
Neuroplastic Changes in Addiction Memory—How Music Therapy and Music-Based Intervention May Reduce Craving: A Narrative Review
 , , , and
, , , and
Abstract
1. Introduction
Methods
2. Music’s Ability to Elicit Emotions and Evoke Memories: A Look at the Brain Correlates
2.1. Neural Correlates of Music-Evoked Emotions
2.2. Music Therapy and Music-Based Interventions for Substance Use Disorder
2.3. Music Is Able to Cue Emotionally Salient Autobiographical Memories
2.3.1. The Role of Emotion in Autobiographical Memory Retrieval
2.3.2. Neural Correlates of Music-Evoked Autobiographical Memories
3. Neural Circuitry of Drug Craving—The Role of a Brain Memory Network
3.1. Evidence from Human Imaging and Resting-State EEG
4. The Role of Perineuronal Nets in Addiction Memory Neuroplasticity

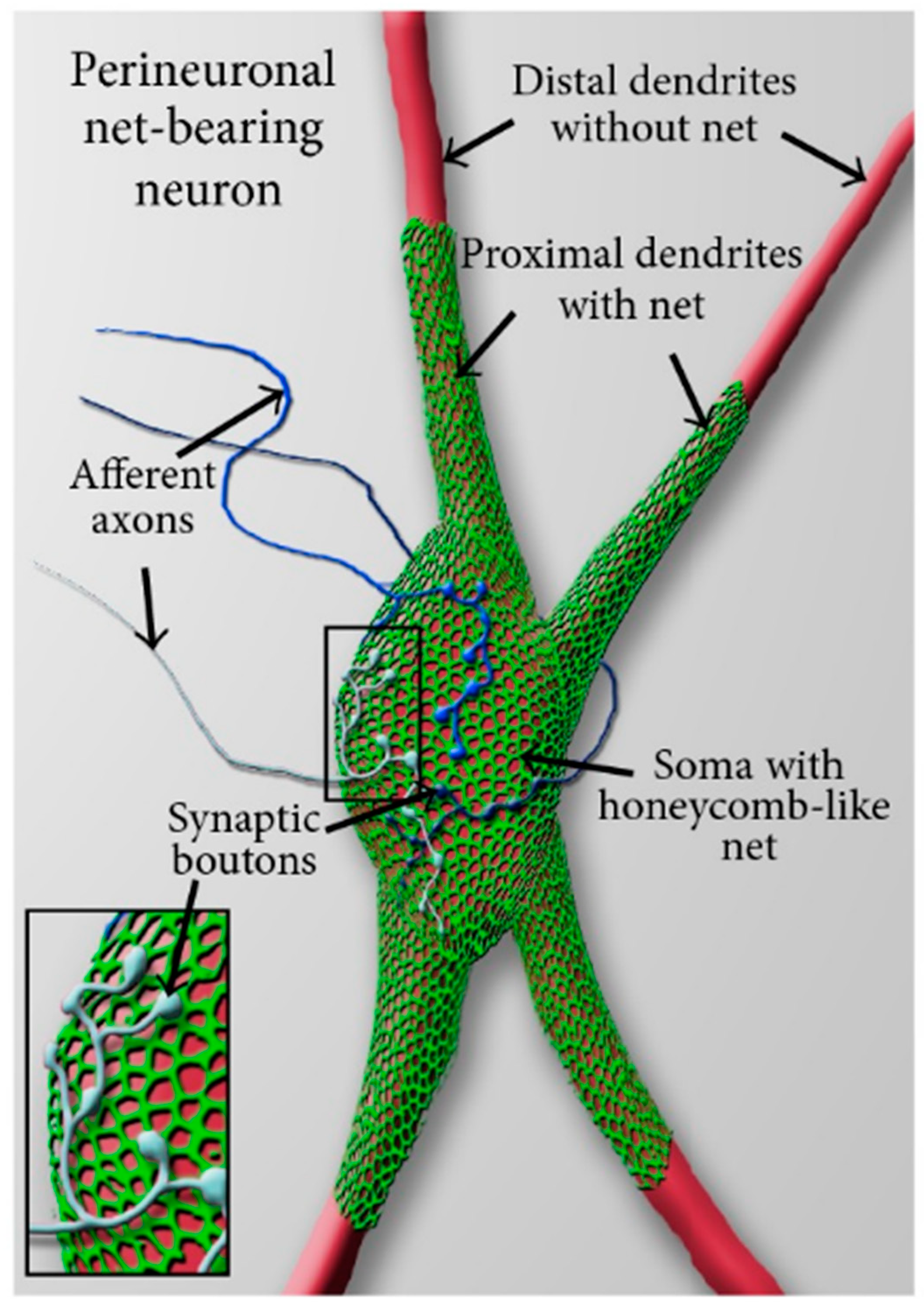
4.1. What Is a Perineuronal Net and How It Is Related to Emotional Memory
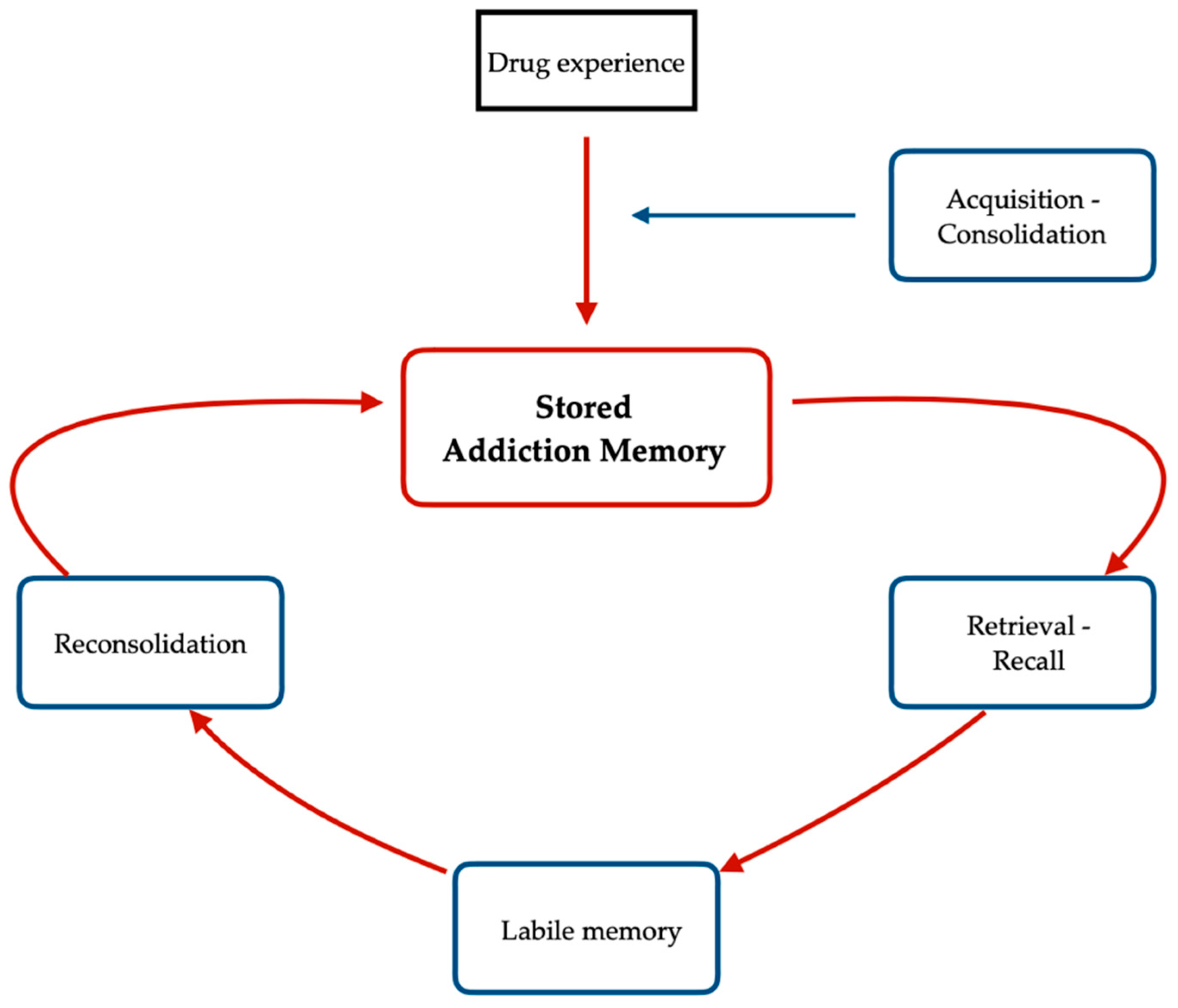
4.2. Acquisition and Reconsolidation of Addiction Memories

5. Conclusions
5.1. Implications for Music Therapy Research and Clinical Practice
5.2. Strengths and Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchie, H.; Roser, M. Our World in Data. Drug Use. Available online: https://ourworldindata.org/drug-use (accessed on 10 September 2022).
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013; p. 483. [Google Scholar]
- Flaudias, V.; Heeren, A.; Brousse, G.; Maurage, P. Toward a triadic approach to craving in addictive disorders: The metacognitive hub model. Harv. Rev. Psychiatry 2019, 27, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Noël, X.; Brevers, D.; Bechara, A. A triadic neurocognitive approach to addiction for clinical interventions. Front. Psychiatry 2013, 4, 179. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Volkow, N.D. Neurobiology of addiction: A neurocircuitry analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef]
- Meyer, J.S.; Quenzer, L.F.; Rice, S.A.; Yates, J.R. Psychopharmacology: Drugs, the Brain, and Behavior; Sinauer Associates: Sunderland, MA, USA, 2005. [Google Scholar]
