
Relationship between Tactile Sensation, Motor Activity, and Differential Brain Activity in Young Individuals



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
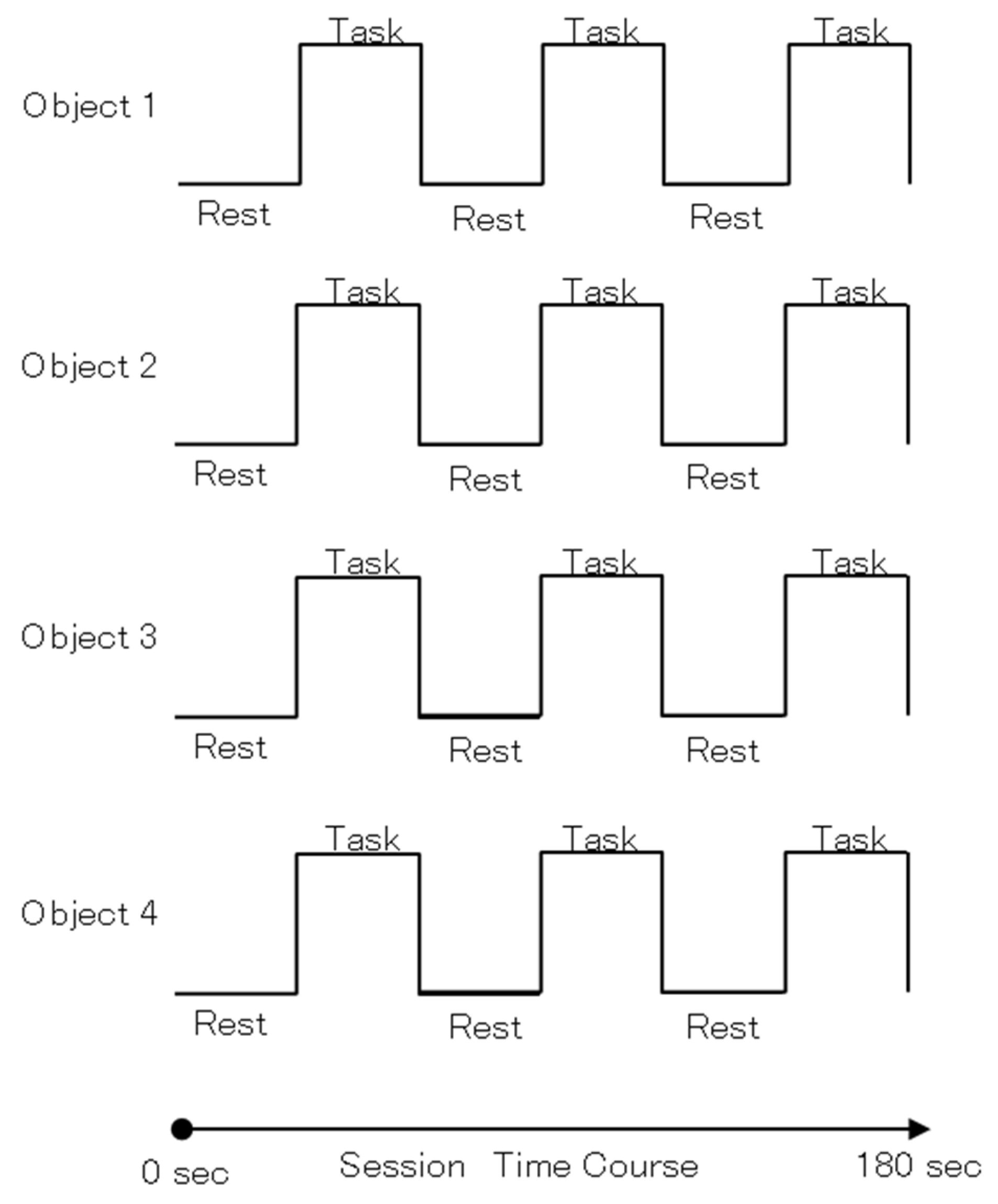
2.2. Stimuli Task
2.3. Apparatus
2.4. MRI Acquisition
2.5. fMRI Data Analyses
2.6. Questionnaire
2.7. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Willoughby, W.R.; Thoenes, K.; Bolding, M. Somatotopic arrangement of the human primary somatosensory cortex derived from functional magnetic resonance imaging. Front. Neurosci. 2021, 14, 598482. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Han, H.; Chui, D.; Shen, Y.; Wu, J. Prominent activation of the intraparietal and somatosensory areas during angle discrimination by intra-active touch. Hum. Brain Mapp. 2012, 33, 2957–2970. [Google Scholar] [CrossRef] [PubMed]
- Ackerley, R.; Hassan, E.; Curran, A.; Wessberg, J.; Olausson, H. An fMRI study on cortical responses during active self-touch and passive touch from others. Front. Behav. Neurosci. 2012, 6, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, H.; Custead, R.; Wang, Y.; Barlow, S. Neural encoding of saltatory pneumotactile velocity in human glabrous hand. PLoS ONE 2017, 12, e0183532. [Google Scholar] [CrossRef] [Green Version]
- Bola, L.; Siuda-Krzywicka, K.; Paplinska, M.; Sumera, E.; Zimmerman, M.; Jednorog, K.; Marchewka, A.; Szwed, M. Structural reorganization of the early visual cortex following Braille training in sighted adults. Sci. Rep. 2017, 7, 17448. [Google Scholar] [CrossRef] [Green Version]
- Landelle, C.; Anton, J.L.; Nazarian, B.; Sein, J.; Gharbi, A.; Felician, O.; Kavounoudias, A. Functional brain changes in the elderly for the perception of hand movements: A greater impairment occurs in proprioception than touch. Neuroimage 2020, 220, 117056. [Google Scholar] [CrossRef]
