The Impact of Chronic Stress on Behavior and Body Mass in New Animal Models

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Description of the Animal Models Used in the Study
2.3. Experimental Design
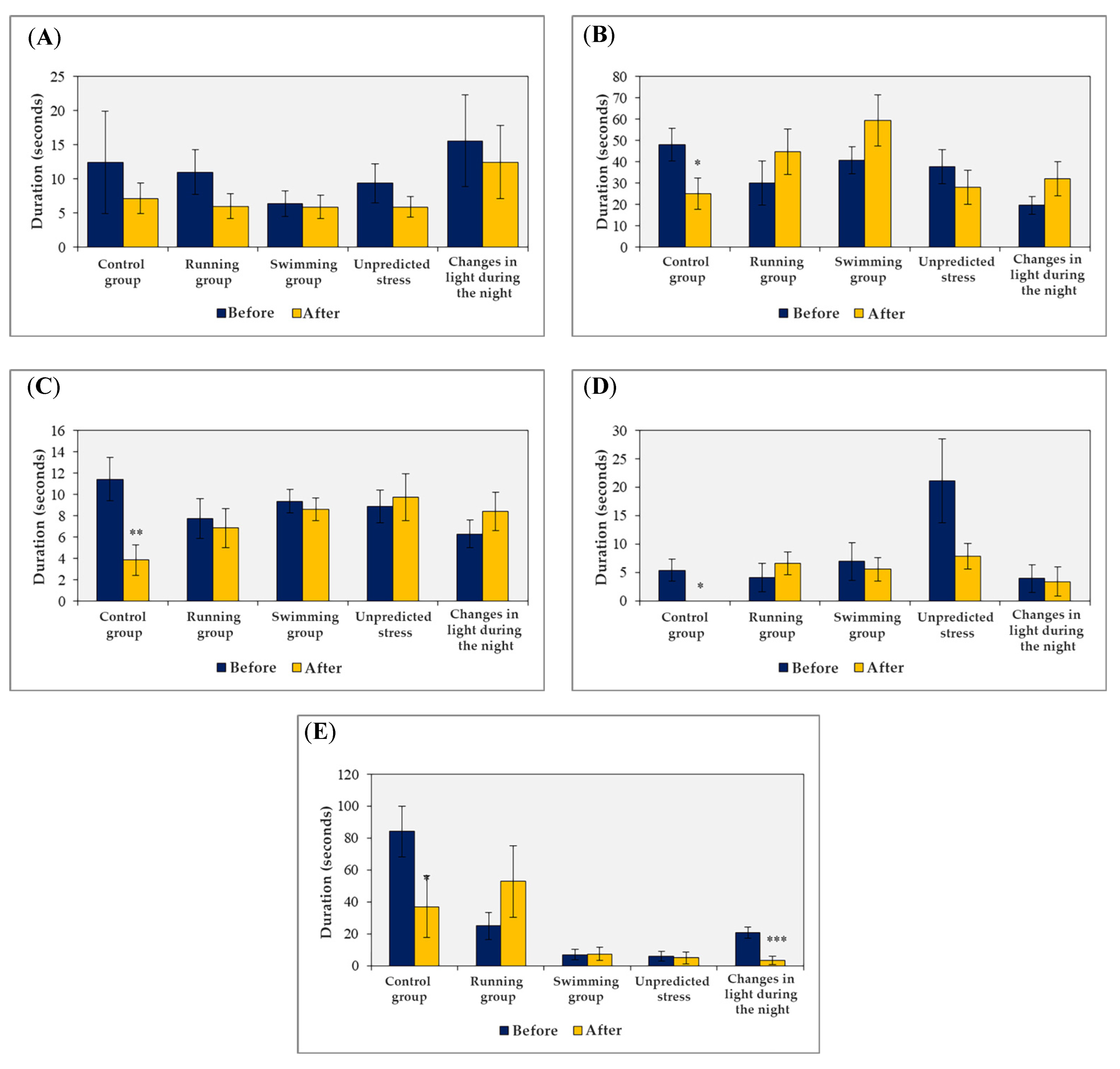
3. Results
3.1. The Effects of Exposure to External Stress Factors on the Behavior of Animals in the Open Field Test
- -
- degree of disinhibition, which corresponds to the amount of time spent in the central square of the maze;
- -
- general mobility, which corresponds to the number of crossings of the lateral squares;
- -
- anxious behavior, which corresponds to the frequency of stretching behavior in the horizontal plane and the duration of freezing or immobility;
- -
- anxiolytic behavior, which corresponds to the duration of fur cleaning behavior.