- Stein, D.; Phillips, K.; Bolton, D.; Fulford, K.; Sadler, J.; Kendler, K. What is a mental/psychiatric disorder? From DSM-IV to DSM-V. Psychol. Med. 2010, 40, 1759–1765. [Google Scholar] [CrossRef]
- McWilliams, N. Psychoanalytic Diagnosis: Understanding Personality Structure in Clinical Process, 2nd ed.; Guilford Press: New York, NY, USA, 2011; p. 26. [Google Scholar]
- Markou, A.; Kosten, T.R.; Koob, G.F. Neurobiological similarities in depression and drug dependence: A self-medication hypothesis. Neuropsychopharmacology 1998, 18, 135–174. [Google Scholar] [CrossRef]
- Koob, G.F.; Le Moal, M. Addiction and the brain antireward system. Annu. Rev. Psychol. 2008, 59, 29–53. [Google Scholar] [CrossRef]
- Hyman, S.E. Addiction: A disease of learning and memory. Am. J. Psychiatry 2005, 162, 1414–1422. [Google Scholar] [CrossRef]
- Sorg, B.A. Reconsolidation of drug memories. Neurosci. Biobehav. Rev. 2012, 36, 1400–1417. [Google Scholar] [CrossRef]
- LeCocq, M.R.; Randall, P.A.; Besheer, J.; Chaudhri, N. Considering Drug-Associated Contexts in Substance Use Disorders and Treatment Development. Neurotherapeutics 2020, 17, 43–54. [Google Scholar] [CrossRef]
- Fachner, J. Music therapy, drugs and state-dependent recall. In Music Therapy and Addictions, 1st ed.; Aldridge, D., Fachner, J., Eds.; Jessica Kingsley Publishers: London, UK, 2010; Volume 1, pp. 18–34. [Google Scholar]
- Fachner, J. Retraining of drug reward, music cues and state-dependent recall in music therapy. Music Med. 2017, 9, 8–14. [Google Scholar] [CrossRef]
- Horesh, T. Drug addicts and their music—A Story of a Complex Relationship. In Music Therapy and Addictions, 1st ed.; Aldridge, D., Fachner, J., Eds.; Jessica Kingsley Publishers: London, UK, 2010; Volume 1, pp. 57–74. [Google Scholar]
- Cross, I. Music, cognition, culture, and evolution. Ann. N. Y. Acad. Sci. 2001, 930, 28–42. [Google Scholar] [CrossRef] [PubMed]
- What is Music Therapy? World Federation of Music Therapy. Available online: http://www.wfmt.info/wfmt-new-home/about-wfmt/ (accessed on 1 September 2022).
- American Music Therapy Association. Available online: www.musictherapy.org (accessed on 1 September 2022).