- Lee, I.S.; Jung, W.M.; Park, H.J.; Chae, Y. Spatial information of somatosensory stimuli in the brain: Multivariate pattern analysis of functional magnetic resonance imaging data. Neural Plast. 2020, 2020, 8307580. [Google Scholar] [CrossRef]
- Härtner, J.; Strauss, S.; Pfannmoller, J.; Lotze, M. Tactile acuity of fingertips and hand representation size in human Area 3b and Area 1 of the primary somatosensory cortex. Neuroimage 2021, 232, 117912. [Google Scholar] [CrossRef]
- Liu, P.; Chrysidou, A.; Doehler, J.; Hebart, M.N.; Wolbers, T.; Kuehn, E. The organizational principles of de-differentiated topographic maps in somatosensory cortex. Elife 2021, 10, e60090. [Google Scholar] [CrossRef]
- Schmidt, T.T.; Schroder, P.; Reinhardt, P.; Blankenburg, F. Rehearsal of tactile working memory: Premotor cortex recruits two dissociable neuronal content representations. Hum. Brain Mapp. 2021, 42, 245–258. [Google Scholar] [CrossRef]
- Velenosi, L.A.; Wu, Y.H.; Schmidt, T.T.; Blankenburg, F. Intraparietal sulcus maintains working memory representations of somatosensory categories in an adaptive, context-dependent manner. Neuroimage 2020, 221, 117146. [Google Scholar] [CrossRef]
- Kilteni, K.; Ehrsson, H.H. Functional connectivity between the cerebellum and somatosensory areas implements the attenuation of self-generated touch. J. Neurosci. 2020, 40, 894–906. [Google Scholar] [CrossRef]
- Masson, H.L.; Van De Plas, S.; Daniels, N.; Op de Beeck, H. The multidimensional representational space of observed socio-affective touch experiences. Neuroimage 2018, 175, 297–314. [Google Scholar] [CrossRef]
