
Reversal Training Discloses Gender Differences in a Spatial Memory Task in Humans



Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Apparatus
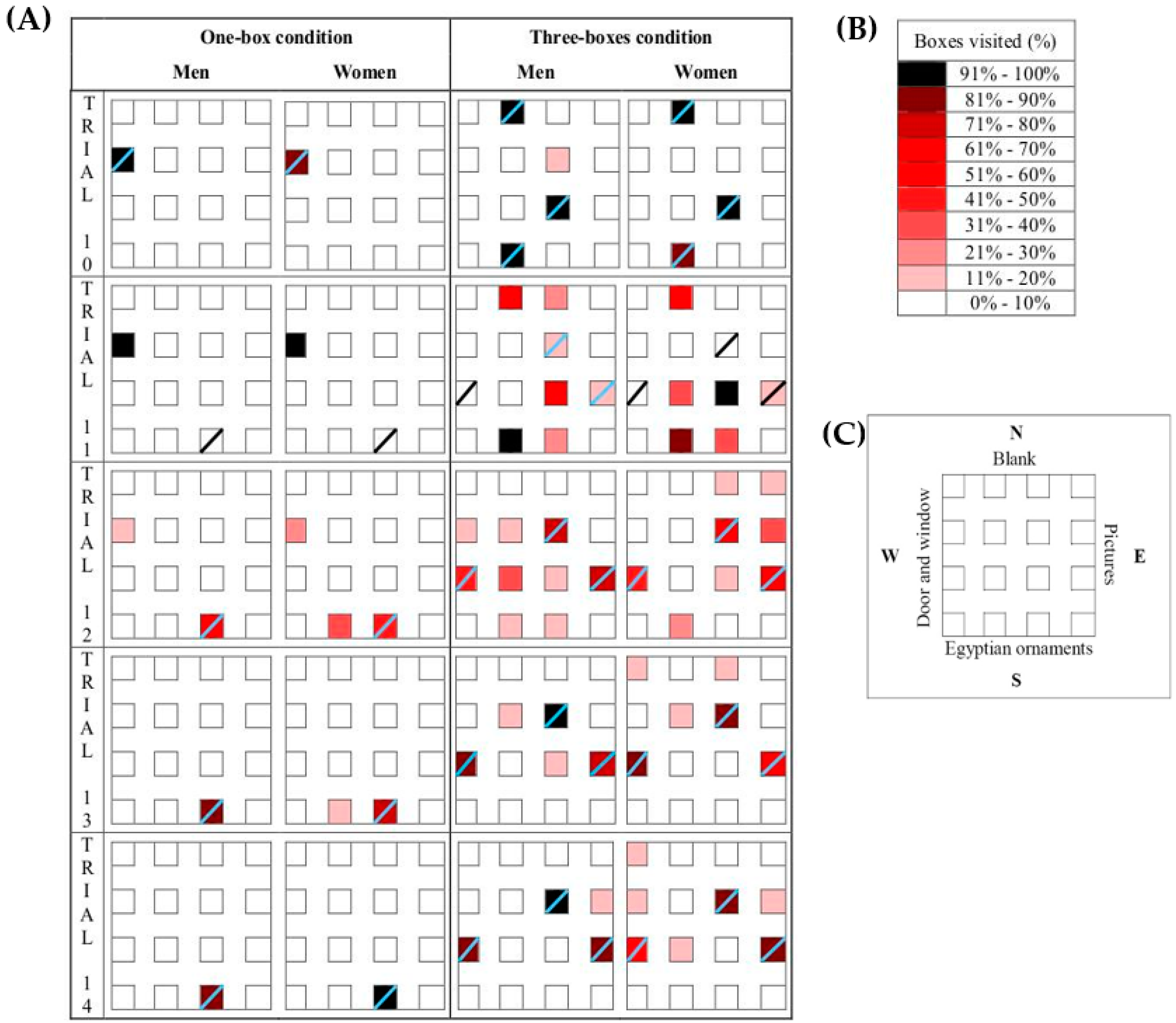
2.3. Procedure
2.4. Statistical Analyses
3. Results
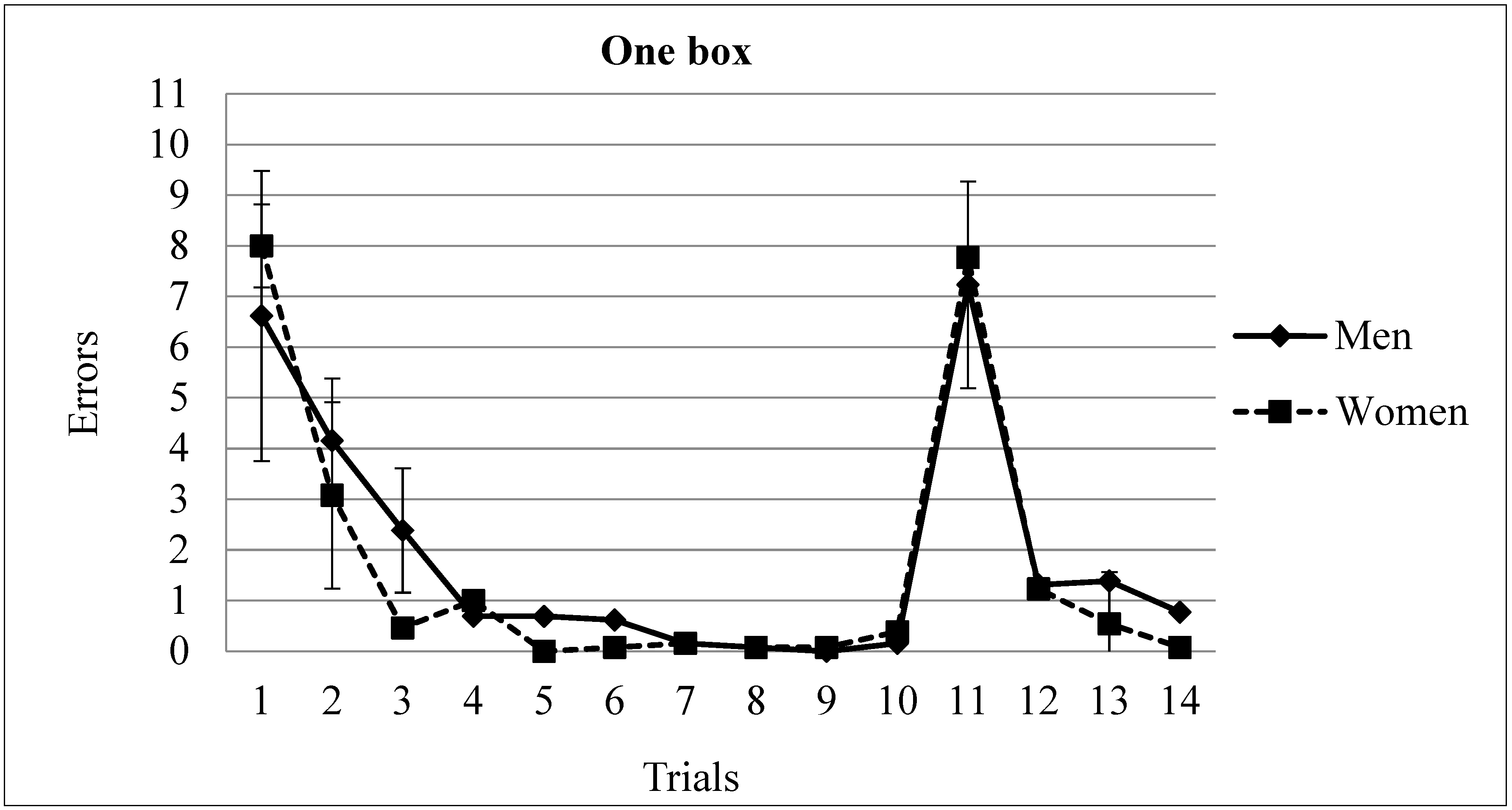
3.1. One Rewarded Box
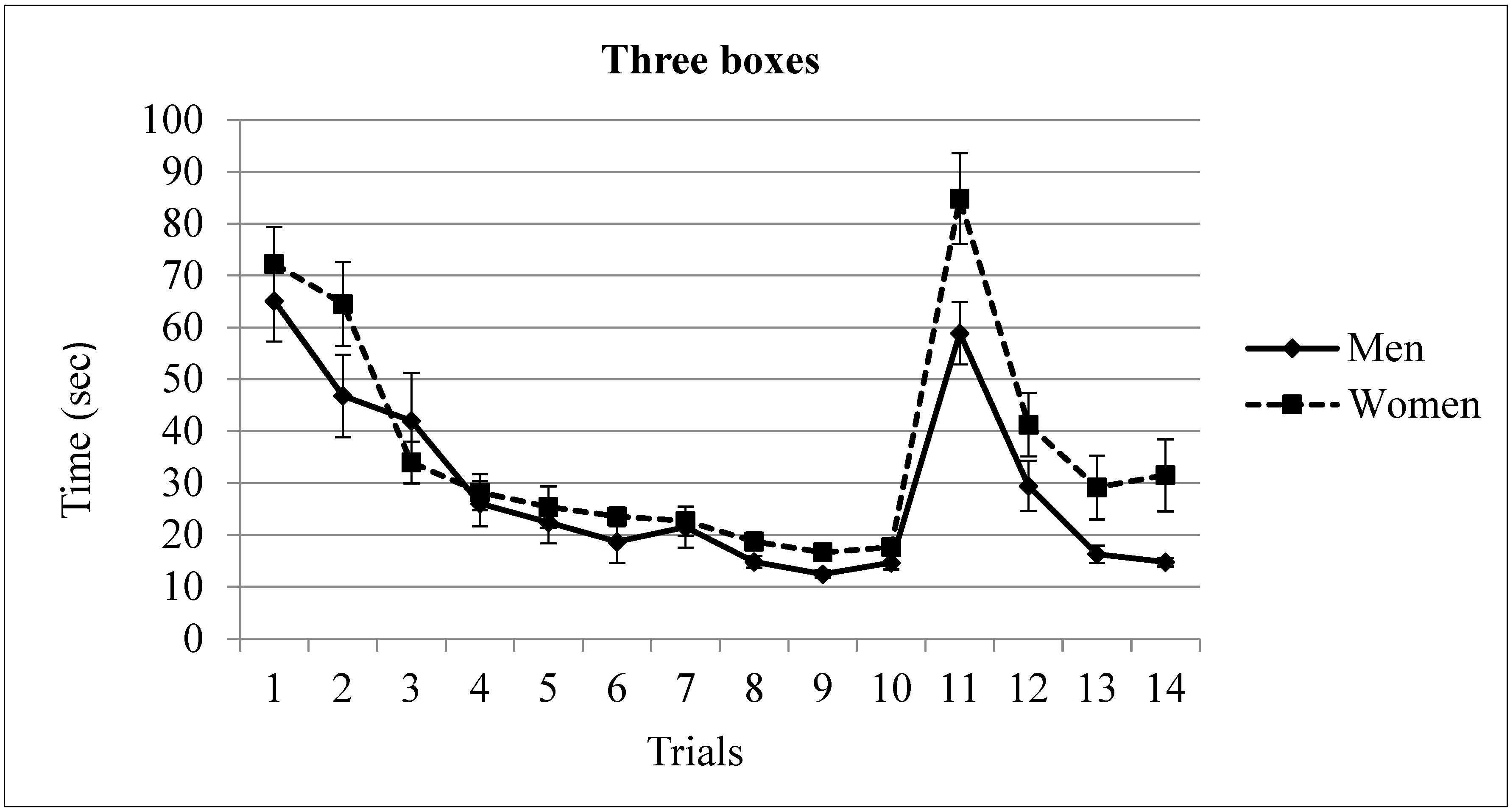
3.2. Three Rewarded Boxes
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madl, T.; Chen, K.; Montaldi, D.; Trappl, R. Computational cognitive models of spatial memory in navigation space: A review. Neural Netw. 2015, 65, 18–43. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Dostrovsky, J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Herweg, N.A.; Kahana, M.J. Spatial Representations in the Human Brain. Front. Hum. Neurosci. 2018, 12, 297. [Google Scholar] [CrossRef]
- Fritch, H.A.; Spets, D.S.; Slotnick, S.D. Functional connectivity with the anterior and posterior hippocampus during spatial memory. Hippocampus 2021, 31, 658–668. [Google Scholar] [CrossRef]
- Uddin, L.Q. Cognitive and behavioural flexibility: Neural mechanisms and clinical considerations. Nat. Rev. Neurosci. 2021, 22, 167–179. [Google Scholar] [CrossRef]
- Berg, E.A. A Simple Objective Technique for Measuring Flexibility in Thinking. J. Gen. Psychol. 1948, 39, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.M.; Rubin, R.B. A New Measure of Cognitive Flexibility. Psychol. Rep. 1995, 76, 623–626. [Google Scholar] [CrossRef]
- Dennis, J.P.; Wal, J.S.V. The Cognitive Flexibility Inventory: Instrument Development and Estimates of Reliability and Validity. Cogn. Ther. Res. 2009, 34, 241–253. [Google Scholar] [CrossRef]
- Colman, A.M. A Dictionary of Psychology, 4th ed.; Oxford University Express: Oxford, UK, 2015. [Google Scholar]
- Schrijver, N.C.; Pallier, P.N.; Brown, V.J.; Würbel, H. Double dissociation of social and environmental stimulation on spatial learning and reversal learning in rats. Behav. Brain Res. 2004, 152, 307–314. [Google Scholar] [CrossRef]
