
Development of Gamma Oscillation during Sentence Processing in Early Adolescence: Insights into the Maturation of Semantic Processing

 , and
, and
Abstract
:1. Introduction
Related Work
2. Materials and Methods
2.1. Participants
2.2. Linguistic Stimuli
2.3. EEG Acquisition
2.4. EEG Pre-Processing
2.5. Time–Frequency Analysis
2.6. Statistical Analysis
3. Results
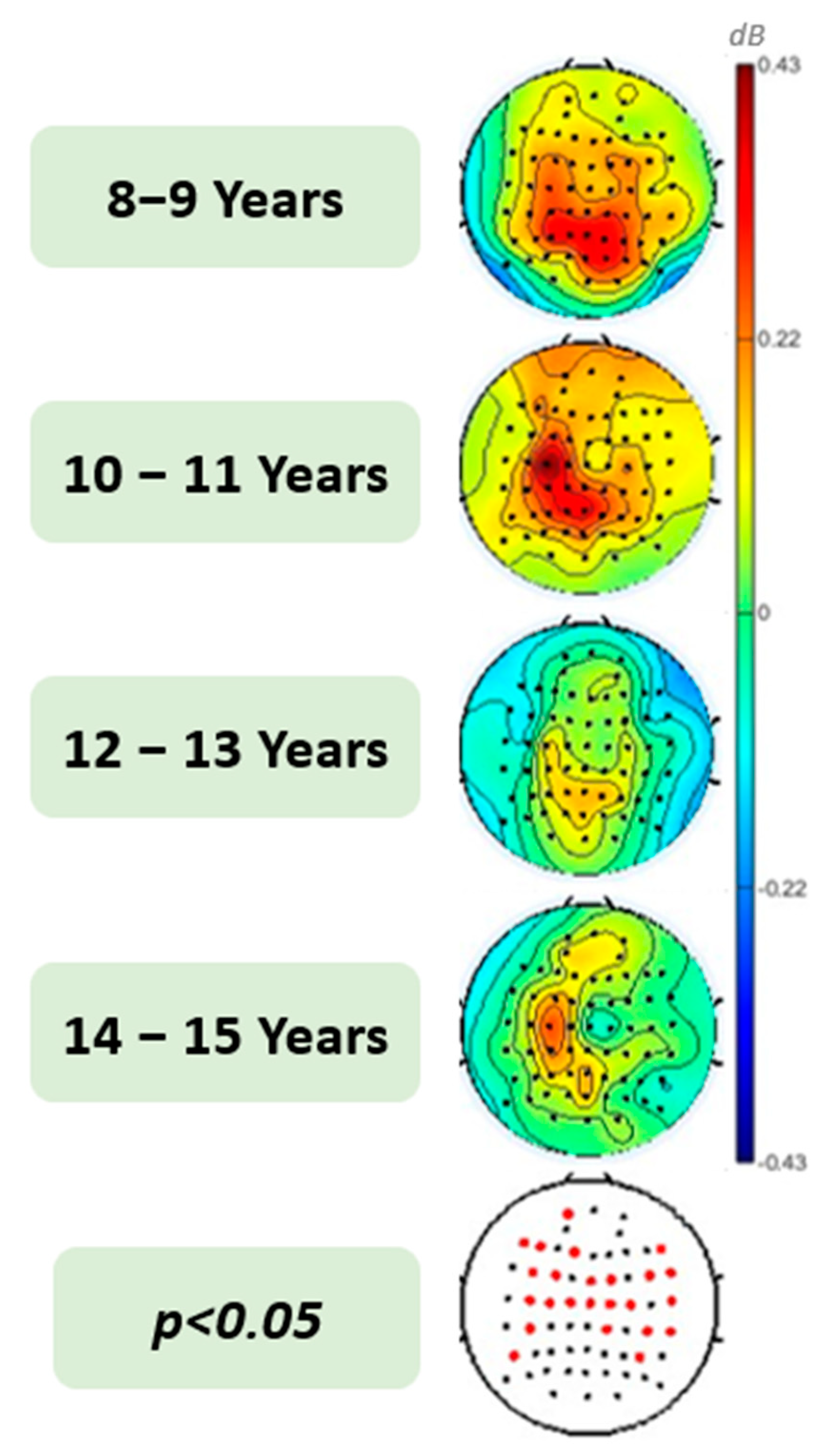
3.1. Spatial Analysis
3.2. Spatiotemporal Analysis
3.3. Temporal Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lew-Williams, C.; Fernald, A. Young Children Learning Spanish Make Rapid Use of Grammatical Gender in Spoken Word Recognition. Psychol. Sci. 2007, 18, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.J.; Schneider, J.M.; Melamed, T.C.; Ralph, Y.K.; Poudel, S.; Raval, V.M.; Mikhail, D.; Abel, A.D. Temporal and topographical changes in theta power between middle childhood and adolescence during sentence comprehension. Dev. Cogn. Neurosci. 2022, 53, 101056. Available online: https://www.sciencedirect.com/science/article/pii/S1878929321001456 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Nippold, M.A.; Hesketh, L.J.; Duthie, J.K.; Mansfield, T.C. Conversational versus expository discourse: A study of syntactic development in children, adolescents, and adults. J. Speech Lang. Hear. Res. 2005, 48, 1048–1064. [Google Scholar] [CrossRef]
- Nuñez, S.C.; Dapretto, M.; Katzir, T.; Starr, A.; Bramen, J.; Kan, E.; Bookheimer, S.; Sowell, E.R. fMRI of syntactic processing in typically developing children: Structural correlates in the inferior frontal gyrus. Dev. Cogn. Neurosci. 2011, 1, 313–323. Available online: https://www.sciencedirect.com/science/article/pii/S1878929311000259 (accessed on 15 November 2023). [CrossRef]
- Schneider, J.M.; Abel, A.D.; Ogiela, D.A.; McCord, C.; Maguire, M.J. Developmental differences in the neural oscillations underlying auditory sentence processing in children and adults. Brain Lang. 2018, 186, 17–25. Available online: https://www.sciencedirect.com/science/article/pii/S0093934X17301190 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Skeide, M.A.; Brauer, J.; Friederici, A.D. Syntax gradually segregates from semantics in the developing brain. NeuroImage 2014, 100, 106–111. Available online: https://www.sciencedirect.com/science/article/pii/S1053811914004650 (accessed on 15 November 2023). [CrossRef]