- Yang, J.; Molfese, P.J.; Yu, Y.; Handwerker, D.A.; Chen, G.; Taylor, P.A.; Ejima, Y.; Wu, J.; Bandettini, P.A. Different activation signatures in the primary sensorimotor and higher-level regions for haptic three-dimensional curved surface exploration. Neuroimage 2021, 231, 117754. [Google Scholar] [CrossRef]
- Woolgar, A.; Zopf, R. Multisensory coding in the multiple-demand regions: Vibrotactile task information is coded in frontoparietal cortex. J. Neurophysiol. 2017, 118, 703–716. [Google Scholar] [CrossRef] [Green Version]
- Federico, G.; Osiurak, F.; Reynaud, E.; Brandimonte, M.A. Semantic congruency effects of prime words on tool visual exploration. Brain Cogn. 2021, 152, 105758. [Google Scholar] [CrossRef]
- Buetefisch, C.M.; Revill, K.P.; Shuster, L.; Hines, B.; Parsons, M. Motor demand-dependent activation of ipsilateral motor cortex. J. Neurophysiol. 2014, 112, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Kunz, M.; Chen, J.I.; Rainville, P. Keeping an eye on pain expression in primary somatosensory cortex. Neuroimage 2020, 217, 116885. [Google Scholar] [CrossRef]
- Pando-Naude, V.; Barrios, F.A.; Alcauter, S.; Pasaye, E.H.; Vase, L.; Brattico, E.; Vuust, P.; Garza-Villarreal, E.A. Functional connectivity of music-induced analgesia in fibromyalgia. Sci. Rep. 2019, 9, 15486. [Google Scholar] [CrossRef] [Green Version]
- Servos, P.; Lederman, S.; Wilson, D.; Gati, J. fMRI-derived cortical maps for haptic shape, texture, and hardness. Brain Res. Cogn. Brain Res. 2001, 12, 307–313. [Google Scholar] [CrossRef]
- Harrewijn, A.; Cardinale, E.M.; Groenewold, N.A.; Bas-Hoogendam, J.M.; Aghajani, M.; Hilbert, K.; Cardoner, N.; Porta-Casteras, D.; Gosnell, S.; Salas, R.; et al. Cortical and subcortical brain structure in generalized anxiety disorder: Findings from 28 research sites