- Gatewood, J.D.; Morgan, M.D.; Eaton, M.; McNamara, I.M.; Stevens, L.F.; Macbeth, A.H.; Meyer, E.A.A.; Lomas, L.M.; Kozub, F.J.; Lambert, K.G.; et al. Motherhood mitigates aging-related decrements in learning and memory and positively affects brain aging in the rat. Brain Res. Bull. 2005, 66, 91–98. [Google Scholar] [CrossRef]
- Watson, D.J.; Stanton, M.E. Intrahippocampal administration of an NMDA-receptor antagonist impairs spatial discrimination reversal learning in weanling rats. Neurobiol. Learn. Mem. 2009, 92, 89–98. [Google Scholar] [CrossRef]
- Dong, Z.; Bai, Y.; Wu, X.; Li, H.; Gong, B.; Howland, J.G.; Huang, Y.; He, W.; Li, T.; Wang, Y.T. Hippocampal long-term depression mediates spatial reversal learning in the Morris water maze. Neuropharmacology 2013, 64, 65–73. [Google Scholar] [CrossRef]
- Hyer, M.; Wegener, A.; Targett, I.; Dyer, S.; Neigh, G. Chronic stress beginning in adolescence decreases spatial memory following an acute inflammatory challenge in adulthood. Behav. Brain Res. 2023, 442, 114323. [Google Scholar] [CrossRef] [PubMed]
- Avigan, P.D.; Cammack, K.; Shapiro, M.L. Flexible spatial learning requires both the dorsal and ventral hippocampus and their functional interactions with the prefrontal cortex. Hippocampus 2020, 30, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Thonnard, D.; Callaerts-Vegh, Z.; D’hooge, R. Effects of orbitofrontal cortex and ventral hippocampus disconnection on spatial reversal learning. Neurosci. Lett. 2021, 750, 135711. [Google Scholar] [CrossRef]
- Izquierdo, A.; Jentsch, D. Reversal learning as a measure of impulsive and compulsive behavior in addictions. Psychofarmachology 2012, 219, 607–620. [Google Scholar] [CrossRef]
- Evans, K.L.; Hampson, E. Sex differences on prefrontally-dependent cognitive tasks. Brain Cogn. 2015, 93, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.F.; Sturz, B.R.; Andriole, K.; Hardesty, S.; Place, M. Precedence of spatial pattern learning revealed by immediate reversal performance. Behav. Process. 2010, 85, 252–264. [Google Scholar] [CrossRef]