- Sihvonen, A.J.; Särkämö, T.; Leo, V.; Tervaniemi, M.; Altenmüller, E.; Soinila, S. Music-based interventions in neurological rehabilitation. Lancet Neurol. 2017, 16, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, L.; Bradt, J.; Stegemann, T.; Koelsch, S. Effects of music therapy and music-based interventions in the treatment of substance use disorders: A systematic review. PLoS ONE 2017, 12, e0187363. [Google Scholar] [CrossRef]
- Ghetti, C.; Chen, X.J.; Brenner, A.K.; Hakvoort, L.G.; Lien, L.; Fachner, J.; Gold, C. Music therapy for people with substance use disorders. Cochrane Database Syst. Rev. 2022, 5, 1–101. [Google Scholar] [CrossRef]
- Lasek, A.W.; Chen, H.; Chen, W.Y. Releasing Addiction Memories Trapped in Perineuronal Nets. Trends Genet. 2018, 34, 197–208. [Google Scholar] [CrossRef]
- Green, B.N.; Johnson, C.D.; Adams, A. Writing narrative literature reviews for peer-reviewed journals: Secrets of the trade. J. Chiropr. Med. 2006, 5, 101–117. [Google Scholar] [CrossRef]
- Sewell, K.R.; Erickson, K.I.; Rainey-Smith, S.R.; Peiffer, J.J.; Sohrabi, H.R.; Brown, B.M. Relationships between physical activity, sleep and cognitive function: A narrative review. Neurosci. Biobehav. Rev. 2021, 130, 369–378. [Google Scholar] [CrossRef]
- De Dreu, M.J.; Van Der Wilk, A.S.D.; Poppe, E.; Kwakkel, G.; van Wegen, E.E. Rehabilitation, exercise therapy and music in patients with Parkinson’s disease: A meta-analysis of the effects of music-based movement therapy on walking ability, balance and quality of life. Park. Relat. Disord. 2012, 18, 114–119. [Google Scholar] [CrossRef]
- Bradt, J.; Dileo, C.; Potvin, N. Music for stress and anxiety reduction in coronary heart disease patients. Cochrane Database Syst. Rev. 2013, 12, 1–87. [Google Scholar]
- Geretsegger, M.; Elefant, C.; Mössler, K.A.; Gold, C. Music therapy for people with autism spectrum disorder. Cochrane Database Syst. Rev. 2014, 6, CD004381. [Google Scholar] [CrossRef] [PubMed]
- Maratos, A.S.; Gold, C.; Wang, X.; Crawford, M.J. Music therapy for depression. Cochrane database Syst. Rev. 2008, 1, CD004517. [Google Scholar] [CrossRef] [PubMed]
- Mössler, K.; Chen, X.; Heldal, T.O.; Gold, C. Music therapy for people with schizophrenia and schizophrenia-like disorders. Cochrane Database Syst. Rev. 2011, 12, 1–70. [Google Scholar]
- Street, A.J.; Zhang, J.; Pethers, S.; Wiffen, L.; Bond, K.; Palmer, H. Neurologic music therapy in acute stroke rehabilitation: Could it be feasible and helpful? Top. Stroke Rehabil. 2020, 27, 541–552. [Google Scholar] [CrossRef]
- Vink, A.C.; Birks, J.S.; Bruinsma, M.S.; Scholten, R.J. Music therapy for people with dementia. Cochrane Database Syst. Rev. 2004, 4, 1–65. [Google Scholar]
- O’Kelly, J.; Fachner, J.C.; Tervaniemi, M. Editorial: Dialogues in Music Therapy and Music Neuroscience: Collaborative Understanding Driving Clinical Advances. Front. Hum. Neurosci. 2016, 10, 585. [Google Scholar] [CrossRef]
- Erkkilä, J.; Punkanen, M.; Fachner, J.; Ala-Ruona, E.; Pöntiö, I.; Tervaniemi, M.; Vanhala, M.; Gold, C. Individual music therapy for depression: Randomised controlled trial. Br. J. Psychiatry 2011, 199, 132–139. [Google Scholar] [CrossRef]
- Juslin, P.N. From everyday emotions to aesthetic emotions: Towards a unified theory of musical emotions. Phys. Life Rev. 2013, 10, 235–266. [Google Scholar] [CrossRef]
- Berridge, K.C.; Kringelbach, M.L. Pleasure systems in the brain. Neuron 2015, 86, 646–664. [Google Scholar] [CrossRef]
- Koelsch, S. Brain correlates of music-evoked emotions. Nat. Rev. Neurosci. 2014, 15, 170–180. [Google Scholar] [CrossRef]
- Zatorre, R.J. Musical pleasure and reward: Mechanisms and dysfunction. Ann. N. Y. Acad. Sci. 2015, 1337, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Blood, A.J.; Zatorre, R.J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc. Natl. Acad. Sci. USA 2001, 98, 11818–11823. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Levitin, D.J. The rewards of music listening: Response and physiological connectivity of the mesolimbic system. NeuroImage 2005, 28, 175–184. [Google Scholar] [CrossRef]
- Salimpoor, V.N.; Benovoy, M.; Larcher, K.; Dagher, A.; Zatorre, R.J. Anatomically distinct dopa-mine release during anticipation and experience of peak emotion to music. Nat. Neurosci. 2011, 14, 257–262. [Google Scholar] [CrossRef]
- Salimpoor, V.N.; van den Bosch, I.; Kovacevic, N.; McIntosh, A.R.; Dagher, A.; Zatorre, R.J. Interactions between the nucleus accumbens and auditory cortices predict music reward value. Science 2013, 340, 216–219. [Google Scholar] [CrossRef]
- Ferreri, L.; Mas-Herrero, E.; Zatorre, R.J.; Ripollés, P.; Gomez-Andres, A.; Alicart, H.; Olivé, G.; Marco-Pallarés, J.; Antonijoan, R.M.; Valle, M.; et al. Dopamine modulates the reward experiences elicited by music. Proc. Natl. Acad. Sci. USA 2019, 116, 3793–3798. [Google Scholar] [CrossRef]
- Aldridge, D. The Aldridge model: An ecological systemic approach. In Music Therapy and Addictions, 1st ed.; Aldridge, D., Fachner, J., Eds.; Jessica Kingsley Publishers: London, UK, 2010; Volume 1, pp. 152–159. [Google Scholar]
- Fachner, J.; Maidhof, C.; Murtagh, D.; De Silva, D.; Pasqualitto, F.; Fernie, F.; Panin, F.; Michell, A.; Muller-Rodriguez, L.; Odell-Miller, H. Music therapy, neural processing, and craving reduction: An RCT protocol for a mixed methods feasibility study in a Community Substance Misuse Treatment Service. Addict. Sci. Clin. Pract. 2022. submitted. [Google Scholar]
