1. Introduction
Glioblastoma is the most aggressive form of glioma, categorized as grade 4 [
1]. Its annual incidence is approximately four cases per 100,000 people [
2,
3]. Despite current treatment modalities, which include surgical resection, radiation therapy, and chemotherapy with temozolomide (TMZ) [
4,
5], this brain tumor has an exceedingly dismal prognosis. The median survival rate of patients with this malignancy does not surpass 1.5 years [
2], and only a small fraction of patients, roughly 7%, survive beyond five years following diagnosis [
2]. Given the extremely unfavorable prognosis for patients with glioblastoma, extensive research is underway to better understand the tumor mechanisms of this disease and develop novel therapeutic approaches. One of the areas of investigation in glioblastoma research pertains to fatty acid metabolism within the tumor.
Fatty acids represent a fundamental component of both cell and intracellular membranes, as well as a crucial source of energy and a precursor to the production of lipid mediators. The synthesis of fatty acids can occur de novo or, in the case of essential polyunsaturated fatty acids (PUFA), through the elongation and desaturation of other PUFAs [
6]. The uptake of fatty acids by cells is facilitated by a variety of transporters, including the cluster of differentiation 36 (CD36) [
7] and solute carrier family 27 (SLC27) [
8]. For fatty acids to be utilized in cellular metabolism, they must first be activated by conversion to a fatty acyl coenzyme A (CoA) through the catalytic activity of acyl-CoA synthetases [
8]. The SLC27A family comprises six members, denoted as SLC27A1 to SLC27A6 [
8]. Previously, these proteins were referred to as very-long-chain acyl-CoA synthetases (ACSVL) or fatty acid transport proteins (FATP). The primary function of these enzymes is to catalyze the conversion of unesterified fatty acids to fatty acyl-CoA, while also facilitating the transport of fatty acids across the cell membrane. It is important to note that these enzymes must be localized in the cell membrane to carry out their transport function directly. Another important feature of acyl-CoA synthetases is the channeling of fatty acids [
8], whereby a specific fatty acid is directed towards a particular metabolic pathway by a corresponding acyl-CoA synthetase, dependent on protein–protein interactions and subcellular localization of the synthetase [
9].
SLC27A1/ACSVL5/FATP1 exhibits activity towards saturated fatty acids (SFA), particularly palmitic acid C16:0, as well as a much higher activity towards lignoceric acid C24:0 [
10,
11,
12]. It can also activate arachidonic acid C20:4n-6 [
13]. SLC27A1 is responsible for fatty acid transport [
14]. SLC27A1 is located in the endoplasmic reticulum [
15] and plays a role in triglyceride and ceramide synthesis [
9,
16,
17,
18]. It also directs fatty acids towards β-oxidation [
18,
19]. There is high expression of SLC27A1 in the brain [
20,
21] where it participates in the transport of fatty acids across the blood–brain barrier (BBB) in conjunction with SLC27A4 [
22], with a particular role in the transport of docosahexaenoic acid (DHA) C22:6n-3 [
23,
24].
SLC27A2/ACSVL1/FATP2 is an enzyme that activates SFAs, including palmitic acid C16:0 and lignoceric acid C24:0 [
25], as well as arachidonic acid C20:4n-6 [
13]. SLC27A2 in peroxisomes plays a role in the β-oxidation of very-long-chain fatty acids [
26] and contributes to the synthesis of glycerophospholipids, specifically in the incorporation of arachidonic acid C20:4n-6 and DHA C22:6n-3 [
27]. Additionally, SLC27A2 is involved in ceramide synthesis [
9] and is responsible for fatty acid uptake [
13,
28]. In the brain, SLC27A2 is expressed at very low levels [
21,
29,
30].
