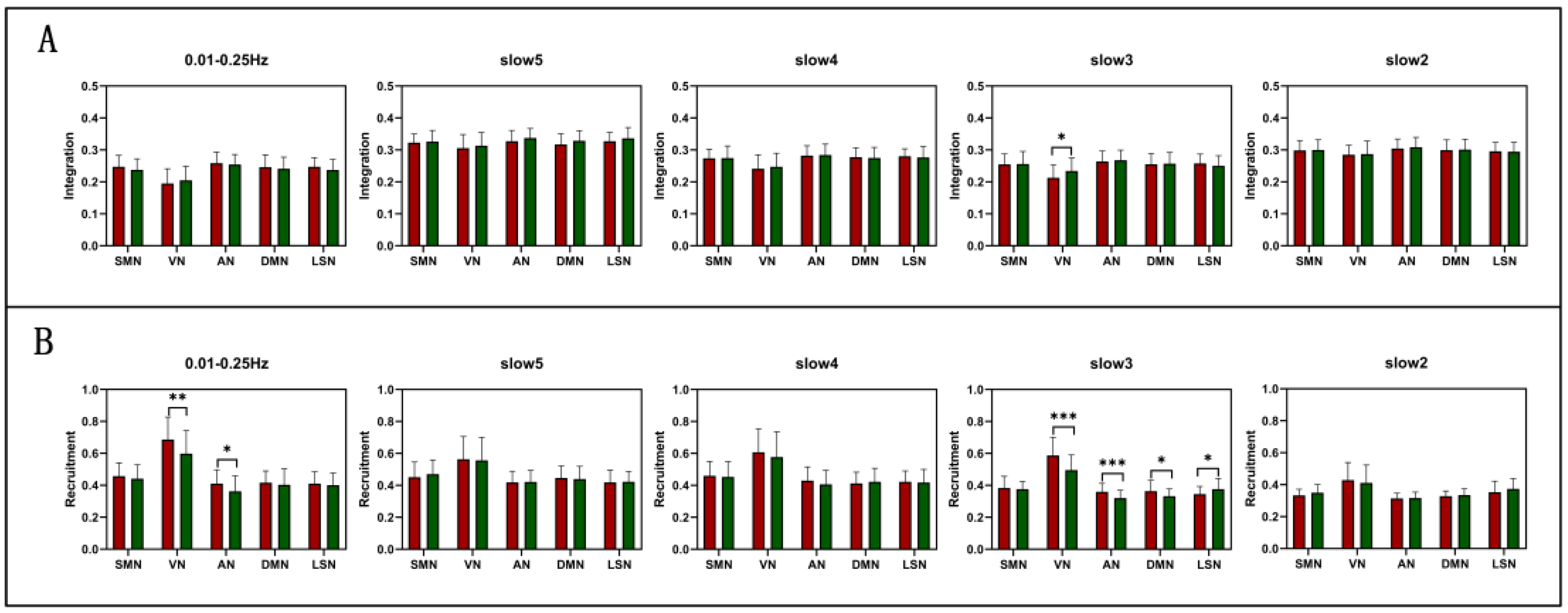
3.2. Group Comparisons of RSN Level at Different Frequencies
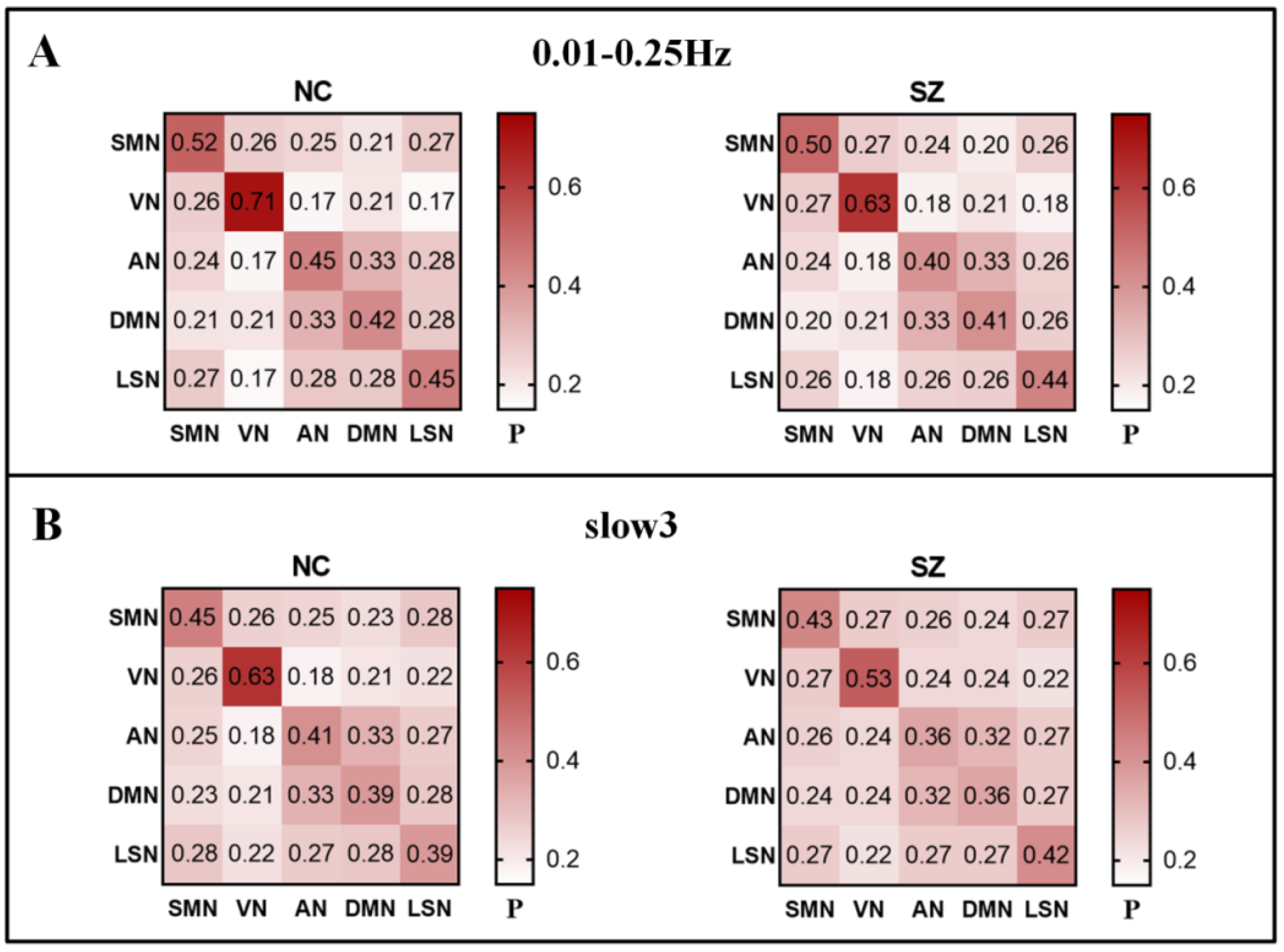
3.3. Group Comparisons of RSN to RSN Integration at Different Frequencies
3.4. Group Comparisons of Node Level at Different Frequencies
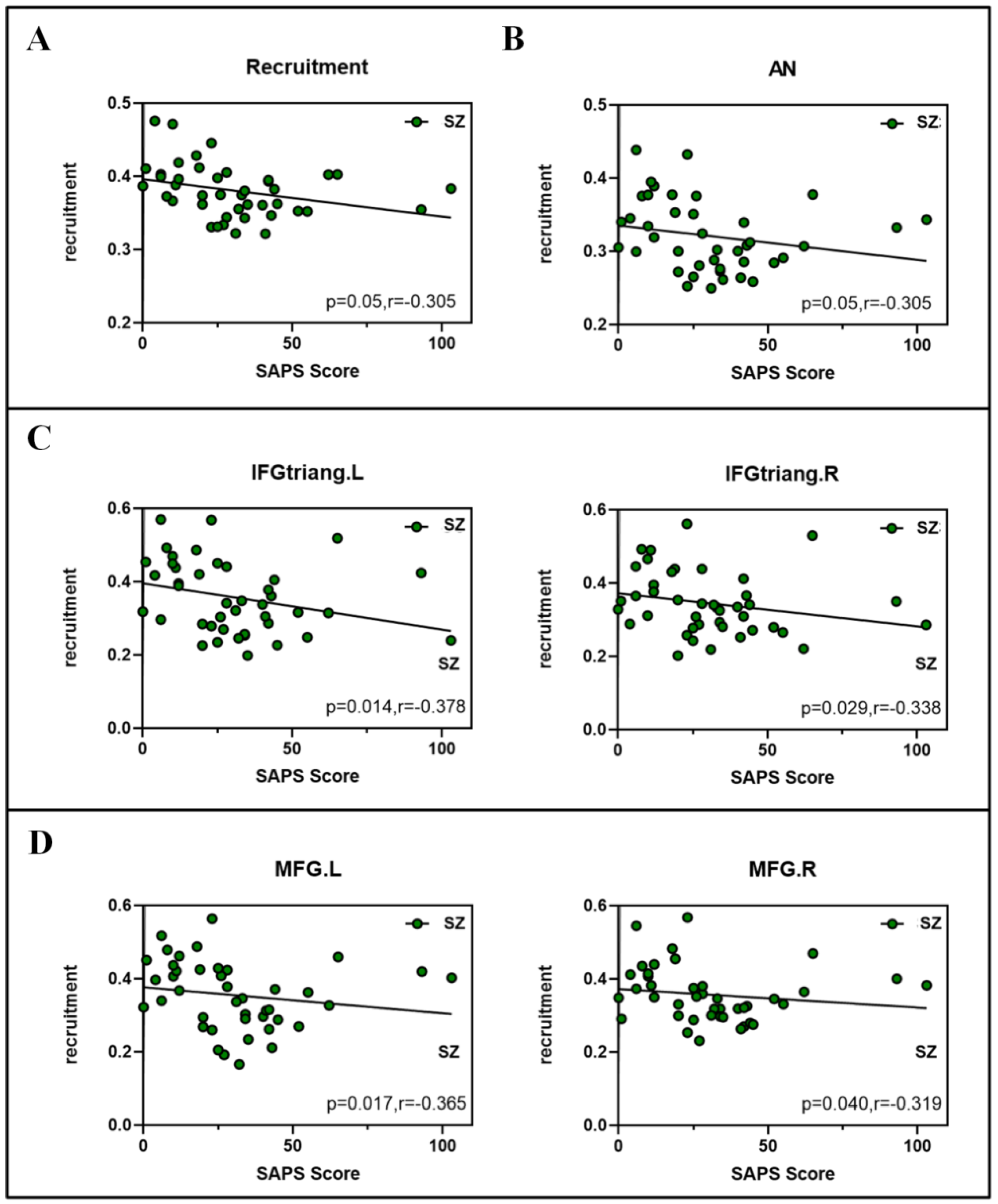
3.5. Correlation between Network Measures and SAPS Scores
4. Discussion
4.1. Reduced Recruitment in SZ Patients
4.2. Abnormal Brain Networks/Regions of Dynamic Reconfiguration in SZ Patients at slow3
4.3. Frequency-Specificity of Multilayer Brain Networks in SZ Patients
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, Y.; He, H.; Duan, M.; Huang, H.; Luo, C.J. Dynamic Functional Connectivity Strength Within Different Frequency-Band in Schizophrenia. Front. Psychiatry 2020, 10, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; He, H.; Duan, M.; Huang, H.; Luo, C.J. Resting-state network connectivity and metastability predict clinical symptoms in schizophrenia. Schizophr. Res. 2018, 201, 208–216. [Google Scholar]
- Wang, M.; Hao, X.; Huang, J.; Wang, K.; Shen, L.; Xu, X.; Liu, M.J.N. Hierarchical Structured Sparse Learning for Schizophrenia Identification. Neuroinformatics 2019, 18, 43–57. [Google Scholar] [CrossRef]
- Zou, H.; Yang, J.J. Multi-frequency Dynamic Weighted Functional Connectivity Networks for Schizophrenia Diagnosis. Appl. Magn. Reson. 2019, 50, 847–859. [Google Scholar] [CrossRef]
- Allen, E.A.; Damaraju, E.; Plis, S.M.; Erhardt, E.B.; Eichele, T.; Calhoun, V.D.J.C.C. Tracking Whole-Brain Connectivity Dynamics in the Resting State. Cereb. Cortex 2014, 24, 663–676. [Google Scholar] [CrossRef]
- Farinha, M.; Amado, C.; Morgado, P.; Cabral, J.J.F. Increased excursions to functional networks in schizophrenia in the absence of task. Front. Neurosci. 2022, 16, 821179. [Google Scholar] [CrossRef]
- Gifford, G.; Crossley, N.; Kempton, M.J.; Morgan, S.; Mcguire, P.J.N.C. Resting State fMRI Based Multilayer Network Configuration in Patients with Schizophrenia. NeuroImage Clin. 2020, 25, 102169. [Google Scholar] [CrossRef]
- Cui, X.; Ding, C.; Wei, J.; Xue, J.; Xiang, J.J. Analysis of Dynamic Network Reconfiguration in Adults with Attention-Deficit/Hyperactivity Disorder Based Multilayer Network. Cereb. Cortex 2021, 31, 4945–4957. [Google Scholar] [CrossRef]
- Braun, U.; SchaFer, A.; Bassett, D.S.; Rausch, F.; Schweiger, J.I.; Bilek, E.; Erk, S.; Romanczuk-Seiferth, N.; Grimm, O.; Geiger, L.S. Dynamic brain network reconfiguration as a potential schizophrenia genetic risk mechanism modulated by NMDA receptor function. Proc. Natl. Acad. Sci. USA 2016, 113, 12568–12573. [Google Scholar] [CrossRef] [Green Version]
- Xiaosong, H.; Bassett, D.S.; Ganne, C.; Sperling, M.R.; Lauren, K.; Tracy, J.I.N. Disrupted dynamic network reconfiguration of the language system in temporal lobe epilepsy. Brain 2018, 141, 1375–1389. [Google Scholar]