- Wang, S.; Allen, R.J.; Lee, J.R.; Hsieh, C.E. Evaluating the developmental trajectory of the episodic buffer component of working memory and its relation to word recognition in children. J. Exp. Child Psychol. 2015, 133, 16–28. Available online: https://www.sciencedirect.com/science/article/pii/S0022096515000144 (accessed on 15 November 2023). [CrossRef]
- Wu, C.Y.; Vissiennon, K.; Friederici, A.D.; Brauer, J. Preschoolers’ brains rely on semantic cues prior to the mastery of syntax during sentence comprehension. NeuroImage 2016, 126, 256–266. Available online: https://www.sciencedirect.com/science/article/pii/S1053811915009477 (accessed on 15 November 2023). [CrossRef]
- Hurks, P.P.M.; Vles, J.S.H.; Hendriksen, J.G.M.; Kalff, A.C.; Feron, F.J.M.; Kroes, M.; van Zeben, T.M.C.B.; Steyaert, J.; Jolles, J. Semantic Category Fluency Versus Initial Letter Fluency Over 60 Seconds as a Measure of Automatic and Controlled Processing in Healthy School-aged Children. J. Clin. Exp. Neuropsychol. 2006, 28, 684–695. [Google Scholar] [CrossRef]
- Mahler, N.A.; Chenery, H.J. A Developmental Perspective on Processing Semantic Context: Preliminary Evidence from Sentential Auditory Word Repetition in School-Aged Children. J. Psycholinguist. Res. 2019, 48, 81–105. Available online: http://www.scopus.com/inward/record.url?scp=85049695118&partnerID=8YFLogxK (accessed on 15 November 2023). [CrossRef]
- Nouwens, S.; Groen, M.A.; Kleemans, T.; Verhoeven, L. The role of semantic retrieval in children’s reading comprehension development in the upper primary grades. J. Res. Read. 2018, 41, 597–614. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/1467-9817.12128 (accessed on 15 November 2023). [CrossRef]
- Hahne, A.; Eckstein, K.; Friederici, A.D. Brain signatures of syntactic and semantic processes during children’s language development. J. Cogn. Neurosci. 2004, 16, 1302–1318. [Google Scholar] [CrossRef] [PubMed]