SLC27A3/ACSVL3/FATP3 is an enzyme that activates SFA, namely stearic acid C18:0, behenic acid C22:0, and lignoceric acid C24:0 [
31,
32]. SLC27A3 participates in the synthesis of ceramides, as demonstrated by experiments on U87 MG cells [
32]. Notably, SLC27A3 does not play a role in fatty acid uptake [
13]. Furthermore, SLC27A3 is involved in the tumorigenesis of glioblastomas, particularly in the functioning of glioblastoma stem cells [
33]. However, expression of SLC27A3 in the brain is very low [
21,
31].
SLC27A4/ACSVL4/FATP4 demonstrates activity towards palmitic acid C16:0 and significantly greater activity towards lignoceric acid C24:0 [
12,
34]. It also activates arachidonic acid C20:4n-6 [
13]. SLC27A4 is responsible for fatty acid uptake [
13,
28,
35]. This protein participates in the production of triglycerides [
17,
36], cholesterol esters [
36], and ceramide [
9]. SLC27A4 is highly expressed in the brain [
21,
34]. In the brain, SLC27A1 and SLC27A4 work together in the transport of fatty acids through the BBB [
22].
SLC27A5/ACSVL6/FATP5 shows activity towards very-long-chain SFA, particularly stearic acid C18:0, and cerotic acid C26:0 [
37]. Additionally, this enzyme displays bile acid-CoA ligase activity [
38]. SLC27A5 is localized in the cell membrane and is responsible for fatty acid uptake, resulting in the production of triglycerides, glycerophospholipids, and cholesteryl esters [
39]. The expression of SLC27A5 is very low in the brain [
21,
39].
SLC27A6/ACSVL2/FATP6 is a cardiac-specific isoform [
40]. SLC27A6 activates arachidonic acid C20:4n-6 and exhibits weak very-long-chain acyl-CoA synthetase activity towards lignoceric acid C24:0 [
13]. This protein preferentially transports palmitic acid C16:0 [
40]. The expression of SLC27A6 is very low in the brain [
21].
Recent research has shown that long-chain and very-long-chain fatty acids can initiate signal transduction by interacting with receptors or transporters. One example is CD36, which can activate a number of signaling pathways, including cAMP, Ca
2+, Src (which activates phospholipase C (PLC), liver kinase B1 (LKB1), and insulin receptor β (IRβ)), ERK MAPK, vascular endothelial growth factor receptor 2 (VEGFR2), and peroxisome proliferator-activated receptor δ (PPARδ) [
41]. Furthermore, long-chain and very-long-chain fatty acids can function as ligands for G protein-coupled receptors, including free fatty acid receptor (FFAR)1/G protein-coupled receptor (GPR)40 and FFAR4/GPR120 [
41,
42]. FFAR1/GPR40 is associated with G
q, G
i, and G
s, while FFAR4/GPR120 only with G
q. However, according to data from the GEPIA portal, these receptors do not appear to play significant roles in glioblastoma cancer processes. Specifically, the expression levels of FFAR1/GPR40 and FFAR4/GPR120 do not differ significantly between GBM tumors and healthy brain tissue, nor are they significantly associated with patient prognosis [
43].
Acyl-CoA synthetases can trigger signal transduction upon activation by fatty acids, utilizing unesterified fatty acids, free coenzyme A (CoASH), and ATP as substrates, and producing fatty acyl-CoA and AMP as products [
44,
45]. AMP functions as a second messenger and is capable of activating AMP-activated protein kinase (AMPK), thereby participating in signal transduction [
45].
In our previous investigations, we examined the expression and function of desaturases [
46] and elongases [
47] in glioblastoma. The findings of our research provided novel insights into the fatty acid biosynthesis within glioblastoma tumors. Nevertheless, the lipid metabolism in this malignancy remains insufficiently elucidated. The objective of this study is to examine the expression of SLC27 in glioblastoma tumors and to investigate whether patient characteristics are linked to the expression of these genes. Furthermore, the study aims to explore the correlation between the expression of SLC27 and the essential enzymes responsible for fatty acid synthesis in glioblastoma tumors.