- Dong, D.; Duan, M.; Wang, Y.; Zhang, X.; Jia, X.; Li, Y.; Xin, F.; Yao, D.; Luo, C. Reconfiguration of Dynamic Functional Connectivity in Sensory and Perceptual System in Schizophrenia. Brain 2018, 29, 3577–3589. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Yan, Z.; Long, Z.; Zheng, J.; Zhang, Y.; Han, S.; Zhao, J.J. Frequency-specific alteration of functional connectivity density in antipsychotic-naive adolescents with early-onset schizophrenia. J. Psychiatr. Res. 2017, 95, 68–75. [Google Scholar]
- Han, S.; Zong, X.; Hu, M.; Yu, Y.; Wang, X.; Long, Z.; Chen, H.J. Frequency-selective alteration in the resting-state corticostriatal-thalamo-cortical circuit correlates with symptoms severity in first-episode drug-naive patients with schizophrenia. Schizophr. Res. 2017, 189, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Chien, Y.; Wang, H.S.; Liu, C.; Liu, C.; Hwang, T.; Tseng, W.Y. Frequency-specific alternations in the amplitude of low-frequency fluctuations in schizophrenia. Brain Mapp. 2012, 35, 627–637. [Google Scholar] [CrossRef]
- Mattar, M.G.; Cole, M.W.; Thompson-Schill, S.L.; Bassett, D.S. A Functional Cartography of Cognitive Systems. PLoS Comput. Biol. 2015, 11, e1004533. [Google Scholar] [CrossRef] [Green Version]
- Cordes, D.; Haughton, V.M.; Arfanakis, K.; Carew, J.D.; Turski, P.A.; Moritz, C.H.; Wendt, G.J. Mapping functionally related regions of brain with functional connectivity in the cerebral cortex in “resting-state” data. Am. J. Neuroradiol. 2001, 21, 1636–1644. [Google Scholar]
- Zuo, X.N.; Martino, A.D.; Kelly, C.; Shehzad, Z.E.; Gee, D.G.; Klein, D.F.; Milham, M.P.J.N. The oscillating brain: Complex and reliable. NeuroImage 2010, 49, 1432–1445. [Google Scholar] [CrossRef] [Green Version]
- Stam, C.J. Modern network science of neurological disorders. Nat. Rev. Neurosci. 2014, 15, 683. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Joliot, M.J.N. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Yong, H.; Wang, J.; Liang, W.; Chen, Z.J.; Yan, C.; Yang, H.; Zang, Y.J. Uncovering Intrinsic Modular Organization of Spontaneous Brain Activity in Humans. PLoS ONE 2009, 4, e5226. [Google Scholar]
- Leonardi, N.; Ville, D.J.N. On spurious and real fluctuations of dynamic functional connectivity during rest. Neuroimage 2015, 104, 430–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivela, M.; Arenas, A.; Barthelemy, M.; Gl Ee Son, J.P.; Moreno, Y.; Porter, M.A. Multilayer Networks. J. Complex Netw. 2013, 2, 203–271. [Google Scholar] [CrossRef] [Green Version]
- Mucha, P.J.; Richardson, T.; Macon, K.; Porter, M.A.; Onnela, J.P.J.S. Community Structure in Time-Dependent, Multiscale, and Multiplex Networks. Science 2010, 328, 876–878. [Google Scholar] [CrossRef]
- Jutla, I.S.; Jeub, L.G.; Mucha, P.J. A Generalized Louvain Method for Community Detection Implemented in MATLAB. 2011. Available online: http://netwiki.amath.unc.edu/GenLouvain/GenLouvain (accessed on 23 March 2022).
- Bassett, D.S.; Wymbs, N.F.; Rombach, M.P.; Porter, M.A.; Mucha, P.J.; Grafton, S.T. Task-Based Core-Periphery Organization of Human Brain Dynamics. PLoS Comput. Biol. 2013, 9, e1003171. [Google Scholar] [CrossRef] [PubMed]
- Hl, A.; Ke, H.; Ypd, E.; Xt, D.; Meng, W.; Bo, M.A.; Ald, G. Dynamic Reconfiguration of Human Brain Networks Across Altered States of Consciousness. Behav. Brain Res. 2021, 419, 113685. [Google Scholar]