- Carter, T.E.; Panisch, L.S. A systematic review of music therapy for psychosocial outcomes of substance use clients. Int. J. Ment. Health Addict. 2020, 19, 1551–1568. [Google Scholar] [CrossRef]
- Albornoz, Y. The Effects of Group Improvisational Music Therapy on Depression in Adolescents and Adults with Substance Abuse. Ph.D. Thesis, Temple University, Philadelphia, PA, USA, 2009. [Google Scholar]
- Albornoz, Y. The effects of group improvisational music therapy on depression in adolescents and adults with substance abuse: A randomized controlled trial. Nord. J. Music Ther. 2011, 20, 208–224. [Google Scholar] [CrossRef]
- Cevasco, A.M.; Kennedy, R.; Generally, N.R. Comparison of movement-to-music, rhythm activities, and competitive games on depression, stress, anxiety, and anger of females in substance abuse rehabilitation. J. Music Ther. 2005, 42, 64–80. [Google Scholar] [CrossRef]
- Silverman, M.J. Effects of music therapy on change and depression on clients in detoxification. J. Addict. Nurs. 2011, 22, 185–192. [Google Scholar] [CrossRef]
- Ghetti, C.M. Effect of music therapy with emotional-approach coping on preprocedural anxiety in cardiac catheterization: A randomized controlled trial. J. Music Ther. 2013, 50, 93–122. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Chen, T.; Wang, Z.; Chen, S.; Zhang, J.; Bao, J.; Su, H.; Tan, H.; Jiang, H.; Du, J.; et al. Effectiveness of music therapy on improving treatment motivation and emotion in female patients with methamphetamine use disorder: A randomized controlled trial. Subst. Abus. 2020, 41, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Eshaghi Farahmand, S.R.; Ahadi, H.; Kalhornia Golkar, M.; Sedaghat, M. Comparison of the Effectiveness of Music Therapy and Cognitive Behavioral Therapy on Quality of Life, Craving and Emotion Regulation in Patients Under Methadone Maintenance Therapy. Iran. J. Psychiatry Clin. Psychol. 2020, 26, 170–187. [Google Scholar] [CrossRef]
- Silverman, M.J. Effects of music therapy on change readiness and craving in patients on a detoxification unit. J. Music Ther. 2011, 48, 509–531. [Google Scholar] [CrossRef]
- Silverman, M.J. Effects of group-based educational songwriting on craving in patients on a detoxification unit: A cluster-randomized effectiveness study. Psychol. Music 2019, 47, 241–254. [Google Scholar] [CrossRef]
- Dingle, G.A.; Kelly, P.J.; Flynn, L.M.; Baker, F.A. The influence of music on emotions and cravings in clients in addiction treatment: A study of two clinical samples. Arts Psychother. 2015, 45, 18–25. [Google Scholar] [CrossRef]
- Short, A.D.; Dingle, G.A. Music as an auditory cue for emotions and cravings in adults with substance use disorders. Psychol. Music 2016, 44, 559–573. [Google Scholar] [CrossRef]
- Horesh, T. Dangerous Music-Working with the Destructive and Healing Powers of Popular Music in the Treatment of Substance Abusers. In Music and Altered States-Consciousness, Transcendence, Therapy and Addiction, 1st ed.; Aldridge, D., Fachner, J., Eds.; Jessica Kingsley Publisher: London, UK, 2006; Volume 1, pp. 125–139. [Google Scholar]
- Robb, S.L.; Carpenter, J.S.; Burns, D.S. Reporting guidelines for music-based interventions. J. Health Psychol. 2011, 16, 342–352. [Google Scholar] [CrossRef]
- Conklin, C.A.; Tiffany, S.T. Applying extinction research and theory to cue-exposure addiction treatments. Addiction 2002, 97, 155–167. [Google Scholar] [CrossRef]
- Loeber, S.; Croissant, B.; Heinz, A.; Mann, K.; Flor, H. Cue exposure in the treatment of alcohol dependence: Effects on drinking outcome, craving and self-efficacy. Br. J. Clin. Psychol. 2006, 45, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Stamou, V.; Chatzoudi, T.; Stamou, L.; Romo, L.; Graziani, P. Music-assisted systematic desensitization for the reduction of craving in response to drug-conditioned cues: A pilot study. Arts Psychother. 2016, 51, 36–45. [Google Scholar] [CrossRef]
- Stamou, V.; Clerveaux, R.; Stamou, L.; Le Rocheleuil, S.; Berejnoi, L.; Romo, L.; Graziani, P. The therapeutic contribution of music in music-assisted systematic desensitization for substance addiction treatment: A pilot study. Arts Psychother. 2017, 56, 30–44. [Google Scholar] [CrossRef]
- Bowen, H.J.; Kensinger, E.A. Recapitulation of emotional source context during memory retrieval. Cortex 2017, 91, 142–156. [Google Scholar] [CrossRef]
- Cabeza, R.; Prince, S.E.; Daselaar, S.M.; Greenberg, D.L.; Budde, M.; Dolcos, F.; LaBar, K.S.; Rubin, D.C. Brain activity during episodic retrieval of autobiographical and laboratory events: An fMRI study using a novel photo paradigm. J. Cogn. Neurosci. 2004, 16, 1583–1594. [Google Scholar] [CrossRef]
- Cabeza, R.; St Jacques, P. Functional neuroimaging of autobiographical memory. Trends Cogn. Sci. 2007, 11, 219–227. [Google Scholar] [CrossRef]
- Dolcos, F.; LaBar, K.S.; Cabeza, R. Remembering one year later: Role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proc. Natl. Acad. Sci. USA 2005, 102, 2626–2631. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, E.; McKinnon, M.C.; Levine, B. The functional neuroanatomy of autobiographical memory: A meta-analysis. Neuropsychologia 2006, 44, 2189–2208. [Google Scholar] [CrossRef]