in the ENIGMA-Anxiety Working Group. Transl. Psychiatry 2021, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Wacker, E.; Spitzer, B.; Lutzkendorf, R.; Bernarding, J.; Blackenburg, F. Tactile motion and pattern processing assessed with high-field FMRI. PLoS ONE 2011, 6, e24860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, R.; Pobric, G.; Saito, S.; Lambon Ralph, M.A. The neural network for tool-related cognition: An activation likelihood estimation meta-analysis of 70 neuroimaging contrasts. Cogn. Neuropsychol. 2016, 33, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Pupíková, M.; Simko, P.; Gajdos, M.; Rektorova, I. Modulation of working memory and resting-state fMRI by tDCS of the right frontoparietal network. Neural Plast. 2021, 2021, 5594305. [Google Scholar] [CrossRef]
- Federico, G.; Osiurak, F.; Brandimonte, M.A. Hazardous tools: The emergence of reasoning in human tool use. Psychol. Res. 2021, 85, 3108–3118. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Completely Disagree | Disagree | Undecided | Agree | Strongly Agree | ||
|---|---|---|---|---|---|---|
| Object 1 | Comfort | 17 | 3 | 2 | 3 | 0 |
| Hard | 0 | 0 | 0 | 3 | 22 | |
| Pain | 0 | 0 | 0 | 5 | 20 | |
| Easy to grasp | 5 | 7 | 3 | 10 | 0 | |
| Object 2 | Comfort | 2 | 4 | 1 | 5 | 13 |
| Hard | 17 | 5 | 0 | 2 | 1 | |
| Pain | 23 | 1 | 0 | 0 | 1 | |
| Easy to grasp | 0 | 1 | 1 | 8 | 15 | |
| Object 3 | Comfort | 6 | 9 | 6 | 3 | 1 |
| Hard | 0 | 2 | 0 | 11 | 12 | |
| Pain | 19 | 6 | 0 | 0 | 0 | |
| Easy to grasp | 5 | 4 | 4 | 8 | 4 | |
| Object 4 | Comfort | 7 | 7 | 1 | 7 | 3 |
| Hard | 25 | 0 | 0 | 0 | 0 | |
| Pain | 25 | 0 | 0 | 0 | 0 | |
| Easy to grasp | 18 | 6 | 1 | 0 | 0 |
| Cluster Size (voxels) | Cluster p-Value (FWE) | T-Value | Z-Score | x {mm} | y {mm} | z {mm} | Hemisphere | Locations | |