- Ding, C.; Xiang, J.; Cui, X.; Wang, X.; Li, D.; Cheng, C.; Wang, B.J. Abnormal Dynamic Community Structure of Patients with Attention-Deficit/Hyperactivity Disorder in the Resting State. J. Atten. Disord. 2022, 26, 34–47. [Google Scholar] [CrossRef]
- Blondel, V.D.; Guillaume, J.L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008, 2008, P10008. [Google Scholar] [CrossRef] [Green Version]
- Chai, L.R.; Mattar, M.G.; Asher, B.I.; Evelina, F.; Bassett, D.S. Functional Network Dynamics of the Language System. Cereb. Cortex 2016, 26, 4148–4159. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Wylie, G.; Pasternak, R.; Butler, P.D.; Javitt, D.C. Magnocellular contributions to impaired motion processing in schizophrenia. Schizophr. Res. 2006, 82, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nir, Y.; Hasson, U.; Levy, I.; Yeshurun, Y.; Malach, R. Widespread functional connectivity and fMRI fluctuations in human visual cortex in the absence of visual stimulation. NeuroImage 2006, 30, 1313–1324. [Google Scholar] [CrossRef]
- Mendoza, J.E.; Foundas, A.L. The Somatosensory Systems. In Clinical Neuroanatomy: A Neurobehavioral Approach; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Conrad, C.D.; Stumpf, W.E. Direct visual input to the limbic system: Crossed retinal projections to the nucleus anterodorsalis thalami in the tree shrew. Exp. Brain Res. 1975, 23, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Kang, J.; Won, E.; Nam, K.; Ham, B.J. Impact of lingual gyrus volume on antidepressant response and neurocognitive functions in Major Depressive Disorder: A voxel-based morphometry study. J. Affect. Disord. 2014, 169, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qiao, L.; Chen, Q.; Yang, W.; Xu, M.; Yao, X.; Dong, Y.J. Gray Matter Volume of the Lingual Gyrus Mediates the Relationship between Inhibition Function and Divergent Thinking. Front. Psychol. 2016, 7, 1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Feng, Y.; Yang, M.; Chen, J.Y.; Li, J.; Huang, Z.C.; Zhang, L.L. Functional Connectivity in Patients With Sensorineural Hearing Loss Using Resting-State MRI. Am. J. Audiol. 2015, 24, 145. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Zhou, S.Y.; Nakamura, K.; Tanino, R.; Suzuki, M.J. Psychiatry, B. A follow-up MRI study of the fusiform gyrus and middle and inferior temporal gyri in schizophrenia spectrum. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 1957–1964. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Lin, H.; Liu, C.; Jiang, Y.; Lin, Y.; Xue, Q.; Xu, P.; Wang, Y. Transcranial Magnetic Stimulation to the Middle Frontal Gyrus During Attention Modes Induced Dynamic Module Reconfiguration in Brain Networks. Front. Neuroinform. 2019, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Arkin, S.C.; Ruiz-Betancourt, D.; Jamerson, E.C.; Smith, R.T.; Patel, G.H. Deficits and compensation: Attentional control cortical networks in schizophrenia. NeuroImage Clin. 2020, 27, 102348. [Google Scholar] [CrossRef] [PubMed]