- Koelsch, S. Towards a neural basis of music-evoked emotions. Trends Cogn. Sci. 2010, 14, 131–137. [Google Scholar] [CrossRef]
- Conway, M.A.; Rubin, D.C. The structure of autobiographical memory. In Theories of Memory, 1st ed.; Collins, A.F., Gathercole, S.E., Conway, M.A., Morris, P.E., Eds.; Lawrence Erlbaum Associates: New York, NY, USA, 2019; Volume 1, pp. 103–138. [Google Scholar]
- Renoult, L.; Tanguay, A.; Beaudry, M.; Tavakoli, P.; Rabipour, S.; Campbell, K.; Moscovitch, M.; Levine, B.; Davidson, P. Personal semantics: Is it distinct from episodic and semantic memory? An electrophysiological study of memory for autobiographical facts and repeated events in honor of Shlomo Bentin. Neuropsychologia 2016, 83, 242–256. [Google Scholar] [CrossRef]
- Ngo, C.T.; Newcombe, N.S.; Olson, I.R. The ontogeny of relational memory and pattern separation. Dev. Sci. 2018, 21, e12556. [Google Scholar] [CrossRef] [PubMed]
- Maguire, E.A.; Mummery, C.J.; Büchel, C. Patterns of hippocampal-cortical interaction dissociate temporal lobe memory subsystems. Hippocampus 2000, 10, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Renoult, L.; Davidson, P.S.; Palombo, D.J.; Moscovitch, M.; Levine, B. Personal semantics: At the crossroads of semantic and episodic memory. Trends Cogn. Sci. 2012, 16, 550–558. [Google Scholar] [CrossRef]
- Graham, K.S.; Lee, A.C.; Brett, M.; Patterson, K. The neural basis of autobiographical and semantic memory: New evidence from three PET studies. Cogn. Affect. Behav. Neurosci. 2003, 3, 234–254. [Google Scholar] [CrossRef]
- Northoff, G.; Bermpohl, F. Cortical midline structures and the self. Trends Cogn. Sci. 2004, 8, 102–107. [Google Scholar] [CrossRef]
- Greenberg, D.L.; Rice, H.J.; Cooper, J.J.; Cabeza, R.; Rubin, D.C.; LaBar, K.S. Co-activation of the amygdala, hippocampus and inferior frontal gyrus during autobiographical memory retrieval. Neuropsychologia 2005, 43, 659–674. [Google Scholar] [CrossRef] [PubMed]
- Damasio, A.R. Time-locked multiregional retroactivation: A systems-level proposal for the neural substrates of recall and recognition. Cognition 1989, 33, 25–62. [Google Scholar] [CrossRef]
- Rubin, D.C. The Basic-Systems Model of Episodic Memory. Perspect. Psychol. Sci. 2006, 1, 277–311. [Google Scholar] [CrossRef]
- Holland, A.C.; Kensinger, E.A. Emotion and autobiographical memory. Phys. Life Rev. 2010, 7, 88–131. [Google Scholar] [CrossRef]
- Ford, J.H.; Addis, D.R.; Giovanello, K.S. Differential neural activity during search of specific and general autobiographical memories elicited by musical cues. Neuropsychologia 2011, 49, 2514–2526. [Google Scholar] [CrossRef]
- Ford, J.H.; Rubin, D.C.; Giovanello, K.S. The effects of song familiarity and age on phenomenological characteristics and neural recruitment during autobiographical memory retrieval. Psychomusicology 2016, 26, 199. [Google Scholar] [CrossRef] [PubMed]
- Janata, P. The neural architecture of music-evoked autobiographical memories. Cereb. Cortex 2009, 19, 2579–2594. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, K.; Ghosh, A. Music-evoked autobiographical memories in everyday life. Psychol. Music 2021, 49, 649–666. [Google Scholar] [CrossRef]
- Janata, P.; Tomic, S.T.; Rakowski, S.K. Characterization of music-evoked autobiographical memories. Memory 2007, 15, 845–860. [Google Scholar] [CrossRef]
- Belfi, A.M.; Karlan, B.; Tranel, D. Music evokes vivid autobiographical memories. Memory 2016, 24, 979–989. [Google Scholar] [CrossRef]
- Healy, D. Psychiatric Drugs Explained, 6th ed.; Elsevier Limited: Amsterdam, The Netherlands, 2016; pp. 1–356. [Google Scholar]
- National Institute on Drug Abuse. Available online: https://www.drugabuse.gov/publications/drugs-brains-behavior-science-addiction/preface (accessed on 10 October 2022).
- Flaudias, V.; Teisseidre, F.; De Chazeron, I.; Chalmeton, M.; Bertin, C.; Izaute, M.; Chakroun-Baggioni, N.; Pereira, B.; Brousse, G.; Maurage, P. A multi-dimensional evaluation of craving and impulsivity among people admitted for alcohol-related problems in emergency department. Psychiatry Res. 2019, 272, 569–571. [Google Scholar] [CrossRef]
- Sayette, M.A.; Shiffman, S.; Tiffany, S.T.; Niaura, R.S.; Martin, C.S.; Schadel, W.G. The measurement of drug craving. Addiction 2000, 95, 189–210. [Google Scholar] [CrossRef]
- Tiffany, S.T.; Carter, B.L.; Singleton, E.G. Challenges in the manipulation, assessment and interpretation of craving relevant variables. Addiction 2000, 95, 177–187. [Google Scholar] [CrossRef]
- Brody, A.L.; Mandelkern, M.A.; London, E.D.; Childress, A.R.; Lee, G.S.; Bota, R.G.; Ho, M.L.; Saxena, S.; Baxter, L.R.; Madsen, D., Jr.; et al. Brain metabolic changes during cigarette craving. Arch. Gen. Psychiatry 2002, 59, 1162–1172. [Google Scholar] [CrossRef]
- Daglish, M.R.; Weinstein, A.; Malizia, A.L.; Wilson, S.; Melichar, J.K.; Lingford-Hughes, A.; Myles, J.S.; Grasby, P.; Nutt, D.J. Functional connectivity analysis of the neural circuits of opiate craving: “More” rather than “different”? Neuroimage 2003, 20, 1964–1970. [Google Scholar] [CrossRef]