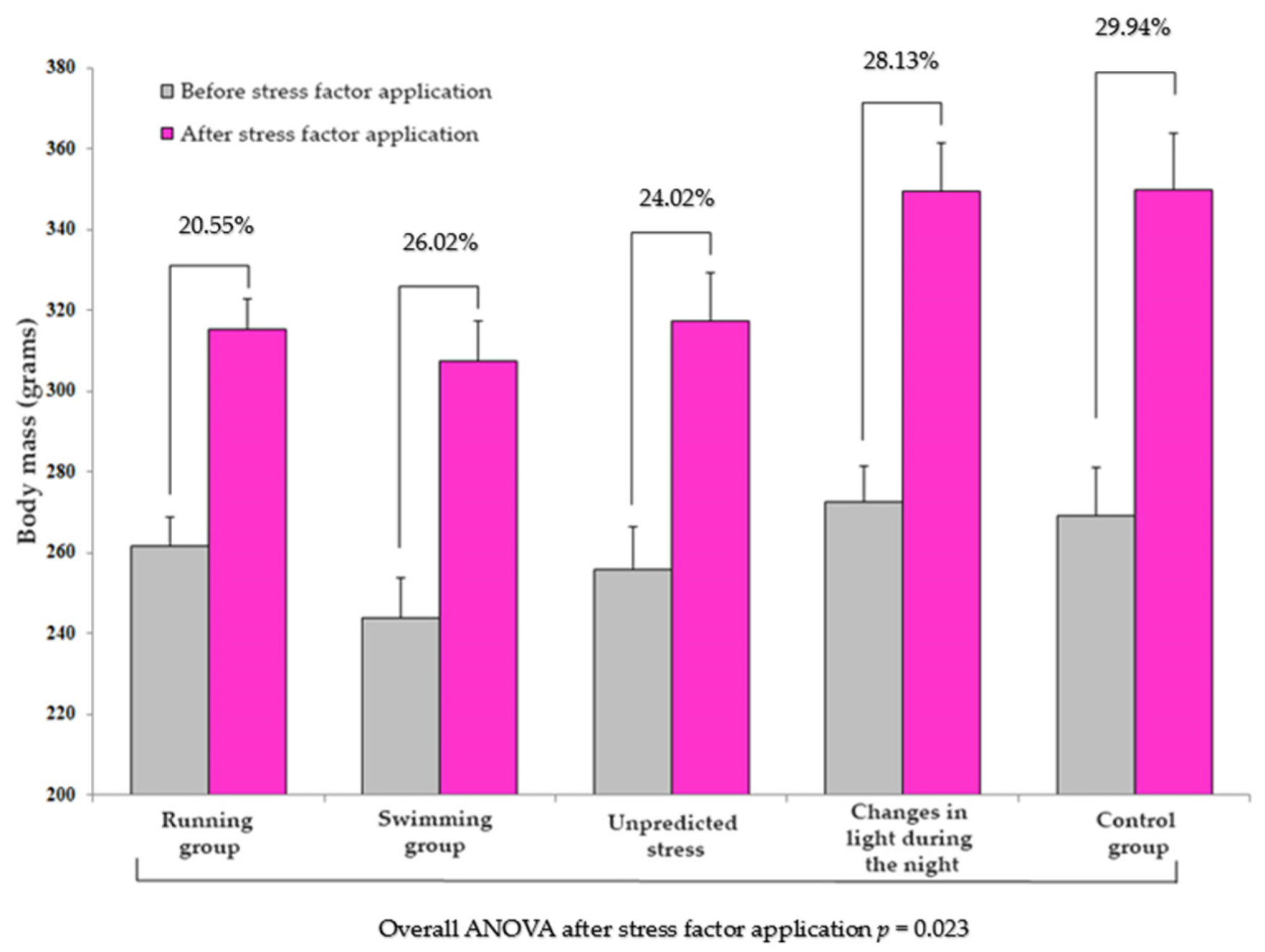
3.2. Dynamics of Body Mass Depending on the Application of the Stress Factor
4. Discussion
5. Limitations and Strengths
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Riaz, M.S.; Bohlen, M.O.; Gunter, B.W.; Quentin, H.; Stockmeier, C.A.; Paul, I.A. Attenuation of social interaction-associated ultrasonic vocalizations and spatial working memory performance in rats exposed to chronic unpredictable stress. Physiol. Behav. 2015, 152, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.R.; Crawley, J.N. Chapter 5—Anxiety-Related Behaviors in Mice. In Methods of Behavior Analysis in Neuroscience, 2nd ed.; Buccafusco, J.J., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009. [Google Scholar]
- Liu, J.; Yeo, H.C.; Overvik-Douki, E.; Hagen, T.; Doniger, S.J.; Chyu, D.W.; Brooks, G.A.; Ames, B.N. Chronically and acutely exercised rats: Biomarkers of oxidative stress and endogenous antioxidants. J. Appl. Physiol. 2000, 89, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L. Melatonin receptors: Role on sleep and circadian rhythm regulation. Sleep Med. 2007, 8, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.C.; Fogaça, M.V.; Aguiar, D.C.; Guimarães, F.S. Animal models of anxiety disorders and stress. Braz. J. Psychiatry 2013, 35, S101–S111. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R.; Strazielle, C. Relations between open-field, elevated plus-maze, and emergence tests as displayed by C57/BL6J and BALB/c mice. J. Neurosci. Methods 2008, 171, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Tatem, K.S.; Quinn, J.L.; Phadke, A.; Yu, Q.; Gordish-Dressman, H.; Nagaraju, K. Behavioral and locomotor measurements using an open field activity monitoring system for skeletal muscle diseases. J. Vis. Exp. 2014, 91, 51785. [Google Scholar] [CrossRef]
- Aslani, S.; Harb, M.R.; Costa, P.S.; Almeida, O.F.; Sousa, N.; Palha, J.A. Day and night: Diurnal phase influences the response to chronic mild stress. Front. Behav. Neurosci. 2014, 8, 82. [Google Scholar] [CrossRef] [PubMed]
- Nagy, S.A.; Vranesics, A.; Varga, Z.; Csabai, D.; Bruszt, N.; Bali, Z.K.; Perlaki, G.; Hernádi, I.; Berente, Z.; Miseta, A.; et al. Stress-Induced Microstructural Alterations Correlate with the Cognitive Performance of Rats: A Longitudinal in vivo Diffusion Tensor Imaging Study. Front. Neurosci. 2020, 14, 474. [Google Scholar] [CrossRef]
- Agorastos, A.; Nicolaides, N.C.; Bozikas, V.P.; Chrousos, G.P.; Pervanidou, P. Multilevel Interactions of Stress and Circadian System: Implications for Traumatic Stress. Front. Psychiatry 2020, 10, 1003. [Google Scholar] [CrossRef] [PubMed]