- Juha, G.; Rasmus, Z.; Mikko, N.; Antti, P.; Synnve, C.J.C.C. Neural Substrate for Metacognitive Accuracy of Tactile Working Memory. Cereb. Cortex 2017, 11, 5343–5352. [Google Scholar]
- Maxim, K.; Alexander, K.; Natalia, M.; Ruslan, M.; Svyatoslav, M.J.F. Deceptive but Not Honest Manipulative Actions Are Associated with Increased Interaction between Middle and Inferior Frontal gyri. Front. Neurosci. 2017, 11, 482. [Google Scholar]
- Swick, D.; Ashley, V.; Turken, A.U.J.B.N. Left inferior frontal gyrus is critical for response inhibition. BMC Neurosci. 2008, 9, 102. [Google Scholar] [CrossRef] [Green Version]
- Kuperberg, G.R.; Broome, M.; Mcguire, P.K.; David, A.S.; Eddy, M.; Ozawa, F.; Kouwe, A.J.A. Regionally Localized Thinning of the Cerebral Cortex in Schizophrenia. Arch. Gen. Psychiatry 2003, 60, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulesu, E.; Frith, C.D.; Frackowiak, R.J.N. The neural correlates of the verbal component of working memory. Nature 1993, 362, 342–345. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, Y.W.; Shim, M.; Jin, M.J.; Lee, S.H.J.F. Altered Cortical Functional Networks in Patients With Schizophrenia and Bipolar Disorder: A Resting-State Electroencephalographic Study. Front. Psychiatry 2020, 11, 661. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Van Essen, D.C.; Raichle, M.E.J.P. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [Green Version]
- Whitfield-Gabrieli, S.; Ford, J.M. Default Mode Network Activity and Connectivity in Psychopathology, in Annual Review of Clinical Psychology, Vol 8, S. NolenHoeksema, Editor. Annu. Rev. Clin. Psychol. 2012, 8, 49–76. [Google Scholar] [CrossRef]
- Song, K.; Li, J.; Zhu, Y.; Ren, F.; Huang, Z. Altered small-world functional network topology in patients with optic neuritis: A resting-state fMRI study. Dis. Markers 2020, 2021, 9948751. [Google Scholar] [CrossRef]
- Ho, N.F.; Chong, P.; Lee, D.R.; Chew, Q.H.; Chen, G.; Sim, K.J.H.R. The Amygdala in Schizophrenia and Bipolar Disorder: A Synthesis of Structural MRI, Diffusion Tensor Imaging, and Resting-State Functional Connectivity Findings. Harv. Rev. Psychiatry 2019, 27, 150–164. [Google Scholar] [CrossRef]
- Katharina, S.; Stephan, B.; Tim, V.; Andrea, F.; Roland, W.; Müri, R.; Sebastian, W.J.S.B. Limbic Interference During Social Action Planning in Schizophrenia. Schizophr. Bull. 2018, 44, 359–368. [Google Scholar]
- Zhang, M.; Yang, F.; Fan, F.; Wang, Z.; Hong, L.E.J.N.C. Abnormal amygdala subregional-sensorimotor connectivity correlates with positive symptom in schizophrenia. NeuroImage Clin. 2020, 26, 102218. [Google Scholar] [CrossRef]
- Berman, R.A.; Gotts, S.J.; McAdams, H.M.; Greenstein, D.; Lalonde, F.; Clasen, L.; Clasen, L.; Watsky, R.E.; Shora, L.; Ordonez, A.E.; et al. Disrupted sensorimotor and social-cognitive networks underlie symptoms in childhood-onset schizophrenia. Brain 2016, 139, 276–291. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Yuan, B.; Cao, Q.; An, L.; Wang, P.; Vance, A.; Sun, L.J. Frequency-specific abnormalities in regional homogeneity among children with attention deficit hyperactivity disorder: A resting-state f MRI study. Sci. Bull. 2016, 61, 682–692. [Google Scholar] [CrossRef] [Green Version]