- De Ridder, D.; Vanneste, S.; Kovacs, S.; Sunaert, S.; Dom, G. Transient alcohol craving suppression by rTMS of dorsal anterior cingulate: An fMRI and LORETA EEG study. Neurosci. Lett. 2011, 496, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Due, D.L.; Huettel, S.A.; Hall, W.G.; Rubin, D.C. Activation in mesolimbic and visuospatial neural circuits elicited by smoking cues: Evidence from functional magnetic resonance imaging. Am. J. Psychiatry 2002, 159, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.; London, E.D.; Newlin, D.B.; Villemagne, V.L.; Liu, X.; Contoreggi, C.; Phillips, R.L.; Kimes, A.S.; Margolin, A. Activation of memory circuits during cue-elicited cocaine craving. Proc. Natl. Acad. Sci. USA 1996, 93, 12040–12045. [Google Scholar] [CrossRef] [PubMed]
- Grüsser, S.M.; Wrase, J.; Klein, S.; Hermann, D.; Smolka, M.N.; Ruf, M.; Weber-Fahr, W.; Flor, H.; Mann, K.; Braus, D.F.; et al. Cue-induced activation of the striatum and medial prefrontal cortex is associated with subsequent relapse in abstinent alcoholics. Psychopharmacology 2004, 175, 296–302. [Google Scholar] [CrossRef]
- Huang, Y.; Mohan, A.; De Ridder, D.; Sunaert, S.; Vanneste, S. The neural correlates of the unified percept of alcohol-related craving: A fMRI and EEG study. Sci. Rep. 2018, 8, 923. [Google Scholar] [CrossRef]
- Kilts, C.D.; Schweitzer, J.B.; Quinn, C.K.; Gross, R.E.; Faber, T.L.; Muhammad, F.; Ely, T.D.; Hoffman, J.M.; Drexler, K.P. Neural activity related to drug craving in cocaine addiction. Arch. Gen. Psychiatry 2001, 58, 334–341. [Google Scholar] [CrossRef]
- Lou, M.; Wang, E.; Shen, Y.; Wang, J. Cue-elicited craving in heroin addicts at different abstinent time: An fMRI pilot study. Subst. Use Misuse 2012, 47, 631–639. [Google Scholar] [CrossRef]
- Maas, L.C.; Lukas, S.E.; Kaufman, M.J.; Weiss, R.D.; Daniels, S.L.; Rogers, V.W.; Kukes, T.J.; Renshaw, P.F. Functional magnetic resonance imaging of human brain activation during cue-induced cocaine craving. Am. J. Psychiatry 1998, 155, 124–126. [Google Scholar] [CrossRef]
- Myrick, H.; Anton, R.F.; Li, X.; Henderson, S.; Drobes, D.; Voronin, K.; George, M.S. Differential brain activity in alcoholics and social drinkers to alcohol cues: Relationship to craving. Neuropsychopharmacology 2004, 29, 393–402. [Google Scholar] [CrossRef]
- Sell, L.A.; Morris, J.; Bearn, J.; Frackowiak, R.S.; Friston, K.J.; Dolan, R.J. Activation of reward circuitry in human opiate addicts. Eur. J. Neurosci. 1999, 11, 1042–1048. [Google Scholar] [CrossRef]
- Seo, D.; Lacadie, C.M.; Tuit, K.; Hong, K.I.; Constable, R.T.; Sinha, R. Disrupted ventromedial prefrontal function, alcohol craving, and subsequent relapse risk. JAMA Psychiatry 2013, 70, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Smolka, M.N.; Bühler, M.; Klein, S.; Zimmermann, U.; Mann, K.; Heinz, A.; Braus, D.F. Severity of nicotine dependence modulates cue-induced brain activity in regions involved in motor preparation and imagery. Psychopharmacology 2006, 184, 577–588. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.J.; Telang, F.; Fowler, J.S.; Logan, J.; Childress, A.R.; Jayne, M.; Ma, Y.; Wong, C. Cocaine cues and dopamine in dorsal striatum: Mechanism of craving in cocaine addiction. J. Neurosci. 2006, 26, 6583–6588. [Google Scholar] [CrossRef] [PubMed]
- Wexler, B.E.; Gottschalk, C.H.; Fulbright, R.K.; Prohovnik, I.; Lacadie, C.M.; Rounsaville, B.J.; Gore, J.C. Functional magnetic resonance imaging of cocaine craving. Am. J. Psychiatry 2001, 158, 86–95. [Google Scholar] [CrossRef]
- Costa, L.; Bauer, L. Quantitative electroencephalographic differences associated with alcohol, cocaine, heroin and dual-substance dependence. Drug Alcohol Depend. 1997, 46, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Roemer, R.A.; Cornwell, A.; Dewart, D.; Jackson, P.; Ercegovac, D.V. Quantitative electroencephalographic analyses in cocaine-preferring polysubstance abusers during abstinence. Psychiatry Res. 1995, 58, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Winterer, G.; Klöppel, B.; Heinz, A.; Ziller, M.; Dufeu, P.; Schmidt, L.G.; Herrmann, W.M. Quantitative EEG (QEEG) predicts relapse in patients with chronic alcoholism and points to a frontally pronounced cerebral disturbance. Psychiatry Res. 1998, 78, 101–113. [Google Scholar] [CrossRef]
- Crockford, D.N.; Goodyear, B.; Edwards, J.; Quickfall, J.; el-Guebaly, N. Cue-induced brain activity in pathological gamblers. Biol. Psychiatry 2005, 58, 787–795. [Google Scholar] [CrossRef]
- Schneider, F.; Habel, U.; Wagner, M.; Franke, P.; Salloum, J.B.; Shah, N.J.; Toni, I.; Sulzbach, C.; Hönig, K.; Maier, W.; et al. Subcortical correlates of craving in recently abstinent alcoholic patients. Am. J. Psychiatry 2001, 158, 1075–1083. [Google Scholar] [CrossRef]
- Li, C.S.R.; Kosten, T.R.; Sinha, R. Sex differences in brain activation during stress imagery in abstinent cocaine users: A functional magnetic resonance imaging study. Biol. Psychiatry 2005, 57, 487–494. [Google Scholar] [CrossRef]
- Ray, W.J.; Cole, H.W. EEG alpha activity reflects attentional demands, and beta activity reflects emotional and cognitive processes. Science 1985, 228, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, C.S.; Strüber, D.; Helfrich, R.F.; Engel, A.K. EEG oscillations: From correlation to causality. Int. J. Psychophysiol. 2016, 103, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Polunina, A.G.; Davydov, D.M. EEG spectral power and mean frequencies in early heroin abstinence. Prog. Neuropsychopharmacol. Biol. Psychiatry 2004, 28, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Slaker, M.; Churchill, L.; Todd, R.P.; Blacktop, J.M.; Zuloaga, D.G.; Raber, J.; Darling, R.A.; Brown, T.E.; Sorg, B.A. Removal of perineuronal nets in the medial prefrontal cortex impairs the acquisition and reconsolidation of a cocaine-induced conditioned place preference memory. J. Neurosci. 2015, 35, 4190–4202. [Google Scholar] [CrossRef]