- Helfrich-Förster, C. Interactions between psychosocial stress and the circadian endogenous clock. PsyCh J. 2017, 6, 277–289. [Google Scholar] [CrossRef]
- Dauchy, R.T.; Dauchy, E.M.; Hanifin, J.P.; Gauthreaux, S.L.; Mao, L.; Belancio, V.P.; Ooms, T.G.; Dupepe, L.M.; Jablonski, M.R.; Warfield, B.; et al. Effects of spectral transmittance through standard laboratory cages on circadian metabolism and physiology in nude rats. J. Am. Assoc. Lab. Anim. Sci. 2013, 52, 146–156. [Google Scholar]
- Gan, E.H.; Quinton, R. Physiological significance of the rhythmic secretion of hypothalamic and pituitary hormones. Prog. Brain Res. 2010, 181, 111–126. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; Fliers, E.; Romijn, J.A.; La Fleur, S.E.; Wortel, J.; Bakker, O.; Endert, E.; Buijs, R.M. The suprachiasmatic nucleus generates the diurnal changes in plasma leptin levels. Endocrinology 2001, 142, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Gündüz, B. Daily rhythm in serum melatonin and leptin levels in the Syrian hamster (Mesocricetus auratus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 132, 393–401. [Google Scholar] [CrossRef]
- Wagner, R.; Oberste-Berghaus, C.; Herpertz, S.; Blum, W.F.; Pelz, B.; Hebebrand, J.; Senf, W.; Mann, K.; Albers, N. Time relationship between circadian variation of serum levels of leptin, insulin and cortisol in healthy subjects. Horm. Res. 2000, 54, 174–180. [Google Scholar] [CrossRef]
- Zinovkin, R.A.; Romaschenko, V.P.; Galkin, I.I.; Zakharova, V.V.; Pletjushkina, O.Y.; Chernyak, B.V.; Popova, E.N. Role of mitochondrial reactive oxygen species in age-related inflammatory activation of endothelium. Aging 2014, 6, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Shen, W.; Liu, Z.; Guan, S.; Liu, J.; Ding, S. Endurance exercise causes mitochondrial and oxidative stress in rat liver: Effects of a combination of mitochondrial targeting nutrients. Life Sci. 2010, 86, 39–44. [Google Scholar] [CrossRef]
- Silva, M.G.; Nunes, P.; Oliveira, P.; Ferreira, R.; Fardilha, M.; Moreira-Gonçalves, D.; Duarte, J.A.; Oliveira, M.M.; Peixoto, F. Long-Term Aerobic Training Improves Mitochondrial and Antioxidant Function in the Liver of Wistar Rats Preventing Hepatic Age-Related Function Decline. Biology 2022, 11, 1750. [Google Scholar] [CrossRef]
- Calil, C.M.; Marcondes, F.K. The comparison of immobility time in experimental rat swimming models. Life Sci. 2006, 79, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.L.; Hurley, C.; Repucci, N.; Drugan, R.C. Behavioral analysis of stress controllability effects in a new swim stress paradigm. Pharmacol. Biochem. Behav. 2001, 68, 263–272. [Google Scholar] [CrossRef]