- Gohel, S.R.; Biswal, B.B. Functional integration between brain regions at rest occurs in multiple-frequency bands. Brain Connect. 2015, 5, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Ying-Hang, W.U.; Ping, N.I.; Li-Yuan, F.U.; Hui, L.I.; Chen, Z.Q. Resting-State Functional MRI Study on the Low-Frequency Fluctuation within Different Band of Amplitude of Alzheimer’s Disease. China Med. Devices 2014, 11, 5–10. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | SZ | NC | Statistical Test |
|---|---|---|---|
| Number of subjects | 42 | 40 | -- |
| Age (years) | 35.19 ± 8.37 | 32.25 ± 8.81 | P = 0.125 |
| Sex (male/female) | 30/12 | 25/15 | P = 0.396 |
| SAPS | 30.67 ± 22.26 | -- | -- |
| NC | SZ | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.01–0.25 Hz | Slow5 | Slow4 | Slow3 | Slow2 | 0.01–0.25 Hz | Slow5 | Slow4 | Slow3 | Slow2 |
| 5.2 | 6.4 | 6 | 7.4 | 4.6 | 5.8 | 7 | 4.8 | 5.4 | 4.6 |
| 7.6 | 5.4 | 7 | 6.6 | 7.2 | 6.6 | 6 | 6.8 | 7.6 | 5.4 |
| 6.6 | 6.2 | 6.2 | 5.8 | 6.2 | 5.2 | 7.6 | 6.2 | 6.4 | 4.8 |
| 6 | 8.6 | 6.2 | 5.4 | 5.2 | 6.4 | 7 | 7 | 5.6 | 6 |
| 7 | 7.8 | 6.8 | 7.6 | 6.2 | 6.2 | 6.2 | 6 | 6.4 | 4.6 |
| 6.6 | 5 | 4.6 | 6.8 | 5.2 | 6 | 6 | 6 | 6.8 | 4.4 |
| 7 | 7.8 | 7.4 | 7.8 | 6.8 | 6 | 6 | 5.2 | 7.8 | 5.8 |
| 5.8 | 4.8 | 6.4 | 8.2 | 6.8 | 5.2 | 7.6 | 7.4 | 5 | 4.8 |
| 5.8 | 7.4 | 6 | 7 | 5.8 | 5.6 | 5.4 | 5.4 | 6.2 | 7 |
| 7.4 | 7.6 | 7.4 | 6.4 | 6.4 | 7.2 | 7 | 6.4 | 7.8 | 6.8 |
| 6.2 | 6.8 | 7 | 7.2 | 5.4 | 6.6 | 4.2 | 6.4 | 6.2 | 4.8 |
| 6 | 7 | 5.4 | 6.6 | 4.8 | 5.2 | 6.4 | 3.6 | 6.8 | 5 |
| 7.4 | 6 | 7.2 | 8.2 | 5.6 | 6 | 5.8 | 5.2 | 5.8 | 6 |
| 7.4 | 7.4 | 7 | 5.8 | 7.8 | 6.4 | 5.2 | 6.2 | 7.6 | 5.6 |
| 6.8 | 6.2 | 8.2 | 6.2 | 5.8 | 7.4 | 6.6 | 7.2 | 9.4 | 7 |
| 7.2 | 6 | 7.2 | 4.8 | 6.4 | 6.4 | 6.4 | 7 | 7.2 | 5 |
| 7.4 | 7.2 | 6.8 | 7.2 | 7.6 | 8 | 5.6 | 7 | 9 | 4.8 |
| 7.8 | 8.6 | 7.6 | 5.4 | 7 | 6.6 | 6 | 5.6 | 5.4 | 7 |
| 6.4 | 6.6 | 8 | 6.2 | 6.2 | 6.4 | 6.8 | 6.8 | 7.6 | 5.2 |
| 6.2 | 6.8 | 6.2 | 7.8 | 5.2 | 7.2 | 8.6 | 6.8 | 7 | 4.2 |
| 6 | 5.8 | 5.6 | 6.4 | 6.8 | 7.6 | 5.2 | 7.2 | 6.4 | 6.4 |
| 7 | 5.8 | 5.2 | 7.4 | 6.2 | 5.6 | 5.2 | 4 | 6.4 | 5.4 |
| 7 | 5.2 | 5 | 4.8 | 5.2 | 6.2 | 6.2 | 7.4 | 8.2 | 6.2 |
| 5.8 | 7.4 | 7.2 | 5 | 7.6 | 7.8 | 6.4 | 6.6 | 6 | 6.2 |
| 6.2 | 7 | 5.2 | 7.6 | 7.2 | 5 | 6.6 | 5.4 | 5.8 | 6.6 |
| 5.4 | 5.8 | 5.4 | 7.4 | 6.2 | 6.6 | 7.6 | 6.8 | 8 | 4.8 |
| 7.8 | 7.2 | 7.4 | 7.4 | 4.4 | 7.8 | 7.2 | 6.8 | 6.2 | 4.6 |
| 6.8 | 6.8 | 6.4 | 7.6 | 6.8 | 6 | 5.8 | 5.6 | 7.4 | 6.8 |
| 7.6 | 6.6 | 7 | 5.8 | 5.4 | 6.2 | 5.2 | 5.6 | 6 | 6.8 |
| 5.6 | 6.6 | 8.2 | 6.6 | 4.8 | 6.8 | 7 | 7 | 6.4 | 6.6 |
| 6.6 | 6.4 | 7.2 | 7.4 | 7.4 | 5.6 | 6.4 | 4.6 | 9.2 | 4.6 |
| 5.6 | 7.6 | 5.4 | 6.6 | 6.2 | 6.2 | 7 | 8 | 6.4 | 4.6 |
| 7 | 6.8 | 5.4 | 6.2 | 5.2 | 7 | 7.4 | 5.2 | 5.4 | 5.2 |
| 6.2 | 7.6 | 6.6 | 7.2 | 6.4 | 6.2 | 7.2 | 6.4 | 7.2 | 6.8 |
| 7 | 7.8 | 7.4 | 7.6 | 7.2 | 6.2 | 7.4 | 7.6 | 5.4 | 5.4 |
| 6.4 | 6.2 | 4.8 | 5.2 | 5 | 6.6 | 7.4 | 7.2 | 7.6 | 6 |