- Xue, Y.X.; Xue, L.F.; Liu, J.F.; He, J.; Deng, J.H.; Sun, S.C.; Han, H.B.; Luo, Y.X.; Xu, L.Z.; Wu, P.; et al. Depletion of perineuronal nets in the amygdala to enhance the erasure of drug memories. J. Neurosci. 2014, 34, 6647–6658. [Google Scholar] [CrossRef]
- Shulz, D.E.; Sosnik, R.; Ego, V.; Haidarliu, S.; Ahissar, E. A neuronal analogue of state-dependent learning. Nature 2000, 403, 549–553. [Google Scholar] [CrossRef]
- Müller, C.P. Episodic memories and their relevance for psychoactive drug use and addiction. Front. Behav. Neurosci. 2013, 7, 34. [Google Scholar] [CrossRef]
- O’Brien, C.P.; Childress, A.R.; Ehrman, R.; Robbins, S.J. Conditioning factors in drug abuse: Can they explain compulsion? J. Psychopharmacol. 1998, 12, 15–22. [Google Scholar] [CrossRef]
- Robbins, T.W.; Ersche, K.D.; Everitt, B.J. Drug addiction and the memory systems of the brain. Ann. N. Y. Acad. Sci. 2008, 1141, 1–21. [Google Scholar] [CrossRef]
- Abel, T.; Lattal, K.M. Molecular mechanisms of memory acquisition, consolidation and retrieval. Curr. Opin. Neurobiol. 2001, 11, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R.; Schwartz, J.H.; Jessell, T.M.; Siegelbaum, S.A.; Hudspeth, A.J. Principles of Neural Science, 5th ed.; McGraw-Hill Education: New York, NY, USA, 2013; pp. 1–1448. [Google Scholar]
- Alberini, C.M. Mechanisms of memory stabilization: Are consolidation and reconsolidation similar or distinct processes? Trends Neurosci. 2005, 28, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.L.; Everitt, B.J.; Thomas, K.L. Independent cellular processes for hippocampal memory consolidation and reconsolidation. Science 2004, 304, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Artinian, J.; McGauran, A.M.T.; De Jaeger, X.; Mouledous, L.; Frances, B.; Roullet, P. Protein degradation, as with protein synthesis, is required during not only long-term spatial memory consolidation but also reconsolidation. Eur. J. Neurosci. 2008, 27, 3009–3019. [Google Scholar] [CrossRef] [PubMed]
- Bender, B.N.; Torregrossa, M.M. Molecular and circuit mechanisms regulating cocaine memory. Cell. Mol. Life Sci. 2020, 77, 3745–3768. [Google Scholar] [CrossRef] [PubMed]
- Kaang, B.K.; Lee, S.H.; Kim, H. Synaptic protein degradation as a mechanism in memory reorganization. Neuroscientist 2009, 15, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Choi, J.H.; Lee, N.; Lee, H.R.; Kim, J.I.; Yu, N.K.; Choi, S.L.; Kim, H.; Kaang, B.K. Synaptic protein degradation underlies destabilization of retrieved fear memory. Science 2008, 319, 1253–1256. [Google Scholar] [CrossRef]
- McClung, C.A.; Ulery, P.G.; Perrotti, L.I.; Zachariou, V.; Berton, O.; Nestler, E.J. DeltaFosB: A molecular switch for long-term adaptation in the brain. Brain Res. Mol. Brain Res. 2004, 132, 146–154. [Google Scholar] [CrossRef]
- Milton, A.L.; Everitt, B.J. The psychological and neurochemical mechanisms of drug memory reconsolidation: Implications for the treatment of addiction. Eur. J. Neurosci. 2010, 31, 2308–2319. [Google Scholar] [CrossRef]
- Nestler, E.J. Molecular mechanisms of drug addiction. Neuropharmacology 2004, 47, 24–32. [Google Scholar] [CrossRef]
- Fawcett, J.W.; Oohashi, T.; Pizzorusso, T. The roles of perineuronal nets and the perinodal extracellular matrix in neuronal function. Nat. Rev. Neurosci. 2019, 20, 451–465. [Google Scholar] [CrossRef]
- Gogolla, N.; Caroni, P.; Lüthi, A.; Herry, C. Perineuronal nets protect fear memories from erasure. Science 2009, 325, 1258–1261. [Google Scholar] [CrossRef] [PubMed]
- LeDoux, J.E. Emotion circuits in the brain. Ann. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef]
- Myers, K.M.; Davis, M. Mechanisms of fear extinction. Mol. Psychiatry 2007, 12, 120–150. [Google Scholar]
- Kim, J.H.; McNally, G.P.; Richardson, R. Recovery of fear memories in rats: Role of gamma-amino butyric acid (GABA) in infantile amnesia. Behav. Neurosci. 2006, 120, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Richardson, R. The effect of temporary amygdala inactivation on extinction and reextinction of fear in the developing rat: Unlearning as a potential mechanism for extinction early in development. J. Neurosci. 2008, 28, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Slaker, M.L.; Jorgensen, E.T.; Hegarty, D.M.; Liu, X.; Kong, Y.; Zhang, F.; Linhardt, R.J.; Brown, T.E.; Aicher, S.A.; Sorg, B.A. Cocaine Exposure Modulates Perineuronal Nets and Synaptic Excitability of Fast-Spiking Interneurons in the Medial Prefrontal Cortex. eNeuro 2018, 5, 1–17. [Google Scholar] [CrossRef]
- Prus, A.J.; James, J.R.; Rosecrans, J.A. Conditioned Place Preference. In Methods of Behavior Analysis in Neuroscience, 2nd ed.; Buccafusco, J.J., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2009; pp. 1–343. [Google Scholar]
- De Winter, F.; Kwok, J.C.F.; Fawcett, J.W.; Vo, T.T.; Carulli, D.; Verhaagen, J. The chemorepulsive protein semaphorin 3A and perineuronal net-mediated plasticity. Neural. Plast. 2016, 2016, 3679545. [Google Scholar] [CrossRef]