- Bruner, C.A.; Vargas, I. The activity of rats in a swimming situation as a function of water temperature. Physiol. Behav. 1994, 55, 21–28. [Google Scholar] [CrossRef]
- Arshadi, S.; Bakhtiyari, S.; Haghani, K.; Valizadeh, A. Effects of Fenugreek Seed Extract and Swimming Endurance Training on Plasma Glucose and Cardiac Antioxidant Enzymes Activity in Streptozotocin-induced Diabetic Rats. Osong Public Health Res. Perspect. 2015, 6, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Teerapornpuntakit, J.; Dorkkam, N.; Wongdee, K.; Krishnamra, N.; Charoenphandhu, N. Endurance swimming stimulates transepithelial calcium transport and alters the expression of genes related to calcium absorption in the intestine of rats. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E775–E786. [Google Scholar] [CrossRef] [PubMed]
- Lapmanee, S.; Charoenphandhu, N.; Krishnamra, N.; Charoenphandhu, J. Anxiolytic-like actions of reboxetine, venlafaxine and endurance swimming in stressed male rats. Behav. Brain Res. 2012, 231, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Ströhle, A. Physical activity, exercise, depression and anxiety disorders. J. Neural. Transm. 2009, 116, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Cotman, C.W.; Berchtold, N.C. Exercise: A behavioral intervention to enhance brain health and plasticity. Trends Neurosci. 2002, 25, 295–301. [Google Scholar] [CrossRef]
- Radak, Z.; Toldy, A.; Szabo, Z.; Siamilis, S.; Nyakas, C.; Silye, G.; Jakus, J.; Goto, S. The effects of training and detraining on memory, neurotrophins and oxidative stress markers in rat brain. Neurochem. Int. 2006, 49, 387–392. [Google Scholar] [CrossRef]
- Filipović, D.; Gavrilović, L.; Dronjak, S.; Radojcić, M.B. The effect of repeated physical exercise on hippocampus and brain cortex in stressed rats. Ann. N. Y. Acad. Sci. 2007, 1096, 207–219. [Google Scholar] [CrossRef]
- Contarteze, R.V.L.; Manchado, F.B.; Gobatto, C.A.; De Mello, M.A.R. Stress biomarkers in rats submitted to swimming and treadmill running exercises. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 151, 415–422. [Google Scholar] [CrossRef]
- Voltarelli, F.A.; Gobatto, C.A.; de Mello, M.A. Determination of anaerobic threshold in rats using the lactate minimum test. Braz. J. Med. Biol. Res. 2002, 35, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. The Chronic Mild Stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2016, 6, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef]