| 6.6 | 9.6 | 7.8 | 8.4 | 6 | 7.4 | 7.4 | 6.8 | 6.8 | 4.6 |
| 7.2 | 8.6 | 8 | 7 | 5.6 | 6.8 | 6.8 | 7 | 7.8 | 4 |
| 6.8 | 7.2 | 8.2 | 6.6 | 6 | 6.8 | 6.6 | 7.4 | 7.8 | 4.6 |
| 5.6 | 7.8 | 6 | 5.8 | 7 | 6.8 | 8.2 | 6.4 | 7 | 4.4 |
| \ | \ | \ | \ | \ | 6.2 | 6.2 | 5 | 8.4 | 6.6 |
| \ | \ | \ | \ | \ | 8 | 6.8 | 8 | 8.2 | 5.6 |
| Characteristic | 0.01–0.25 Hz | Slow5 | Slow4 | Slow3 | Slow2 | |
|---|---|---|---|---|---|---|
| Integration | SMN | T = −1.277 P = 0.205 | T = 0.478 P = 0.634 | T = 0.171 P = 0.865 | T = −0.246 P = 0.807 | T = 0.073 P = 0.942 |
| VN | T = 1.012 P = 0.314 | T = 0.279 P = 0.781 | T = 2.263 P = 0.026 * | T = 0.522 P = 0.603 | T = 0.522 P = 0.603 | |
| AN | T = −0.781 P = 0.437 | T = 0.639 P = 0.525 | T = 0.378 P = 0.707 | T = 0.109 P = 0.914 | T = 1.124 P = 0.264 | |
| DMN | T = −1.010 P = 0.316 | T = 0.455 P = 0.651 | T = 0.048 P = 0.962 | T = −0.564 P = 0.574 | T = 1.394 P = 0.167 | |
| LSN | T = −1.610 P = 0.111 | T = −0.041 P = 0.967 | T = 1.457 P = 0.149 | T = −0.576 P = 0.566 | T = 1.031 P = 0.305 | |
| Recruitment | SMN | T = −0.783 P = 0.436 | T = 1.761 P = 0.082 | T = −0.677 P = 0.501 | T = −0.196 P = 0.845 | T = 0.889 P = 0.377 |
| VN | T = −2.840 P = 0.006 ** | T = −0.701 P = 0.485 | T = −4.101 P = 0.000 *** | T = −0.830 P = 0.409 | T = −0.272 P = 0.787 | |
| AN | T = −2.392 P = 0.019 * | T = 0.07 P = 0.945 | T = −3.557 P = 0.001 *** | T = −1.266 P = 0.209 | T = 0.195 P = 0.846 | |
| DMN | T = −1.123 P = 0.265 | T = 1.352 P = 0.180 | T = −2.178 P = 0.032 * | T = 0.390 P = 0.698 | T = −0.803 P = 0.424 | |
| LSN | T = −0.713 P = 0.478 | T = 1.489 P = 0.140 | T = 2.456 P = 0.016 * | T = −0.431 P = 0.668 | T = 0.163 P = 0.871 | |
| RSN1 | RSN2 | NC (SD) | SZ (SD) | P (FDR) |
|---|---|---|---|---|
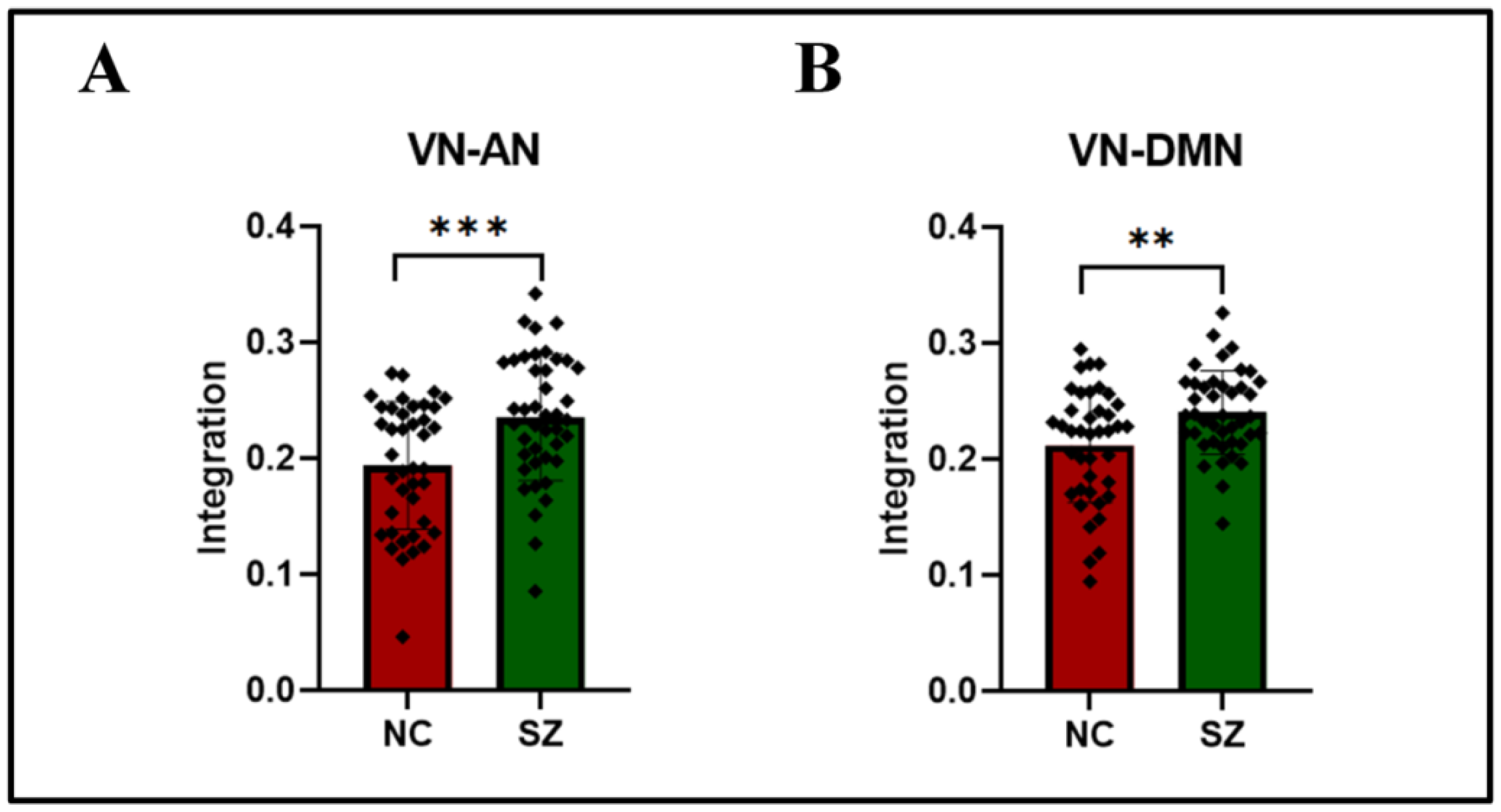
| Visual network (VN) | Attention network (AN) | 0.194 (0.055) | 0.235 (0.055) | 0.000 |
| Visual network (VN) | Default mode network (DMN) | 0.212 (0.049) | 0.240 (0.036) | 0.001 |
| Frequency | RSN1 | RSN2 | NC (SD) | SZ (SD) | P (FDR) |
|---|---|---|---|---|---|
| 0.01–0.25 Hz | visual network (VN) | somatosenery/motor and auditory network (SMN) | 0.258 (0.115) | 0.267 (0.132) | 0.731 |
| visual network (VN) | attention network (AN) | 0.168 (0.082) | 0.183 (0.062) | 0.367 | |
| visual network (VN) | default mode network (DMN) | 0.201 (0.071) | 0.213 (0.055) | 0.774 | |
| visual network (VN) | limbic/paralimbic and subcortical network (LSN) | 0.165 (0.069) | 0.180 (0.056) | 0.312 | |
| Slow5 | visual network (VN) | somatosenery/motor and auditory network (SMN) | 0.348 (0.110) | 0.344 (0.126) | 0.858 |
| visual network (VN) | attention network (AN) | 0.292 (0.088) | 0.295 (0.069) | 0.839 | |
| visual network (VN) | default mode network (DMN) | 0.321 (0.078) | 0.329 (0.068) | 0.639 | |
| visual network (VN) | limbic/paralimbic and subcortical network (LSN) | 0.294 (0.067) | 0.307 (0.089) | 0.469 | |
| Slow4 | visual network (VN) | somatosenery/motor and auditory network (SMN) | 0.311 (0.119) | 0.299 (0.108) | 0.613 |
| visual network (VN) | attention network (AN) | 0.210 (0.075) | 0.215 (0.071) | 0.706 | |