- Schoenfeld, R.; Foreman, N.; Leplow, B. Ageing and spatial reversal learning in humans: Findings from a virtual water maze. Behav. Brain Res. 2014, 270, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Antonova, E.; Parslow, D.; Brammer, M.; Simmons, A.; Williams, S.; Dawson, G.; Morris, R. Scopolamine disrupts hippocampal activity during allocentric spatial memory in humans: An fMRI study using a virtual reality analogue of the Morris Water Maze. J. Psychopharmacol. 2011, 25, 1256–1265. [Google Scholar] [CrossRef]
- Danjo, T. Allocentric representations of space in the hippocampus. Neurosci. Res. 2020, 153, 1–7. [Google Scholar] [CrossRef]
- Ramos, J.M.; Morón, I. Ventral hippocampus lesions and allocentric spatial memory in the radial maze: Anterograde and retrograde deficits. Behav. Brain Res. 2021, 417, 113620. [Google Scholar] [CrossRef]
- Astur, R.S.; Ortiz, M.L.; Sutherland, R.J. A characterization of performance by men and women in a virtual Morris water task: A large and reliable sex difference. Behav. Brain Res. 1998, 93, 185–190. [Google Scholar] [CrossRef]
- Cimadevilla, J.M.; Fenton, A.A.; Bures, J. Transient sex differences in the between-sessions but not in the within-session memory underlying an active place avoidance task in weanling rats. Behav. Neurosci. 2001, 115, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Lövdén, M.; Herlitz, A.; Schellenbach, M.; Grossman-Hutter, B.; Krüger, A.; Lindenberger, U. Quantitative and qualitative sex differences in spatial navigation. Scand. J. Psychol. 2007, 48, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Cimadevilla, J.M.; Cánovas, R.; Iribarne, L.; Soria, A.; López, L. A virtual-based task to assess place avoidance in humans. J. Neurosci. Methods 2011, 196, 45–50. [Google Scholar] [CrossRef] [PubMed]
- León, I.; Tascón, L.; Cimadevilla, J.M. Age and gender-related differences in a spatial memory task in humans. Behav. Brain Res. 2016, 306, 8–12. [Google Scholar] [CrossRef]