|---|---|---|---|---|---|---|---|---|---|
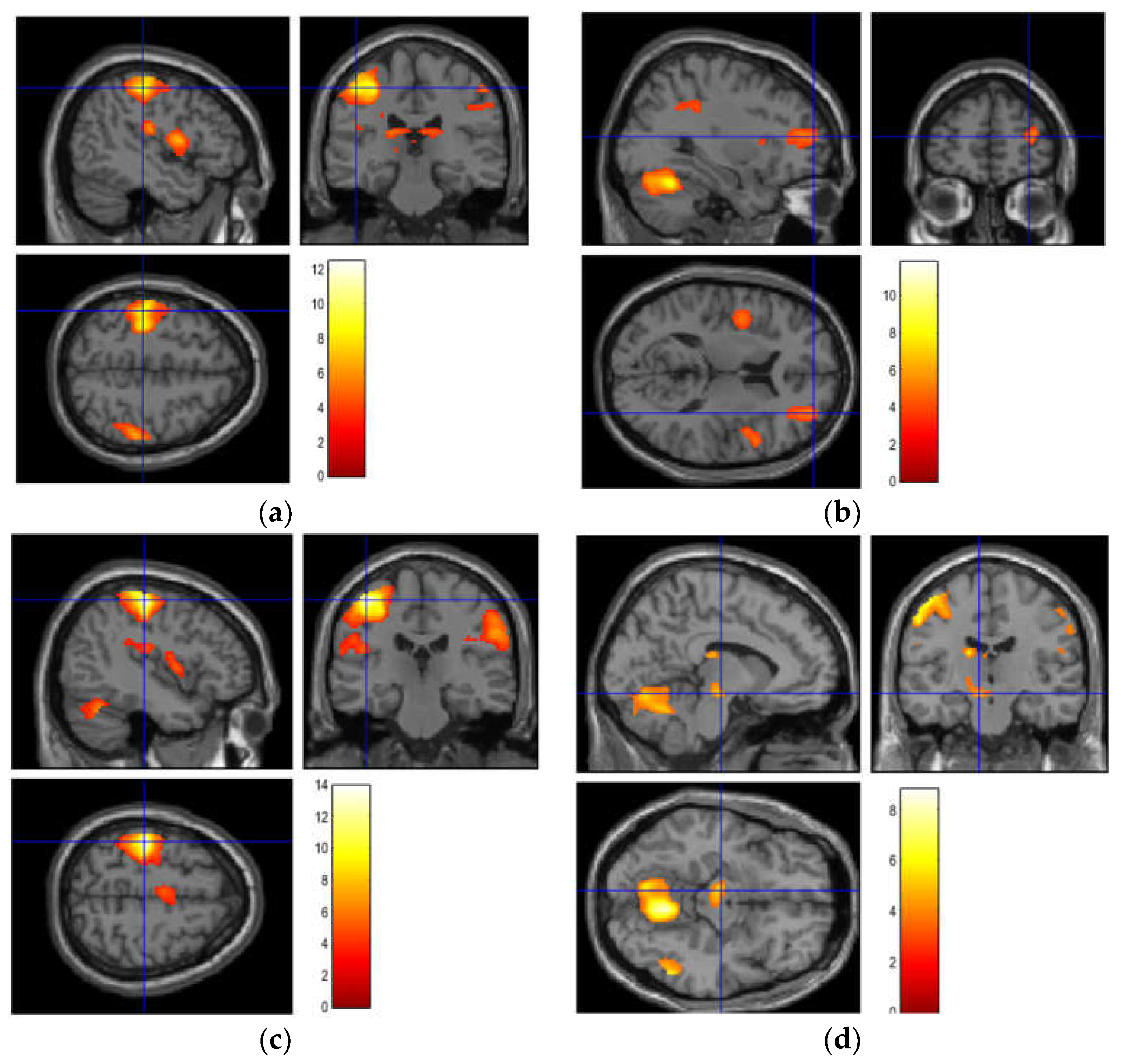
| Object 1 | 3797 | <0.001 | 12.43 | 6.96 | 22 | −52 | −26 | Right | Lateral hemisphere of the cerebellum |
| 1989 | <0.001 | 9.18 | 6.02 | −46 | −26 | 52 | Left | Postcentral gyrus | |
| 502 | 0.004 | 6.97 | 5.15 | −20 | −30 | 16 | Left | Thalamus proper | |
| 827 | <0.001 | 6.62 | 4.98 | −46 | 4 | 10 | Left | Precentral gyrus | |
| 539 | 0.003 | 6.39 | 4.88 | 52 | −32 | 52 | Right | Supramarginal gyrus | |
| 394 | 0.012 | 5.59 | 4.46 | 26 | 40 | 16 | Right | Middle frontal gyrus | |
| 635 | 0.001 | 5.49 | 4.41 | 48 | 6 | 2 | Right | Anterior insula | |
| Object 2 | 5231 | <0.001 | 11.77 | 6.79 | 20 | −56 | −22 | Right | Lateral hemisphere of the cerebellum |
| 2557 | <0.001 | 9.53 | 6.14 | −36 | −28 | 46 | Left | Precentral gyrus | |
| 2520 | <0.001 | 7.18 | 5.24 | 56 | −18 | 40 | Right | Supramarginal gyrus | |
| 715 | <0.001 | 5.7 | 4.52 | 58 | 10 | 24 | Right | Precentral gyrus | |
| 337 | 0.019 | 5.14 | 4.2 | 32 | 54 | 12 | Right | Middle frontal gyrus | |
| Object 3 | 2642 | <0.001 | 13.91 | 7.3 | −42 | −24 | 58 | Left | Postcentral gyrus |
| 4589 | <0.001 | 9.34 | 6.08 | 18 | −54 | −22 | Right | Lateral hemisphere of the cerebellum | |
| 2219 | <0.001 | 7.91 | 5.55 | 60 | −18 | 36 | Right | Supramarginal gyrus | |
| 397 | 0.014 | 5.13 | 4.2 | −44 | 0 | 8 | Left | Anterior insula | |
| 378 | 0.017 | 5.11 | 4.19 | −4 | −10 | 54 | Left | Supplementary motor cortex | |
| Object 4 | 3729 | <0.001 | 8.78 | 5.88 | 16 | −54 | −20 | Right | Lateral hemisphere of the cerebellum |
| 2140 | <0.001 | 7.5 | 5.38 | −52 | −20 | 48 | Left | Postcentral gyrus | |
| 395 | 0.004 | 7.4 | 5.34 | −18 | −20 | 18 | Left | Thalamus proper | |