| visual network (VN) | default mode network (DMN) | 0.254 (0.069) | 0.260 (0.069) | 0.666 | |
| visual network (VN) | limbic/paralimbic and subcortical network (LSN) | 0.232 (0068) | 0.236 (0.071) | 0.774 | |
| Slow3 | visual network (VN) | somatosenery/motor and auditory network (SMN) | 0.271 (0.074) | 0.271 (0.071) | 0.552 |
| visual network (VN) | attention network (AN) | 0.194 (0.055) | 0.235 (0.055) | 0.000 | |
| visual network (VN) | default mode network (DMN) | 0.212 (0.049) | 0.240 (0.036) | 0.001 | |
| visual network (VN) | limbic/paralimbic and subcortical network (LSN) | 0.220 (0.052) | 0.215 (0.043) | 0.797 | |
| Slow2 | visual network (VN) | somatosenery/motor and auditory network (SMN) | 0.313 (0.055) | 0.311 (0.061) | 0.856 |
| visual network (VN) | attention network (AN) | 0.294 (0.035) | 0.297 (0.054) | 0.759 | |
| visual network (VN) | default mode network (DMN) | 0.284 (0.041) | 0.290 (0.044) | 0.458 | |
| visual network (VN) | limbic/paralimbic and subcortical network (LSN) | 0.269 (0.042) | 0.280 (0.051) | 0.271 |
| Frequency | Network | Name | Abb | ROI | NC (SD) | SZ (SD) | P (FDR) |
|---|---|---|---|---|---|---|---|
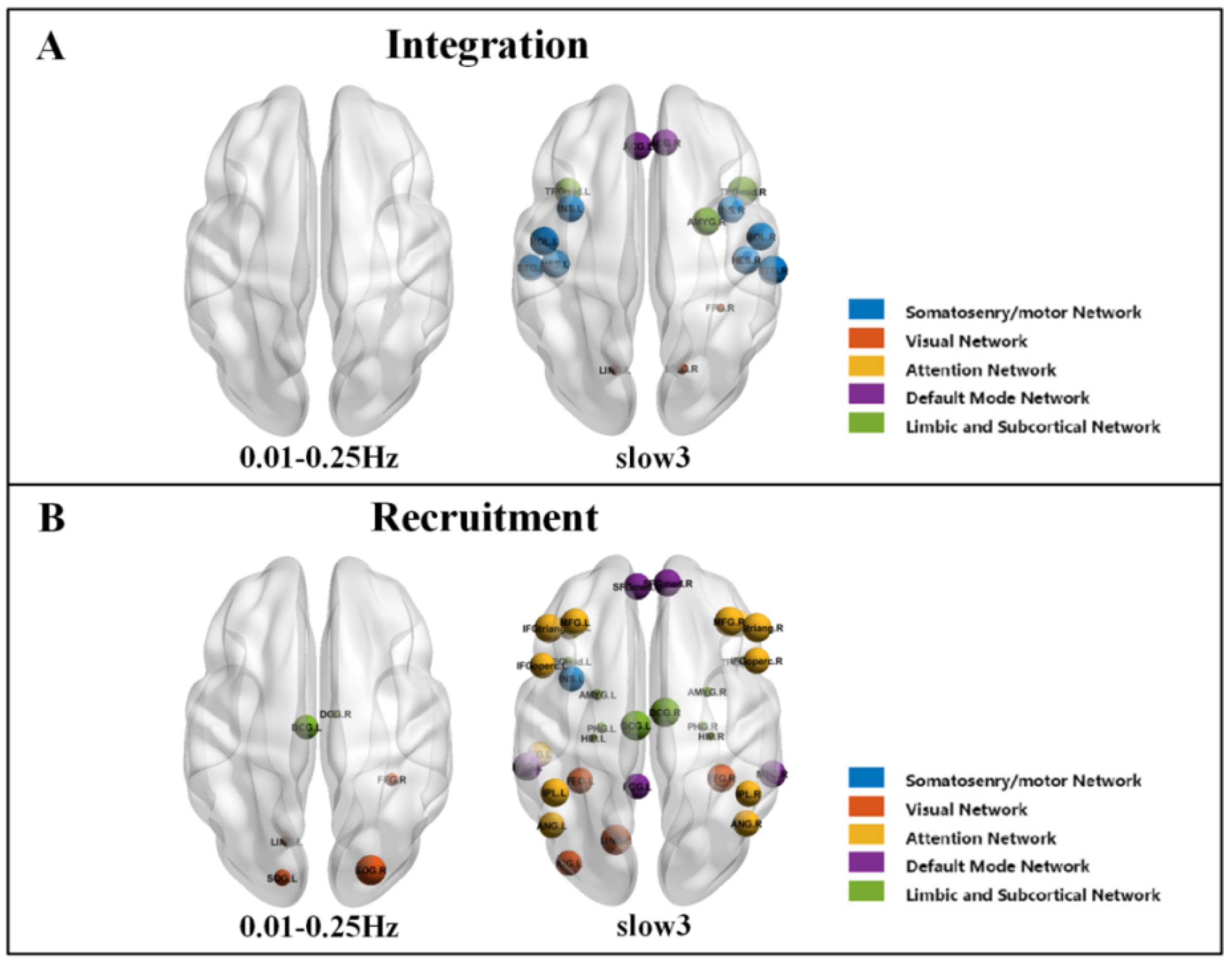
| Slow3 | SMN | Rolandic_Oper | ROL.L | 17 | 0.289 (0.053) | 0.245 (0.053) | 0.001 |
| ROL.R | 18 | 0.279 (0.041) | 0.243 (0.053) | 0.011 | |||
| Insula | INS.L | 29 | 0.300 (0.055) | 0.258 (0.062) | 0.015 | ||
| INS.R | 30 | 0.300 (0.055) | 0.260 (0.060) | 0.015 | |||
| Heschl | HES.L | 79 | 0.283 (0.049) | 0.244 (0.052) | 0.011 | ||
| HES.R | 80 | 0.280 (0.044) | 0.244 (0.048) | 0.011 | |||
| Temporal_Sup | STG.L | 81 | 0.286 (0.051) | 0.253 (0.055) | 0.036 | ||
| STG.R | 82 | 0.284 (0.049) | 0.243 (0.047) | 0.000 | |||
| VN | Lingual | LING.L | 47 | 0.212 (0.053) | 0.248 (0.050) | 0.015 | |
| LING.R | 48 | 0.208 (0.052) | 0.239 (0.054) | 0.045 | |||
| Fusiform_R | FFG.R | 56 | 0.241 (0.058) | 0.288 (0.047) | 0.000 | ||
| DMN | Cingulum_Ant | ACG.L | 31 | 0.297 (0.052) | 0.260 (0.052) | 0.015 | |
| ACG.R | 32 | 0.297 (0.054) | 0.263 (0.052) | 0.032 | |||
| LSN | Amygdala_R | AMYG.R | 42 | 0.272 (0.056) | 0.233 (0.046) | 0.011 | |
| Temporal_Pole_Mid | TPOmid.L | 87 | 0.280 (0.052) | 0.248 (0.046) | 0.028 | ||
| TPOmid.R | 88 | 0.294 (0.058) | 0.247 (0.041) | 0.000 |
| Frequency | Network | Name | Abb | ROI | NC (SD) | SZ (SD) | P (FDR) |
|---|---|---|---|---|---|---|---|
| 0.01–0.25 Hz | VN | Lingual_L | LING.L | 47 | 0.743 (0.143) | 0.634 (0.169) | 0.037 |
| Occipital_Sup | SOG.L | 49 | 0.769 (0.114) | 0.676 (0.145) | 0.037 | ||
| SOG.R | 50 | 0.769 (0.118) | 0.661 (0.158) | 0.037 | |||
| Fusiform_R | FFG.R | 56 | 0.534 (0.247) | 0.371 (0.219) | 0.037 | ||
| LSN | Cingulum_Mid | DCG.L | 33 | 0.287 (0.131) | 0.200 (0.097) | 0.037 | |
| DCG.R | 34 | 0.281 (0.122) | 0.199 (0.106) | 0.037 | |||
| Slow3 | SMN | Insula_L | INS.L | 29 | 0.456 (0.116) | 0.395 (0.110) | 0.048 |
| VN | Lingual_L | LING.L | 47 | 0.610 (0.129) | 0.477 (0.143) | 0.001 | |
| Occipital_Inf_L | IOG.L | 53 | 0.494 (0.157) | 0.405 (1.173) | 0.048 | ||
| Fusiform | FFG.L | 55 | 0.312 (0.166) | 0.232 (0.112) | 0.040 | ||