- Tascón, L.; García-Moreno, L.M.; Cimadevilla, J.M. Almeria spatial memory recognition task (ASMRT): Gender differences emerged in a new passive spatial task. Neurosci. Lett. 2017, 651, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Tascón, L.; Di Cicco, C.; Piccardi, L.; Palmiero, M.; Bocchi, A.; Cimadevilla, J.M. Sex Differences in Spatial Memory: Comparison of Three Tasks Using the Same Virtual Context. Brain Sci. 2021, 11, 757. [Google Scholar] [CrossRef]
- Chen, W.; Liu, B.; Li, X.; Wang, P.; Wang, B. Sex Differences in Spatial Memory. Neuroscience 2020, 443, 140–147. [Google Scholar] [CrossRef]
- Chai, X.J.; Jacobs, L.F. Effects of cue types on sex differences in human spatial memory. Behav. Brain Res. 2010, 208, 336–342. [Google Scholar] [CrossRef]
- Piccardi, L.; Risetti, M.; Nori, R.; Tanzilli, A.; Bernardi, L.; Guariglia, C. Perspective changing in primary and secondary learning: A gender difference study. Learn. Individ. Differ. 2011, 21, 114–118. [Google Scholar] [CrossRef]
- Palermo, L.; Iaria, G.; Guariglia, C. Mental imagery skills and topographical orientation in humans: A correlation study. Behav. Brain Res. 2008, 192, 248–253. [Google Scholar] [CrossRef]
- Voyer, D.; Saint-Aubin, J.; Altman, K.; Doyle, R.A. Sex differences in tests of mental rotation: Direct manipulation of strategies with eye-tracking. J. Exp. Psychol. Hum. Percept. Perform. 2020, 46, 871–889. [Google Scholar] [CrossRef]
- Dabbs, J.M.; Chang, E.-L.; A Strong, R.; Milun, R. Spatial Ability, Navigation Strategy, and Geographic Knowledge Among Men and Women. Evol. Hum. Behav. 1998, 19, 89–98. [Google Scholar] [CrossRef]
- Andreano, J.M.; Cahill, L. Sex influences on the neurobiology of learning and memory. Learn. Mem. 2009, 16, 248–266. [Google Scholar] [CrossRef] [PubMed]
- Van den Bos, R.; Homberg, J.; de Visser, L. A critical review of sex differences in decision making tasks: Focus on the Iowan Glambing Task. Behav. Brain Res. 2013, 238, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Coluccia, E.; Louse, G. Gender differences in spatial orientation: A review. J. Environ. Psychol. 2004, 24, 329–340. [Google Scholar] [CrossRef]