| 281 | 0.019 | 6.44 | 4.9 | 56 | −56 | −10 | Right | Inferior temporal gyrus | |
| 1417 | <0.001 | 6.16 | 4.76 | 54 | −24 | 48 | Right | Supramarginal gyrus | |
| 222 | 0.044 | 5.69 | 4.52 | −8 | −16 | −12 | Left | Ventral diencephalon |
| Object 1 | Object 2 | Object 3 | Object 4 | F-Value | p-Value | |
|---|---|---|---|---|---|---|
| Precentral gyrus | 0.358 ± 0.483 | 0.446 ± 0.424 | 0.234 ± 0.369 | 0.221 ± 0.380 | 1.716 | 0.169 |
| Postcentral gyrus | 1.124 ± 0.363 | 1.251 ± 0.782 | 1.128 ± 0.415 | 0.790 ± 0.593 | 2.640 | 0.054 |
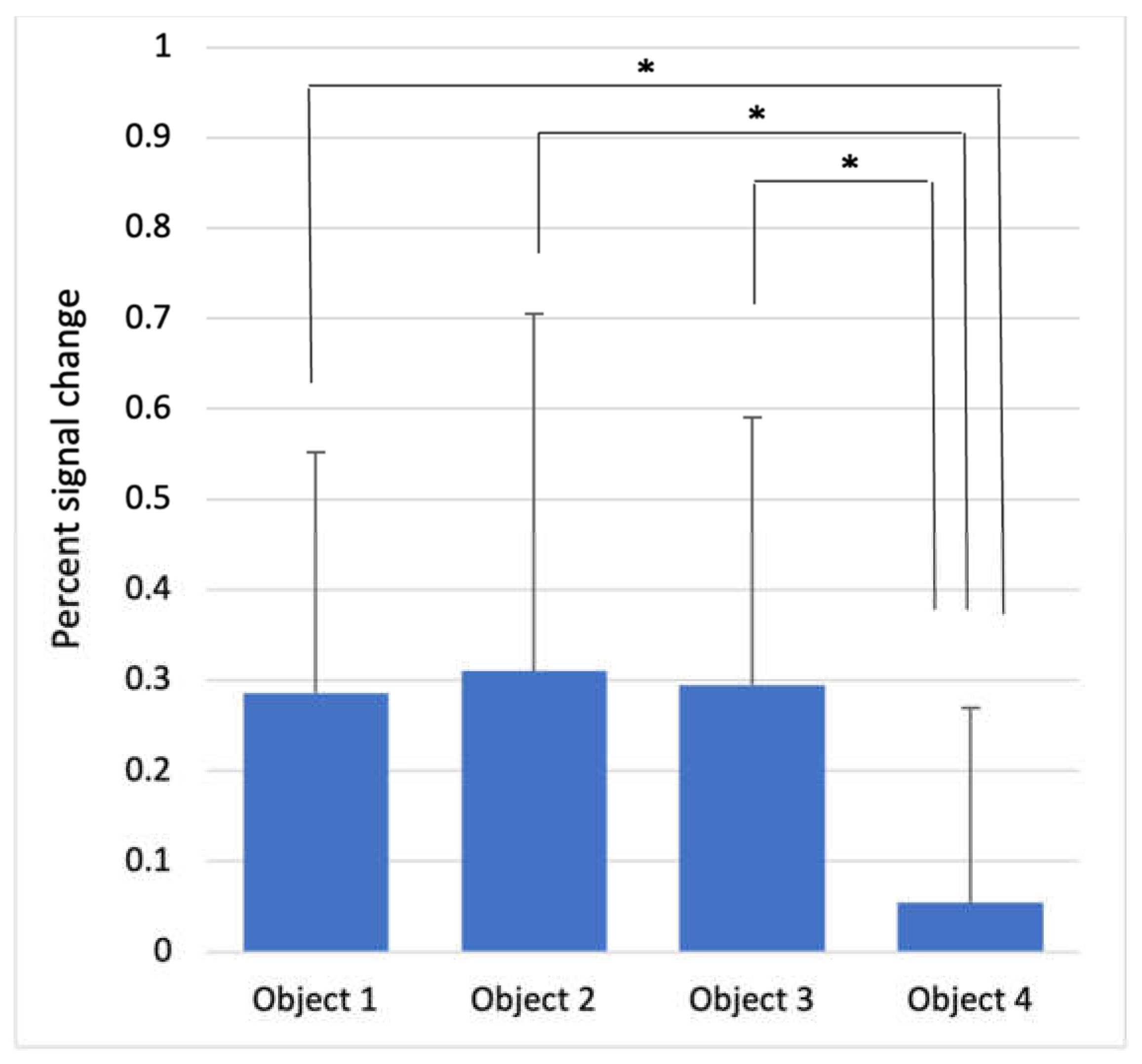
| Anterior insula | 0.286 ± 0.266 | 0.310 ± 0.395 | 0.295 ± 0.296 | 0.054 ± 0.215 | 4.278 | <0.001 |
| Lateral hemisphere of the cerebellum | 0.801 ± 0.401 | 0.968 ± 0.392 | 0.874 ± 0.527 | 0.645 ± 0.438 | 2.490 | 0.065 |
| Ventral diencephalon | 0.028 ± 0.252 | 0.106 ± 0.335 | 0.100 ± 0.289 | 0.206 ± 0.215 | 1.834 | 0.146 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, R.; Sakurai, N.; Nagasaka, K.; Kasai, S.; Kodama, N. Relationship between Tactile Sensation, Motor Activity, and Differential Brain Activity in Young Individuals. Brain Sci. 2022, 12, 924. https://doi.org/10.3390/brainsci12070924
Kobayashi R, Sakurai N, Nagasaka K, Kasai S, Kodama N. Relationship between Tactile Sensation, Motor Activity, and Differential Brain Activity in Young Individuals. Brain Sciences. 2022; 12(7):924. https://doi.org/10.3390/brainsci12070924
Chicago/Turabian StyleKobayashi, Ryota, Noriko Sakurai, Kazuaki Nagasaka, Satoshi Kasai, and Naoki Kodama. 2022. "Relationship between Tactile Sensation, Motor Activity, and Differential Brain Activity in Young Individuals" Brain Sciences 12, no. 7: 924. https://doi.org/10.3390/brainsci12070924