- Bastiaansen, M.C.M.; van der Linden, M.; ter Keurs, M.; Dijkstra, T.; Hagoort, P. Theta Responses Are Involved in Lexical—Semantic Retrieval during Language Processing. J. Cogn. Neurosci. 2005, 17, 530–541. [Google Scholar] [CrossRef]
- Hald, L.A.; Bastiaansen, M.C.M.; Hagoort, P. EEG theta and gamma responses to semantic violations in online sentence processing. Brain Lang. 2006, 96, 90–105. Available online: https://www.sciencedirect.com/science/article/pii/S0093934X05001239 (accessed on 15 November 2023). [CrossRef]
- Lam, N.H.L.; Schoffelen, J.M.; Uddén, J.; Hultén, A.; Hagoort, P. Neural activity during sentence processing as reflected in theta, alpha, beta, and gamma oscillations. NeuroImage 2016, 142, 43–54. Available online: https://www.sciencedirect.com/science/article/pii/S1053811916002032 (accessed on 15 November 2023). [CrossRef]
- Maguire, M.J.; Brier, M.R.; Ferree, T.C. EEG theta and alpha responses reveal qualitative differences in processing taxonomic versus thematic semantic relationships. Brain Lang. 2010, 114, 16–25. Available online: https://www.sciencedirect.com/science/article/pii/S0093934X10000520 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Meyer, L. The neural oscillations of speech processing and language comprehension: State of the art and emerging mechanisms. Eur. J. Neurosci. 2018, 48, 2609–2621. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/ejn.13748 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Penolazzi, B.; Angrilli, A.; Job, R. Gamma EEG activity induced by semantic violation during sentence reading. Neurosci. Lett. 2009, 465, 74–78. Available online: https://www.sciencedirect.com/science/article/pii/S0304394009011719 (accessed on 15 November 2023). [CrossRef]
- Schneider, J.M.; Maguire, M.J. Developmental differences in the neural correlates supporting semantics and syntax during sentence processing. Dev. Sci. 2019, 22, e12782. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/desc.12782 (accessed on 15 November 2023). [CrossRef]
- Bastiaansen, M.; Hagoort, P. Oscillatory neuronal dynamics during language comprehension. Prog. Brain Res. 2006, 159, 179–196. Available online: https://www.sciencedirect.com/science/article/pii/S0079612306590120 (accessed on 15 November 2023).