- Fang, X.; Jiang, S.; Wang, J.; Bai, Y.; Kim, C.S.; Blake, D.; Weintraub, N.L.; Lei, Y.; Lu, X.Y. Chronic unpredictable stress induces depression-related behaviors by suppressing AgRP neuron activity. Mol. Psychiatry 2021, 26, 2299–2315. [Google Scholar] [CrossRef]
- Zhang, B.; Dong, W.; Ma, Z.; Duan, S.; Han, R.; Lv, Z.; Liu, X.; Mao, Y. Hyperbaric oxygen improves depression-like behaviors in chronic stress model mice by remodeling gut microbiota and regulating host metabolism. CNS Neurosci. Ther. 2023, 29, 239–255. [Google Scholar] [CrossRef] [PubMed]
- Der-Avakian, A.; Markou, A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 2012, 35, 68–77. [Google Scholar] [CrossRef]
- Taylor, P.J.; Gooding, P.; Wood, A.M.; Tarrier, N. The role of defeat and entrapment in depression, anxiety, and suicide. Psychol. Bull. 2011, 137, 391–420. [Google Scholar] [CrossRef]
- Şahin, T.D.; Karson, A.; Balcı, F.; Yazır, Y.; Bayramgürler, D.; Utkan, T. TNF-alpha inhibition prevents cognitive decline and maintains hippocampal BDNF levels in the unpredictable chronic mild stress rat model of depression. Behav. Brain Res. 2015, 292, 233–240. [Google Scholar] [CrossRef]
- Yirmiya, R.; Goshen, I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef]
- Voumvourakis, K.I.; Sideri, E.; Papadimitropoulos, G.N.; Tsantzali, I.; Hewlett, P.; Kitsos, D.; Stefanou, M.; Bonakis, A.; Giannopoulos, S.; Tsivgoulis, G.; et al. The Dynamic Relationship between the Glymphatic System, Aging, Memory, and Sleep. Biomedicines 2023, 11, 2092. [Google Scholar] [CrossRef]
- Raynaud, A.; Meunier, N.; Acquistapace, A.; Bombail, V. Chronic variable stress exposure in male Wistar rats affects the first step of olfactory detection. Behav. Brain Res. 2015, 291, 36–45. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: Central role of the brain. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef] [PubMed]
- Vasconcellos, A.P.; Tabajara, A.S.; Ferrari, C.; Rocha, E.; Dalmaz, C. Effect of chronic stress on spatial memory in rats is attenuated by lithium treatment. Physiol. Behav. 2003, 79, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.; Cowan, C.S.M.; Callaghan, B.L.; Kan, J.M. Effects of early-life stress on fear memory in the developing rat. Curr. Opin. Behav. Sci. 2016, 7, 15–20. [Google Scholar] [CrossRef]
- Steimer, T. Animal models of anxiety disorders in rats and mice: Some conceptual issues. Dialogues Clin. Neurosci. 2011, 13, 495–506. [Google Scholar] [CrossRef]
- Rincón-Cortés, M.; Sullivan, R.M. Emergence of social behavior deficit, blunted corticolimbic activity and adult depression-like behavior in a rodent model of maternal maltreatment. Transl. Psychiatry 2016, 6, e930. [Google Scholar] [CrossRef]