3. Results
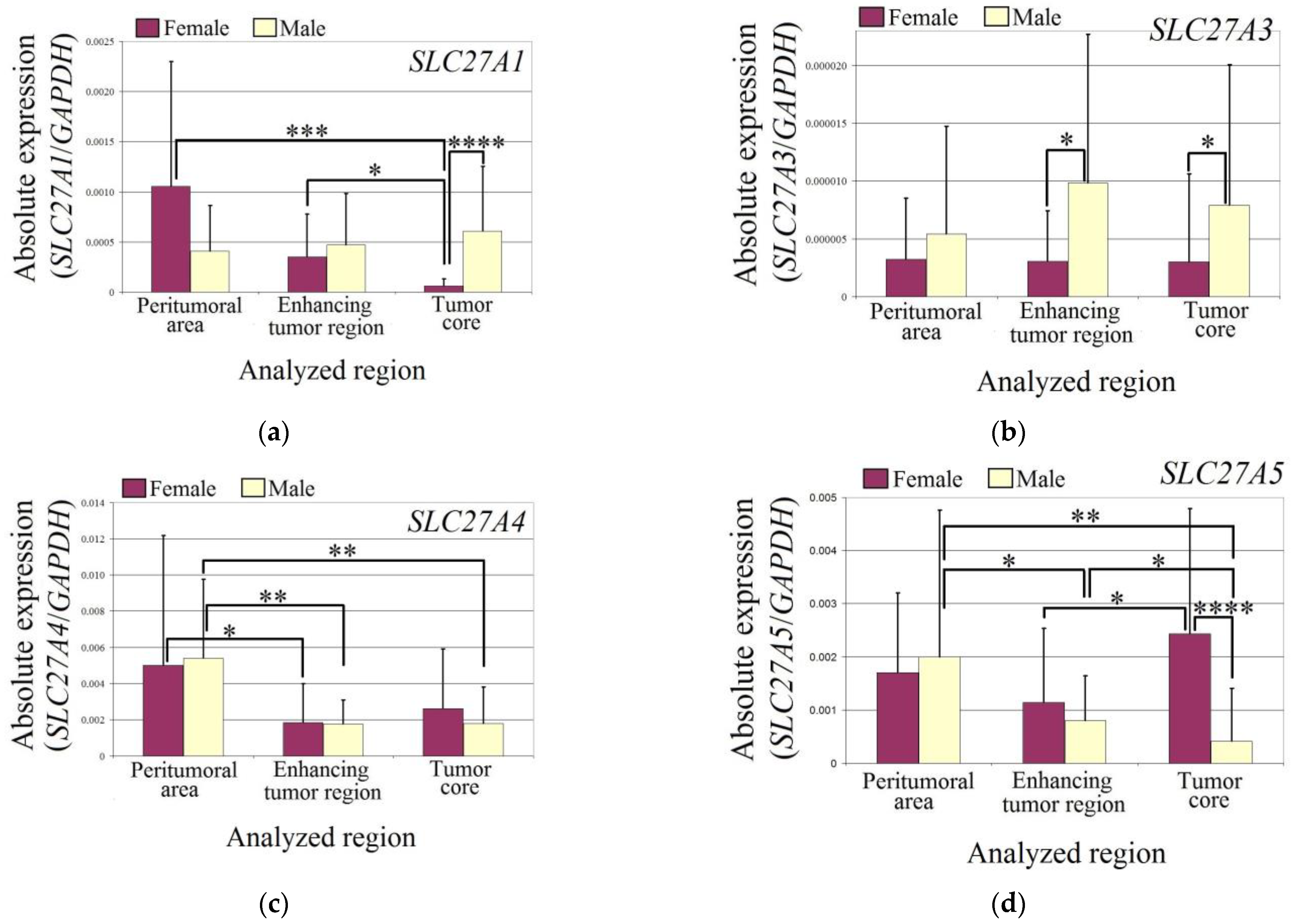
3.1. SLC27A4 and SLC27A6 Expression in the Glioblastoma Tumor Was Lower Than in the Peritumoral Area
Changes in the expression of individual SLC27 genes may contribute to alterations in the metabolism of fatty acids in glioblastoma tumors. Therefore, in the first stage of the study, the expression of
SLC27A genes was examined in two regions of the glioblastoma: the enhancing tumor region and the tumor core. The expression of selected genes was also analyzed in the peritumoral area, which is commonly considered as a control to which the results of the glioblastoma tumor analysis can be compared [
50].