- Boyd, J.; George-Warren, H. Musicians in Tune: Seventy-Five Contemporary Musicians Discuss the Creative Process; Fireside, a Simon & Schuster Imprimateur: New York, NY, USA, 1992; pp. 1–288. [Google Scholar]
- Silverman, M.J.; Bourdaghs, S.; Abbazio, J.; Riegelman, A. A systematic review of music-induced substance craving. Music Sci. 2021, 10298649211030314. [Google Scholar] [CrossRef]
- Chen, J.Y.; Cao, J.P.; Wang, Y.C.; Li, S.Q.; Wang, Z.Z. A New Measure for Assessing the Intensity of Addiction Memory in Illicit Drug Users: The Addiction Memory Intensity Scale. J. Clin. Med. 2018, 7, 467. [Google Scholar] [CrossRef]
- Shi, H.S.; Luo, Y.X.; Xue, Y.X.; Wu, P.; Zhu, W.L.; Ding, Z.B.; Lu, L. Effects of sleep deprivation on retrieval and reconsolidation of morphine reward memory in rats. Pharmacol. Biochem. Behav. 2011, 98, 299–303. [Google Scholar] [CrossRef]
- Favuzzi, E.; Marques-Smith, A.; Deogracias, R.; Winterflood, C.M.; Sánchez-Aguilera, A.; Mantoan, L.; Maeso, P.; Fernandes, C.; Ewers, H.; Rico, B. Activity-Dependent Gating of Parvalbumin Interneuron Function by the Perineuronal Net Protein Brevican. Neuron 2017, 95, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Sale, A.; Maya Vetencourt, J.F.; Medini, P.; Cenni, M.C.; Baroncelli, L.; De Pasquale, R.; Maffei, L. Environmental enrichment in adulthood promotes amblyopia recovery through a reduction of intracortical inhibition. Nat. Neurosci. 2007, 10, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Alvin, J. Music Therapy, 1st ed.; Stainer & Bell Ltd.: London, UK, 1966; pp. 1–192. [Google Scholar]
- Fachner, J.C.; Maidhof, C.; Grocke, D.; Nygaard Pedersen, I.; Trondalen, G.; Tucek, G.; Bonde, L.O. “Telling me not to worry…” Hyperscanning and Neural Dynamics of Emotion Processing During Guided Imagery and Music. Front. Psychol. 2019, 10, 1561. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, T.; Hou, Y.; Pan, Y.; Hu, Y. Interpersonal brain synchronization associated with working alliance during psychological counseling. Psychiatry Res. Neuroimaging 2018, 282, 103–109. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Brain Region | BA | Domain Involved | Main Findings |
|---|---|---|---|
| dlPFC | 46 | AM | ↑ activation during AM recollection [66] |
| 46 | MEAM | ↑ activation during familiar and popular music triggering AM [82,84] | |
| 46 | Craving | ↑ activation during the presentation of drug-related cues [101] | |
| N.A. * | PNNs | Causal role in acquisition and reconsolidation of addiction memories in preclinical study [118] | |
| vmPFC | 9/10 | AM | ↑ activation during AM recollection [76] |
| 25/11 | MEAM | ↑ activation during familiar and popular music triggering AM [82] | |
| 11/32/25 | Craving | ↑ activation during the presentation of drug-related cues [105] | |
| mPFC | 10/32 | AM | ↑ activation during AM recollection [66] |
| 8/9/10/11/32 | MEAM | ↑ activation during familiar and popular music triggering AM [82,84] | |
| 9/10 | Craving | ↑ activation during the presentation of drug-related cues [98] | |
| OFC | 11 | AM | ↑ activation during emotional content recollection [65] |
| 11 | MEAM | ↑ activation during familiar and popular music triggering AM [82] | |
| 11/47 | Craving | ↑ activation during the presentation of drug-related cues [93,94] | |
| PCC | 23/29/30/31 | AM | ↑ activation during AM recollection [66,69] |
| 30/31 | MEAM | ↑ activation during familiar and popular music triggering AM [82] | |
| 31 | Craving | ↑ activation during the presentation of drug-related cues [114] | |
| Amygdala | N.A. | AM | ↑ activation during emotional content recollection [65,69] |
| N.A. | MEAM | ↑ activation during familiar and popular music triggering AM [82] | |
| N.A. | Craving | ↑ activation during the presentation of drug-related cues [97,101] | |
| N.A. * | PNNs | Causal role in acquisition and reconsolidation of addiction memories in preclinical study [119] | |
| Hippocampus | N.A. | AM | ↑ activation during AM recollection [66] |
| N.A. | MEAM | ↑ activation during familiar and popular music triggering AM [82] | |
| N.A. * | Craving | ↑ activation during the presentation of drug-related cues [97,100,101,106,112,113] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasqualitto, F.; Panin, F.; Maidhof, C.; Thompson, N.; Fachner, J. Neuroplastic Changes in Addiction Memory—How Music Therapy and Music-Based Intervention May Reduce Craving: A Narrative Review. Brain Sci. 2023, 13, 259. https://doi.org/10.3390/brainsci13020259
Pasqualitto F, Panin F, Maidhof C, Thompson N, Fachner J. Neuroplastic Changes in Addiction Memory—How Music Therapy and Music-Based Intervention May Reduce Craving: A Narrative Review. Brain Sciences. 2023; 13(2):259. https://doi.org/10.3390/brainsci13020259
Chicago/Turabian StylePasqualitto, Filippo, Francesca Panin, Clemens Maidhof, Naomi Thompson, and Jörg Fachner. 2023. "Neuroplastic Changes in Addiction Memory—How Music Therapy and Music-Based Intervention May Reduce Craving: A Narrative Review" Brain Sciences 13, no. 2: 259. https://doi.org/10.3390/brainsci13020259
APA StylePasqualitto, F., Panin, F., Maidhof, C., Thompson, N., & Fachner, J. (2023). Neuroplastic Changes in Addiction Memory—How Music Therapy and Music-Based Intervention May Reduce Craving: A Narrative Review. Brain Sciences, 13(2), 259. https://doi.org/10.3390/brainsci13020259