- Timofte, D.; Ciobica, A.; Honceriu, C.; Trofin, F.; Ciobica, A.; Cojocaru, D. A mini-review on the effects of exercising and obsesity in Parkinson’s disease. Acad. Rom. Sci. Ann. Ser. Biol. Sci. 2017, 6, 100–113. [Google Scholar]
- Trofin, F.; Ciobica, A.; Cojocaru, D.; Chirazi, M.; Honceriu, C.; Trofin, L.; Serban, D.; Timofte, D.; Cojocaru, S.I.; Anton, E. Increased oxidative stress status in rat serum after five minutes treadmill exercise. Cent. Eur. J. Med. 2014, 9, 722–728. [Google Scholar] [CrossRef]
- Bains, R.S.; Wells, S.; Sillito, R.R.; Armstrong, J.D.; Cater, H.L.; Banks, G.; Nolan, P.M. Assessing mouse behaviour throughout the light/dark cycle using automated in-cage analysis tools. J. Neurosci. Methods 2018, 300, 37–47. [Google Scholar] [CrossRef]
- Padurariu, M.; Antioch, I.; Balmus, I.; Ciobica, A.; El-Lethey, H.S.; Kamel, M.M. Describing some behavioural animal models of anxiety and their mechanistics with special reference to oxidative stress and oxytocin relevance. Int. J. Vet. Sci. Med. 2017, 5, 98–104. [Google Scholar] [CrossRef]
- Leopoldo, A.S.; Lima-Leopoldo, A.P.; Nascimento, A.F.; Luvizotto, R.A.; Sugizaki, M.M.; Campos, D.H.; da Silva, D.C.; Padovani, C.R.; Cicogna, A.C. Classification of different degrees of adiposity in sedentary rats. Braz. J. Med. Biol. Res. 2016, 49, e5028. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Delgado, R.; Tapia Osorio, A.; Saderi, N.; Escobar, C. Disruption of circadian rhythms: A crucial factor in the etiology of depression. Depress. Res. Treat. 2011, 2011, 839743. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Members | Stress Factors |
|---|---|---|
| 1 | 1–8 | Running |
| 2 | 9–16 | Swimming |
| 3 | 17–24 | Unpredicted stress (alternating running, swimming, light) |
| 4 | 25–32 | Changes in light during the dark phase |
| 5 | 33–40 | Control group |
| Group | Day 9 | Day 10 | Day 11 | Day 12 | Day 13 |
|---|---|---|---|---|---|
| Running group | 1 km/h 7 min | 1.5 km/h 7 min | 1.5 km/h 10 min | 1.5 km/h 15 min | 2 km/h 15 min |
| Swimming group | 5 min | 8 min | 11 min | 15 min | 15 min |
| Unpredicted stress | Run 1 km/h 7 min | Swim 7 min | Run 1 km/h 10 min | Swim 15 min | Run 1.5 km/h 10 min |
| Groups | Days 14–18 | Days 19–23 | Days 24–28 | Days 29–34 |
|---|---|---|---|---|
| Running group | 2 km/h 15 min | 3 km/h 15 min | 3.5 km/h 15 min | 4 km/h 15 min |
| Swimming group | 15 min | 25 min | 35 min | 45 min |