The expression levels of
SLC27A1,
SLC27A3, and
SLC27A5 did not differ between the two analyzed regions of the glioblastoma tumor and the peritumoral area in all patients (
Figure 2). However, in the enhancing tumor region and the tumor core, the expression of
SLC27A4 was significantly lower compared to the peritumoral area (
p = 0.0007 and
p = 0.004, respectively). Similarly, the expression of
SLC27A6 was also significantly lower in the enhancing tumor region and the tumor core compared to the peritumoral area (
p = 0.014 and
p = 0.0019, respectively).
3.2. When Comparing Tumors between Sexes, the Expression of Different SLC27As Differed in Glioblastoma Tumors
Lipid metabolism is dependent on sex, particularly on sex hormones. Studies on breast cancer have shown that estrogen receptor-β (ER-β) increases the expression of
SLC27A1 [
52]. Estrogen receptors also influence the expression of long-chain acyl-CoA synthetases, other enzymes responsible for the activation of fatty acids. Specifically, estrogen receptor-α (ER-α) decreases the expression of acyl-CoA synthetase long-chain family member 4 (
ACSL4) [
53]. Due to the possibility of differences in the expression of the investigated SLC27 genes based on the sex of the patients, the expression results of SLC27A representatives were analyzed by sex and compared to each other.
The expression of SLC27A1 was reduced in the tumor core of female glioblastoma patients compared to the enhancing tumor region (p = 0.03) and peritumoral area (p = 0.0002). Additionally, the expression of SLC27A1 in the tumor core of female glioblastoma patients was lower than in male patients (p = 0.000003).
In the studied regions of glioblastoma tumors, the expression of SLC27A3 did not differ from the peritumoral area in males and females. However, in the enhancing tumor region (p = 0.04) and tumor core (p = 0.017), the expression of SLC27A3 was lower in female patients than in male patients.
The expression of SLC27A4 was lower in both males and females in the enhancing tumor region (in females p = 0.019; in males p = 0.004) and tumor core (in males p = 0.004) compared to the peritumoral area.
In males, the expression of SLC27A5 was lower in the glioblastoma tumor (in the enhancing tumor region p = 0.05 and in the tumor core p = 0.01) compared to the peritumoral area. However, the expression of SLC27A5 in the tumor core was lower than in the enhancing tumor region (p = 0.04). In females, the expression of SLC27A5 was higher in the tumor core compared to the enhancing tumor region (p = 0.019). Additionally, the expression of SLC27A5 in the tumor core was higher in females than in males (p = 0.00003).
The expression of
SLC27A6 was reduced in both sexes (
Figure 3). In female patients, the expression of
SLC27A6 in the enhancing tumor region was lower than in the peritumoral area (
p = 0.04). However, there was no difference in the expression of
SLC27A6 between the tumor core and peritumoral area in female patients (
p > 0.05). In male patients, the expression of
SLC27A6 in the tumor core was lower than in the peritumoral area (
p = 0.0009). Additionally, the expression of
SLC27A6 in the tumor core was lower than in the enhancing tumor region (
p = 0.006).
3.3. The Expression of SLC27A1 and SLC27A3 Negatively Correlated with the Expression of SLC27A4, SLC27A5, and SLC27A6
SLC27 representatives may have similar functions [
13]. They may be responsible for the channeling of fatty acids for the production of specific lipids [
8]. Additionally, the action of individual SLC27 representatives may exclude each other if they induce the channeling of fatty acids to different metabolic pathways. Therefore, to demonstrate the correlations between individual SLC27 representatives, the correlation between the expression levels of the investigated genes was analyzed.