- Nori, R.; Piccardi, L.; Migliori, M.; Guidazzoli, A.; Frasca, F.; De Luca, D.; Giusberti, F. The virtual reality Walking Corsi Test. Comput. Hum. Behav. 2015, 48, 72–77. [Google Scholar] [CrossRef]
- Cánovas, R.; Espínola, M.; Iribarne, L.; Cimadevilla, J.M. A new virtual task to evaluate human place learning. Behav. Brain Res. 2008, 190, 112–118. [Google Scholar] [CrossRef]
- Parsons, T.D.; Rizzo, A.R.; Van Der Zaag, C.; McGee, J.S.; Buckwalter, J.G. Gender Differences and Cognition Among Older Adults. Aging Neuropsychol. Cogn. 2005, 12, 78–88. [Google Scholar] [CrossRef]
- León, I.; Cimadevilla, J.M.; Tascón, L. Developmental gender differences in children in a virtual spatial memory task. Neuropsychology 2014, 28, 485–495. [Google Scholar] [CrossRef]
- Cánovas, R.; León, I.; Serrano-Castro, P.J.; Roldán, M.D.; Cimadevilla, J.M. Spatial navigation impairment in patients with refractory temporal lobe epilepsy: Evidence from a new virtual reality-based task. Epilepsy Behav. 2011, 22, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, R.; Cimadevilla, J.M. Sexual orientation and spatial memory. Psichothema 2011, 23, 752–758. [Google Scholar]
- Overman, W.H. Sex differences in early childhood, adolescence and adulthood on cognitive tasks that rely on orbital prefrontal cortex. Brain Cogn. 2003, 55, 134–147. [Google Scholar] [CrossRef]
- Davis, S.J.C.; Coltheart, M. Rehabilitation of topographical disorientation: An experimental single case study. Neuropsychol. Rehab. 1999, 9, 1–30. [Google Scholar] [CrossRef]
- Brunsdon, R.; Nickels, L.; Coltheart, M. Topographical disorientation: Towards an integrated framework for assessment. Neuropsychol. Rehabil. 2007, 17, 34–52. [Google Scholar] [CrossRef]
- Goldberg, E. The Executive Brain. In Frontal Lobes and Civilized Mind, 1st ed.; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Nori, R.; Grandicelli, S.; Giusberti, F. Aligment effect: Primary-secondary learning and cognitive styles. Perception 2006, 35, 1233–1249. [Google Scholar] [CrossRef]