- Bastiaansen, M.; Hagoort, P. Frequency-based Segregation of Syntactic and Semantic Unification during Online Sentence Level Language Comprehension. J. Cogn. Neurosci. 2015, 27, 2095–2107. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, Z.; Bastiaansen, M. Integration or Predictability? A Further Specification of the Functional Role of Gamma Oscillations in Language Comprehension. Front. Psychol. 2012, 3, 187. Available online: https://www.frontiersin.org/articles/10.3389/fpsyg.2012.00187 (accessed on 15 November 2023). [CrossRef]
- Prystauka, Y.; Lewis, A.G. The Power of Neural Oscillations to Inform Sentence Comprehension: A Linguistic Perspective. Lang Linguist. Compass 2019, 13, e12347. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7546279/ (accessed on 15 November 2023). [CrossRef] [PubMed]
- Kaiser, J.; Lutzenberger, W. Human gamma-band activity: A window to cognitive processing. NeuroReport 2005, 16, 207. Available online: https://journals.lww.com/neuroreport/Fulltext/2005/02280/Combined_EEG_and_MEG_recordings_of_visual_40_Hz.1.aspx?casa_token=e-lSlYwy7ncAAAAA:3IAdC0s2Z-QNQRufwgdGtUKnefB63_R97NGfpSeZ2yw6gfsvosTlMopYaIwSefPe27aDf1KprpjfU-hegz02SC0kFA (accessed on 15 November 2023). [CrossRef] [PubMed]
- Ulloa, J.L. The Control of Movements via Motor Gamma Oscillations. Front. Hum. Neurosci. 2022, 15, 787157. Available online: https://www.frontiersin.org/articles/10.3389/fnhum.2021.787157 (accessed on 15 November 2023). [CrossRef]
- Muthukumaraswamy, S.D. Functional Properties of Human Primary Motor Cortex Gamma Oscillations. J. Neurophysiol. 2010, 104, 2873–2885. Available online: https://journals.physiology.org/doi/full/10.1152/jn.00607.2010 (accessed on 15 November 2023). [CrossRef]
- Keil, A.; Gruber, T.; Müller, M.M. Functional correlates of macroscopic high-frequency brain activity in the human visual system. Neurosci. Biobehav. Rev. 2001, 25, 527–534. Available online: https://www.sciencedirect.com/science/article/pii/S0149763401000318 (accessed on 15 November 2023). [CrossRef]
- Friese, U.; Köster, M.; Hassler, U.; Martens, U.; Trujillo-Barreto, N.; Gruber, T. Successful memory encoding is associated with increased cross-frequency coupling between frontal theta and posterior gamma oscillations in human scalp-recorded EEG. NeuroImage 2013, 66, 642–647. Available online: https://www.sciencedirect.com/science/article/pii/S105381191201097X (accessed on 15 November 2023). [CrossRef]
- Gruber, T.; Tsivilis, D.; Montaldi, D.; Müller, M.M. Induced gamma band responses: An early marker of memory encoding and retrieval. NeuroReport 2004, 15, 1837. Available online: https://journals.lww.com/neuroreport/fulltext/2004/08060/Induced_gamma_band_responses__an_early_marker_of.30.aspx?casa_token=G5ex5piwVk4AAAAA:0WiZDlFm54-hHMYBi4XQaFtjR8wjJctAyelblKH_QysP3_PQkB3OCnfw7moUyr_xoz-zoE6N9boOGeBkl_z9S32xaQ (accessed on 15 November 2023). [CrossRef]
- Osipova, D.; Takashima, A.; Oostenveld, R.; Fernández, G.; Maris, E.; Jensen, O. Theta and Gamma Oscillations Predict Encoding and Retrieval of Declarative Memory. J. Neurosci. 2006, 26, 7523–7531. Available online: https://www.jneurosci.org/content/26/28/7523 (accessed on 15 November 2023). [CrossRef]
- Honkanen, R.; Rouhinen, S.; Wang, S.H.; Palva, J.M.; Palva, S. Gamma Oscillations Underlie the Maintenance of Feature-Specific Information and the Contents of Visual Working Memory. Cereb. Cortex 2015, 25, 3788–3801. [Google Scholar] [CrossRef]
- Roux, F.; Uhlhaas, P.J. Working memory and neural oscillations: Alpha–gamma versus theta–gamma codes for distinct WM information? Trends Cogn. Sci. 2014, 18, 16–25. Available online: https://www.cell.com/trends/cognitive-sciences/abstract/S1364-6613(13)00231-3 (accessed on 15 November 2023). [CrossRef]
- van Vugt, M.K.; Schulze-Bonhage, A.; Litt, B.; Brandt, A.; Kahana, M.J. Hippocampal Gamma Oscillations Increase with Memory Load. J. Neurosci. 2010, 30, 2694–2699. Available online: https://www.jneurosci.org/content/30/7/2694 (accessed on 15 November 2023). [CrossRef]