The expression of SLC27A1 and SLC27A3 negatively correlated with the expression of SLC27A4, SLC27A5, and SLC27A6. The expression correlation between different regions of the tumor was rare. Only two SLC27 genes, SLC27A3 and SLC27A4, showed a correlation between their expression in the enhancing tumor region and the tumor core of the glioblastoma. Specifically, a negative correlation was found between the peritumoral area and the tumor core for SLC27A1, suggesting differences in fatty acid metabolism between these two regions.
In general, the expression levels of SLC27 genes were highly correlated within the same group. There was a negative correlation between each SLC27 gene in the first group and the second group. SLC27A1 and SLC27A3 belong to the first group, while SLC27A4, SLC27A5, and SLC27A6 belong to the second.
Positive correlations were observed between SLC27A4 and SLC27A5, SLC27A4 and SLC27A6, and SLC27A5 and SLC27A6 in all three tumor regions. In addition, a positive correlation was found between the expression of SLC27A1 and SLC27A3 in the glioblastoma tumor and between the expression of SLC27A1 and SLC27A4 in the tumor core. Negative correlations were observed between SLC27A3 and SLC27A4 in the enhancing tumor region and between SLC27A1 and SLC27A5 in the tumor core.
In rare cases, there was a correlation between the expression of different
SLC27 genes in different tumor locations (
Table 2). For example, a positive correlation was found between
SLC27A3 in the enhancing tumor region and
SLC27A1 in the tumor core, and between
SLC27A4 in the peritumoral area and
SLC27A6 in the tumor core. A negative correlation was found between
SLC27A1 in the peritumoral area and
SLC27A6 in the tumor core, between
SLC27A3 in the enhancing tumor region and
SLC27A5 and
SLC27A6 in the tumor core, and between
SLC27A3 in the tumor core and
SLC27A5 in the peritumoral area.
In women, as in all patients, there was a correlation between the expression of
SLC27A4 with
SLC27A5,
SLC27A4 with
SLC27A6, and
SLC27A5 with
SLC27A6 (
Table 3). However, no significant correlation was found between the expression of individual
SLC27 genes across different regions of the glioblastoma tumor. Women also showed a negative correlation in the expression of
SLC27A3 in the peritumoral area with the expression of
SLC27A4,
SLC27A5, and
SLC27A6 in the enhancing tumor region. Women also showed a negative correlation between the expression of
SLC27A1 in the peritumoral area and the expression of
SLC27A3 in the tumor core.
In men, SLC27 expression showed correlations similar to the analysis of all patients. There was a correlation of SLC27A3 expression between the enhancing tumor region and tumor core. Additionally, there was a correlation of SLC27A6 between the peritumoral area and the tumor core.
Similarly to the analysis of all patients, male samples showed a positive correlation of
SLC27A1 expression with
SLC27A3,
SLC27A4 with
SLC27A5,
SLC27A4 with
SLC27A6, and
SLC27A5 with
SLC27A6 in the studied regions of the glioblastoma tumor (
Table 4). Additionally, there was a positive correlation between
SLC27A5 expression in the tumor core and
SLC27A6 expression in the peritumoral area and between
SLC27A5 expression in the peritumoral area and
SLC27A6 expression in the tumor core. There was also a negative correlation between
SLC27A3 expression in the enhancing tumor region and
SLC27A5 in the peritumoral area.
3.4. The Expression of SLC27 in the Peritumoral Area Correlated with BMI
The process of carcinogenesis is extremely complex, with some models describing it as a Darwinian process in which the microenvironment plays a significant role [
54]. There are various factors that contribute to carcinogenesis, which can be divided into environmental (such as pollution) and patient-related factors. Examples of the latter group include age [
55], patient height [
56,
57], obesity [
58], and smoking [
59]. These characteristics influence the development of tumors. To investigate the possible correlations between SLC27 and carcinogenesis related to these characteristics, we analyzed the correlation between the expression of the investigated SLC27 genes and patient characteristics such as age, height, weight, BMI, and the number of cigarette packs smoked during their lifetime.