- Iachini, T.; Sergi, I.; Ruggiero, G.; Gnisci, A. Gender differences in object location memory in a real three-dimensional environment. Brain Cogn. 2005, 59, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Escamilla, J.C.; Castro, J.J.F.; Baliyan, S.; Ortells-Pareja, J.J.; Rodríguez, J.J.O.; Cimadevilla, J.M. Allocentric Spatial Memory Performance in a Virtual Reality-Based Task is Conditioned by Visuospatial Working Memory Capacity. Brain Sci. 2020, 10, 552. [Google Scholar] [CrossRef]
- Wolbers, T.; Hegarty, M. What determines our navigational abilities? Trends Cogn. Sci. 2010, 14, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, R.; Fernández-García, R.; Cimadevilla, J.M. Effect of reference frames and number of cues available on the spatial orientation of males and females in a virtual memory task. Behav. Brain Res. 2011, 216, 116–121. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Low Difficulty | High Difficulty |
|---|---|---|
| Men (n = 30) | 13 | 17 |
| Age | 20.54 ± 2.96 | 21.59 ± 5.48 |
| Women (n = 30) | 13 | 17 |
| Age | 18.54 ± 0.88 | 20.76 ± 3.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tascón, L.; León, I.; Fernández, R.; Cimadevilla, J.M. Reversal Training Discloses Gender Differences in a Spatial Memory Task in Humans. Brain Sci. 2023, 13, 740. https://doi.org/10.3390/brainsci13050740
Tascón L, León I, Fernández R, Cimadevilla JM. Reversal Training Discloses Gender Differences in a Spatial Memory Task in Humans. Brain Sciences. 2023; 13(5):740. https://doi.org/10.3390/brainsci13050740
Chicago/Turabian StyleTascón, Laura, Irene León, Rubén Fernández, and José Manuel Cimadevilla. 2023. "Reversal Training Discloses Gender Differences in a Spatial Memory Task in Humans" Brain Sciences 13, no. 5: 740. https://doi.org/10.3390/brainsci13050740
APA StyleTascón, L., León, I., Fernández, R., & Cimadevilla, J. M. (2023). Reversal Training Discloses Gender Differences in a Spatial Memory Task in Humans. Brain Sciences, 13(5), 740. https://doi.org/10.3390/brainsci13050740