- Chaieb, L.; Leszczynski, M.; Axmacher, N.; Höhne, M.; Elger, C.E.; Fell, J. Theta-gamma phase-phase coupling during working memory maintenance in the human hippocampus. Cogn. Neurosci. 2015, 6, 149–157. [Google Scholar] [CrossRef]
- Lenroot, R.K.; Giedd, J.N. Brain development in children and adolescents: Insights from anatomical magnetic resonance imaging. Neurosci. Biobehav. Rev. 2006, 30, 718–729. Available online: https://www.sciencedirect.com/science/article/pii/S0149763406000455 (accessed on 15 November 2023). [CrossRef]
- Diamond, A.; Goldman-Rakic, P.S. Comparison of human infants and rhesus monkeys on Piaget’s AB task: Evidence for dependence on dorsolateral prefrontal cortex. Exp. Brain Res. 1989, 74, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Luna, B.; Sweeney, J.A. Studies of brain and cognitive maturation through childhood and adolescence: A strategy for testing neurodevelopmental hypotheses. Schizophr. Bull. 2001, 27, 443–455. [Google Scholar] [CrossRef]
- Poudel, S.; Denicola-Prechtl, K.; Nelson, J.A.; Behboudi, M.H.; Benitez-Barrera, C.; Castro, S.; Maguire, M.J. Rethinking household size and children’s language environment. Dev. Psychol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.J.; Schneider, J.M. Socioeconomic status related differences in resting state EEG activity correspond to differences in vocabulary and working memory in grade school. Brain Cogn. 2019, 137, 103619. Available online: https://www.sciencedirect.com/science/article/pii/S0278262618304755 (accessed on 1 November 2023). [CrossRef] [PubMed]
- Brauer, J.; Anwander, A.; Friederici, A.D. Neuroanatomical Prerequisites for Language Functions in the Maturing Brain. Cereb. Cortex 2011, 21, 459–466. [Google Scholar] [CrossRef]
- Friederici, A.D.; Brauer, J.; Lohmann, G. Maturation of the Language Network: From Inter- to Intrahemispheric Connectivities. PLoS ONE 2011, 6, e20726. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0020726 (accessed on 15 November 2023). [CrossRef]
- Brauer, J.; Anwander, A.; Perani, D.; Friederici, A.D. Dorsal and ventral pathways in language development. Brain Lang. 2013, 127, 289–295. Available online: https://www.sciencedirect.com/science/article/pii/S0093934X13000709 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Gogtay, N.; Giedd, J.N.; Lusk, L.; Hayashi, K.M.; Greenstein, D.; Vaituzis, A.C.; Nygent, T.F., III; Herman, D.H.; Clasen, L.S.; Toga, A.W.; et al. Dynamic mapping of human cortical development during childhood through early adulthood. Proc. Natl. Acad. Sci. USA 2004, 101, 8174–8179. Available online: https://www.pnas.org/doi/abs/10.1073/pnas.0402680101 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Krause, C.M.; Salminen, P.A.; Sillanmäki, L.; Holopainen, I.E. Event-related desynchronization and synchronization during a memory task in children. Clin. Neurophysiol. 2001, 112, 2233–2240. Available online: https://www.sciencedirect.com/science/article/pii/S1388245701006848 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Spironelli, C.; Angrilli, A. Developmental aspects of language lateralization in delta, theta, alpha and beta EEG bands. Biol. Psychol. 2010, 85, 258–267. Available online: https://www.sciencedirect.com/science/article/pii/S0301051110001924 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Benasich, A.A.; Gou, Z.; Choudhury, N.; Harris, K.D. Early cognitive and language skills are linked to resting frontal gamma power across the first 3 years. Behav. Brain Res. 2008, 195, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Cantiani, C.; Piazza, C.; Mornati, G.; Molteni, M.; Riva, V. Oscillatory gamma activity mediates the pathway from socioeconomic status to language acquisition in infancy. Infant Behav. Dev. 2019, 57, 101384. Available online: https://www.sciencedirect.com/science/article/pii/S0163638319301390 (accessed on 15 November 2023). [CrossRef]