A correlation analysis was conducted on the expression of the investigated SLC27 representatives with patient characteristics such as age, height, weight, BMI, and the number of cigarette packs smoked in their lifetime. Across all patients, a correlation was found between the expression levels of the investigated SLC27 and BMI in the peritumoral area. For SLC27A1, a positive correlation was observed, while for SLC27A4, SLC27A5, and SLC27A6, a negative correlation was found. Additionally, a negative correlation was found between SLC27A5 expression in the tumor core and BMI. Furthermore, a negative correlation was found between the expression of SLC27A1 in the peritumoral area and patient age. Additionally, a positive correlation was observed between the expression of SLC27A1 in the tumor core and patient weight.
In female patients with glioblastoma, a positive correlation was found between the expression of SLC27A4, SLC27A5, and SLC27A6 and the number of cigarette packs smoked in their lifetime. SLC27A4 and SLC27A6 also showed a positive correlation in the peritumoral area. Similarly to the analysis of all patients, a negative correlation was observed between the expression of SLC27A4, SLC27A5, and SLC27A6 and BMI in the peritumoral area. Furthermore, a negative correlation was found between the expression of SLC27A1 in the enhancing tumor region and a positive correlation between SLC27A1 in the enhancing tumor region and patient weight. A negative correlation was also found between the expression of SLC27A5 and patient weight in the tumor core. Additionally, a negative correlation was observed between the expression of SLC27A1, SLC27A3, SLC27A5, and SLC27A6 in the peritumoral area and patient age. A correlation was also found between the expression of SLC27A and patient age in glioblastoma. Specifically, a positive correlation was found between SLC27A1 and SLC27A4 with age in the tumor core, while a negative correlation was found with SLC27A5. In the enhancing tumor region, a positive correlation was found between age and the expression of SLC27A6 in female patients.
In male patients, a correlation was found between the expression of
SLC27 in glioblastoma and BMI (
Table 5). A positive correlation was observed between the expression of
SLC27A1 and
SLC27A3 in the enhancing tumor region, while a negative correlation was found with the expression of
SLC27A4,
SLC27A5, and
SLC27A6. In the tumor core, a negative correlation was found between the expression of
SLC27A5 and BMI. Similar to the analyses of all patients, a negative correlation was observed between the expression of
SLC27A5 and BMI in the peritumoral area. Furthermore, a correlation was found between the expression of the investigated
SLC27 and patient age. Specifically, a positive correlation was observed between
SLC27A3 and age in the enhancing tumor region, and a negative correlation was observed with
SLC27A5.
3.5. SLC27 Expression Correlated with Major Fatty Acid Synthesis Enzymes
We conducted an analysis to investigate the correlation between the expression of
SLC27 members and the expression of key fatty acid synthesis enzymes, including
ELOVL1,
ELOVL3,
ELOVL6,
SCD, and
FADS2. Expression data for
ELOVL1,
ELOVL3,
ELOVL6,
SCD, and
FADS2 were obtained from raw data published in our previous papers, where we examined the expression of elongases [
47] and desaturases [
46] in glioblastomas.
ELOVL6 is involved in the synthesis of stearic acid C18:0 from palmitic acid C16:0 [
6], which is responsible for
the de novo synthesis of the majority of SFA and monounsaturated fatty acid (MUFA).
ELOVL1 and
ELOVL3 are responsible for the further elongation of stearic acid C18:0 up to 26 carbons in length [
6]. According to GEPIA, these two elongases play a significant role in the tumorigenesis of glioblastomas. Elevated expression of
ELOVL1 and
ELOVL3 in this tumor type is associated with a poorer prognosis [
43].
SCD is a desaturase responsible for the production of MUFA from SFA, while
FADS2 is the first enzyme in the biosynthesis pathway of PUFA, regulating the entire pathway [
6].