| Changes in light during the night | Every hour 4 times (22–23, 24–1, 2–3, 4–5) | Every hour 6 times (21, 22–23, 24–1, 2–3, 4–5, 6–7) | Every 30 min—6 times (24–24:30, 1–1:30, 2–2:30, 3–3:30, 4–4:30, 5–5:30) | Every 30 min—8 times (23–23:30, 24–24:30, 1–1:30, 2–2:30, 3–3:30, 4–4:30, 5–5:30, 6–6:30) |
| Control group | No stressor was applied | |||
| Day | Activity |
|---|---|
| 14 | Swim—15 min |
| 15 | Run 2 km/h—10 min |
| 16 | The light was alternated on and off every hour during the dark cycle. (22–23, 24–1, 2–3, 4–5) |
| 17 | Swim—15 min |
| 18 | Run 2 km/h—10 min |
| 19 | The light was alternated on and off every hour during the dark cycle. (20–21, 22–23, 24–1, 2–3, 4–5, 6–7) |
| 20 | Swim—20 min |
| 21 | Run 2.5 km/h—10 min |
| 22 | The light was alternated on and off every hour during the dark cycle. (20–21, 22–23, 24–1, 2–3, 4–5, 6–7) |
| 23 | Swim—20 min |
| 24 | Run 2.5 km/h—10 min |
| 25 | The light was alternated on and off every 30 min, 6 times during the dark cycle. (light on from 24 to 24:30, 1 to 1:30, 2 to 2:30, 3:00 to 3:30, 4 to 4:30, 5 to 5:30) |
| 26 | Swim—25 min |
| 27 | Run 3 km/h—10 min |
| 28 | The light was alternated on and off every 30 min, 6 times during the dark cycle. (light on from 24 to 24:30, 1 to 1:30, 2 to 2:30, 3:00 to 3:30, 4 to 4:30, 5 to 5:30) |
| 29 | Swim—25 min |
| 30 | Run 3 km/h—10 min |
| 31 | The light was alternated on and off every 30 min, 8 times during the dark cycle. (light on from 23 to 23:30, 24:00 to 24:30,1 to 1:30, 2 to 2:30, 3:00 to 3:30, 4 to 4:30, 5 to 5:30, 6 to 6:30) |
| 32 | Swim—30 min |
| 33 | Run 3.5 km/h—10 min |
| 34 | The animals were sacrificed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexa, A.I.; Zamfir, C.L.; Bogdănici, C.M.; Oancea, A.; Maștaleru, A.; Abdulan, I.M.; Brănișteanu, D.C.; Ciobîcă, A.; Balmuș, M.; Stratulat-Alexa, T.; et al. The Impact of Chronic Stress on Behavior and Body Mass in New Animal Models. Brain Sci. 2023, 13, 1492. https://doi.org/10.3390/brainsci13101492
Alexa AI, Zamfir CL, Bogdănici CM, Oancea A, Maștaleru A, Abdulan IM, Brănișteanu DC, Ciobîcă A, Balmuș M, Stratulat-Alexa T, et al. The Impact of Chronic Stress on Behavior and Body Mass in New Animal Models. Brain Sciences. 2023; 13(10):1492. https://doi.org/10.3390/brainsci13101492
Chicago/Turabian StyleAlexa, Anisia Iuliana, Carmen Lăcrămioara Zamfir, Camelia Margareta Bogdănici, Andra Oancea, Alexandra Maștaleru, Irina Mihaela Abdulan, Daniel Constantin Brănișteanu, Alin Ciobîcă, Miruna Balmuș, Teodora Stratulat-Alexa, and et al. 2023. "The Impact of Chronic Stress on Behavior and Body Mass in New Animal Models" Brain Sciences 13, no. 10: 1492. https://doi.org/10.3390/brainsci13101492