- Lum, J.A.G.; Clark, G.M.; Bigelow, F.J.; Enticott, P.G. Resting state electroencephalography (EEG) correlates with children’s language skills: Evidence from sentence repetition. Brain Lang. 2022, 230, 105137. Available online: https://www.sciencedirect.com/science/article/pii/S0093934X22000670 (accessed on 15 November 2023). [CrossRef]
- Heim, S.; Keil, A.; Choudhury, N.; Thomas Friedman, J.; Benasich, A.A. Early gamma oscillations during rapid auditory processing in children with a language-learning impairment: Changes in neural mass activity after training. Neuropsychologia 2013, 51, 990–1001. Available online: https://www.sciencedirect.com/science/article/pii/S0028393213000171 (accessed on 15 November 2023). [CrossRef]
- Friederici, A.D. Children’s sensitivity to function words during sentence comprehension. De Gruyter Mouton 1983, 21, 717–740. Available online: https://www.degruyter.com/document/doi/10.1515/ling.1983.21.5.717/html?lang=en (accessed on 15 November 2023). [CrossRef]
- Duffau, H.; Gatignol, P.; Mandonnet, E.; Peruzzi, P.; Tzourio-Mazoyer, N.; Capelle, L. New insights into the anatomo-functional connectivity of the semantic system: A study using cortico-subcortical electrostimulations. Brain 2005, 128 Pt 4, 797–810. [Google Scholar] [CrossRef]
- Friederici, A.D. Pathways to language: Fiber tracts in the human brain. Trends Cogn. Sci. 2009, 13, 175–181. [Google Scholar] [CrossRef]
- Friederici, A.D. The brain basis of language processing: From structure to function. Physiol. Rev. 2011, 91, 1357–1392. [Google Scholar] [CrossRef]
- Sakai, K.L. Language Acquisition and Brain Development. Science 2005, 310, 815–819. Available online: https://www.science.org/doi/10.1126/science.1113530 (accessed on 15 November 2023). [CrossRef]
- Vigneau, M.; Beaucousin, V.; Hervé, P.Y.; Duffau, H.; Crivello, F.; Houdé, O.; Mazoyer, B.; Tzourio-Mazoyer, N. Meta-analyzing left hemisphere language areas: Phonology, semantics, and sentence processing. NeuroImage 2006, 30, 1414–1432. Available online: https://www.sciencedirect.com/science/article/pii/S1053811905024511 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Caplan, D. Functional neuroimaging studies of syntactic processing. J Psycholinguist. Res. 2001, 30, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Santi, A.; Grodzinsky, Y. fMRI adaptation dissociates syntactic complexity dimensions. NeuroImage 2010, 51, 1285–1293. Available online: https://www.sciencedirect.com/science/article/pii/S1053811910003216. (accessed on 15 November 2023). [CrossRef]
- Brauer, J.; Friederici, A.D. Functional neural networks of semantic and syntactic processes in the developing brain. J. Cogn. Neurosci. 2007, 19, 1609–1623. [Google Scholar] [CrossRef] [PubMed]
- Rommers, J.; Dijkstra, T.; Bastiaansen, M. Context-dependent Semantic Processing in the Human Brain: Evidence from Idiom Comprehension. J. Cogn. Neurosci. 2013, 25, 762–776. [Google Scholar] [CrossRef]
- Giannopoulos, A.E.; Zioga, I.; Kontoangelos, K.; Papageorgiou, P.; Kapsali, F.; Capsalis, C.N.; Papageorgiou, C. Deciding on Optical Illusions: Reduced Alpha Power in Body Dysmorphic Disorder. Brain Sci. 2022, 12, 293. Available online: https://www.mdpi.com/2076-3425/12/2/293 (accessed on 15 November 2023). [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. Available online: https://www.sciencedirect.com/science/article/pii/S0165027003003479 (accessed on 15 November 2023). [CrossRef]
- Winkler, I.; Haufe, S.; Tangermann, M. Automatic classification of artifactual ICA-components for artifact removal in EEG signals. Behav. Brain Funct. 2011, 7, 30. [Google Scholar] [CrossRef]
- Winkler, I.; Brandl, S.; Horn, F.; Waldburger, E.; Allefeld, C.; Tangermann, M. Robust artifactual independent component classification for BCI practitioners. J. Neural Eng. 2014, 11, 035013. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.2517-6161.1995.tb02031.x (accessed on 15 November 2023). [CrossRef]
- Muller, P.; Parmigiani, G.; Rice, K.; FDR and Bayesian Multiple Comparisons Rules. Johns Hopkins University, Dept of Biostatistics Working Papers 18 July 2006. Available online: https://biostats.bepress.com/jhubiostat/paper115 (accessed on 15 November 2023).