| FFG.L | 56 | 0.349 (0.176) | 0.233 (0.115) | 0.004 | |||
| AN | Frontal_Mid | MFG.L | 7 | 0.421 (0.960) | 0.355 (0.093) | 0.010 | |
| MFG.R | 8 | 0.429 (0.088) | 0.357 (0.075) | 0.002 | |||
| Frontal_Inf_Oper | IFGoperc.L | 11 | 0.384 (0.112) | 0.328 (0.093) | 0.048 | ||
| IFGoperc.R | 12 | 0.375 (0.107) | 0.317 (0.086) | 0.031 | |||
| Frontal_Inf_Tri | IFGtriang.L | 13 | 0.434 (0.091) | 0.357 (0.098) | 0.003 | ||
| IFGtriang.R | 14 | 0.423 (0.095) | 0.345 (0.086) | 0.002 | |||
| Frontal_Inf_Orb | ORBinf.L | 15 | 0.394 (0.085) | 0.346 (0.088) | 0.048 | ||
| ORBinf.R | 16 | 0.399 (0.101) | 0.334 (0.082) | 0.011 | |||
| Parietal_Inf | IPL.L | 61 | 0.365 (0.101) | 0.297 (0.068) | 0.004 | ||
| IPL.R | 62 | 0.369 (0.106) | 0.317 (0.067) | 0.034 | |||
| Angular | ANG.L | 65 | 0.365 (0.101) | 0.313 (0.074) | 0.034 | ||
| ANG.R | 66 | 0.372 (0.099) | 0.316 (0.082) | 0.023 | |||
| Temporal_Inf_L | ITG.L | 89 | 0.309 (0.093) | 0.260 (0.089) | 0.048 | ||
| DMN | Frontal_Sup_Medial | SFGmed.L | 23 | 0.442 (0.096) | 0.385 (0.078) | 0.017 | |
| SFGmed.R | 24 | 0.446 (0.091) | 0.387 (0.081) | 0.011 | |||
| Cingulum_Post_L | PCG.L | 35 | 0.340 (0.111) | 0.287 (0.081) | 0.048 | ||
| Temporal_Mid | MTG.L | 85 | 0.341 (0.108) | 0.268 (0.076) | 0.004 | ||
| MTG.R | 86 | 0.324 (0.112) | 0.258 (0.072) | 0.010 | |||
| LSN | Cingulum_Mid | DCG.L | 33 | 0.270 (0.107) | 0.184 (0.074) | 0.001 | |
| DCG.R | 34 | 0.267 (0.101) | 0.189 (0.073) | 0.002 | |||
| Hippocampus | HIP.L | 37 | 0.396 (0.075) | 0.479 (0.099) | 0.001 | ||
| HIP.R | 38 | 0.389 (0.080) | 0.483 (0.105) | 0.001 | |||
| ParaHippocampal | PHG.L | 39 | 0.395 (0.071) | 0.468 (0.103) | 0.003 | ||
| PHG.R | 40 | 0.389 (0.074) | 0.470 (0.106) | 0.002 | |||
| Amygdala | AMYG.L | 41 | 0.392 (0.082) | 0.466 (0.108) | 0.005 | ||
| AMYG.R | 42 | 0.386 (0.089) | 0.469 (0.108) | 0.003 | |||
| Temporal_Pole_Mid | TPOmid.L | 87 | 0.324 (0.085) | 0.423 (0.131) | 0.002 | ||
| TPOmid.R | 88 | 0.336 (0.099) | 0.409 (0.123) | 0.017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhang, Y.; Xiang, J.; Wang, B.; Li, D.; Cheng, X.; Liu, T.; Cui, X. Frequency-Specific Analysis of the Dynamic Reconfiguration of the Brain in Patients with Schizophrenia. Brain Sci. 2022, 12, 727. https://doi.org/10.3390/brainsci12060727
Yang Y, Zhang Y, Xiang J, Wang B, Li D, Cheng X, Liu T, Cui X. Frequency-Specific Analysis of the Dynamic Reconfiguration of the Brain in Patients with Schizophrenia. Brain Sciences. 2022; 12(6):727. https://doi.org/10.3390/brainsci12060727
Chicago/Turabian StyleYang, Yanli, Yang Zhang, Jie Xiang, Bin Wang, Dandan Li, Xueting Cheng, Tao Liu, and Xiaohong Cui. 2022. "Frequency-Specific Analysis of the Dynamic Reconfiguration of the Brain in Patients with Schizophrenia" Brain Sciences 12, no. 6: 727. https://doi.org/10.3390/brainsci12060727
APA StyleYang, Y., Zhang, Y., Xiang, J., Wang, B., Li, D., Cheng, X., Liu, T., & Cui, X. (2022). Frequency-Specific Analysis of the Dynamic Reconfiguration of the Brain in Patients with Schizophrenia. Brain Sciences, 12(6), 727. https://doi.org/10.3390/brainsci12060727