- Fedorenko, E.; Scott, T.L.; Brunner, P.; Coon, W.G.; Pritchett, B.; Schalk, G.; Kanwisher, N. Neural correlate of the construction of sentence meaning. Proc. Natl. Acad. Sci. USA 2016, 113, E6256–E6262. Available online: https://pnas.org/doi/full/10.1073/pnas.1612132113 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Marslen-Wilson, W.; Tyler, L.K. Processing structure of sentence perception. Nature 1975, 257, 784–786. Available online: https://www.nature.com/articles/257784a0 (accessed on 15 November 2023). [CrossRef] [PubMed]
- Gentner, D. Verb semantic structures in memory for sentences: Evidence for componential representation. Cogn. Psychol. 1981, 13, 56–83. Available online: https://www.sciencedirect.com/science/article/pii/0010028581900049 (accessed on 15 November 2023). [CrossRef]
- Priyastini, N.; Puspani, I.A.M.; Antari, N.M.S. The Semantic Roles of the English Action Verbs. J. Hum. Fak. Ilmu Budaya Unud 2016, 16, 159–166. Available online: https://ojs.unud.ac.id/index.php/sastra/article/download/33496/20298 (accessed on 15 November 2023).
- Wang, J.; Wagley, N.; Rice, M.L.; Booth, J.R. Semantic and syntactic specialization during auditory sentence processing in 7–8-year-old children. Cortex 2021, 145, 169–186. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sentence 1 | Go outside and play with the neg | Target word: Cat |
| Sentence 2 | I’m covered with hair from the neg | |
| Sentence 3 | All night, I heard the meowing of the neg |
| He spilled food on his wesh. |
| The dog slept on the presh. |
| To keep the sun away, I wear a ruch. |
| Hi favorite toy of all time is the therg. |
| When I sit there, I can see the pleg. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behboudi, M.H.; Castro, S.; Chalamalasetty, P.; Maguire, M.J. Development of Gamma Oscillation during Sentence Processing in Early Adolescence: Insights into the Maturation of Semantic Processing. Brain Sci. 2023, 13, 1639. https://doi.org/10.3390/brainsci13121639
Behboudi MH, Castro S, Chalamalasetty P, Maguire MJ. Development of Gamma Oscillation during Sentence Processing in Early Adolescence: Insights into the Maturation of Semantic Processing. Brain Sciences. 2023; 13(12):1639. https://doi.org/10.3390/brainsci13121639
Chicago/Turabian StyleBehboudi, Mohammad Hossein, Stephanie Castro, Prasanth Chalamalasetty, and Mandy J. Maguire. 2023. "Development of Gamma Oscillation during Sentence Processing in Early Adolescence: Insights into the Maturation of Semantic Processing" Brain Sciences 13, no. 12: 1639. https://doi.org/10.3390/brainsci13121639
APA StyleBehboudi, M. H., Castro, S., Chalamalasetty, P., & Maguire, M. J. (2023). Development of Gamma Oscillation during Sentence Processing in Early Adolescence: Insights into the Maturation of Semantic Processing. Brain Sciences, 13(12), 1639. https://doi.org/10.3390/brainsci13121639