For all patients, we found a positive correlation between the expression of SLC27A4, SLC27A5, and SLC27A6, and the expression of SCD and FADS2 in the peritumoral area. In all studied regions, SLC27A1 expression was positively correlated with ELOVL6 expression. Furthermore, SLC27A3 expression was positively correlated with ELOVL6 expression in the enhancing tumor region and tumor core. With regard to ELOVL1 and ELOVL3, no significant correlation with the analyzed SLC27 was found in all patients.
In women, there was a positive correlation between ELOVL1 expression and SLC27A, SLC27A5, and SLC27A6 expression in the tumor core. We observed a positive correlation between the expression of SLC27A4, SLC27A5, and SLC27A6 and the expression of SCD and FADS2 in the peritumoral area. Additionally, a positive correlation between the expression of SLC27A1 and SCD, and FADS2 was observed in the peritumoral area. Finally, a positive correlation between the expression of SLC27A1 and ELOVL6 was found in the tumor core, but no correlation was observed in the enhancing tumor region.
In men, a positive correlation was observed between the expression of
ELOVL1 and
SLC27A1, and
SLC27A3 in the tumor core (
Table 6). Furthermore, a negative correlation was found between the expression of
ELOVL1, and
SLC27A5 and
SLC27A6, which is opposite to the correlation observed in women. In men, we found a positive correlation between the expression of
SLC27A4 and
SLC27A6 and the expression of
SCD in the peritumoral area. Additionally, a positive correlation between the expression of
SLC27A1 and
ELOVL6 was observed in all three studied regions.
4. Discussion
4.1. SLC27 Expression in Glioblastoma
In this study, we showed that
SLC27A4 and
SLC27A6 expression was lower in the glioblastoma compared to the peritumoral area, while the expression of the other examined
SLC27 genes did not differ between the glioblastoma and peritumoral area. However, it is difficult to compare our results with other scientific articles because analyses of
SLC27 expression are not available in the PubMed database (
https://pubmed.ncbi.nlm.nih.gov, accessed 25 February 2023). There are only two available studies that examine
SLC27A3 [
32,
60]. Therefore, to compare our findings with other research groups, we used the Gene Expression Profiling Interactive Analysis (GEPIA) database (
http://gepia.cancer-pku.cn, accessed 25 February 2022) [
43] and the transcriptomic analysis of different grades of glioma conducted by Seifert et al. [
61]. GEPIA contains analyses of gene expression in over 30 types of cancer, including glioblastoma [
43]. These are expression analyses taken from The Cancer Genome Atlas (TCGA) [
62] and data obtained from healthy tissues from the Genotype-Tissue Expression (GTEx) database [
63,
64]. Transcriptomic analysis of different grades of glioma conducted by Seifert et al. was analyzed using the REpository of Molecular BRAin Neoplasia DaTa (Rembrandt) [
65].
In this study, we found that
SLC27A1 and
SLC27A5 expression did not differ between the glioblastoma and peritumoral area. These results are consistent with GEPIA [
43] and Seifert et al. [
61]. We also found that there was a lower expression of
SLC27A4 in the glioblastoma compared to the peritumoral area, and these results were consistent with Seifert et al. [
61]. However, GEPIA reports that
SLC27A4 expression does not differ between glioblastomas and healthy brain tissue [
43]. We also found lower expression of
SLC27A6 in the glioblastoma compared to the peritumoral area. This is in contrast to GEPIA [
43] and Seifert et al. [
61], where
SLC27A6 expression did not differ between the glioblastoma and healthy brain tissue.
In this study, we did not obtain results for
SLC27A2, an enzyme characterized by very low expression in the brain [
21,
29,
30]. GEPIA analyses also show that the level of
SLC27A2 expression is about 200 times lower in glioblastoma than among other
SLC27 genes [
43]. As a result, it can be inferred that the level of
SLC27A2 expression is below the detection threshold of the method used in this study. Furthermore, the low expression of
SLC27A2 in glioblastoma tumors suggests that it does not play a significant role in the tumorigenesis processes in glioblastoma. This is supported by the lack of association between the level of
SLC27A2 expression in glioblastoma tumors and patient prognosis [
43].
4.2. Differences in SLC27 Expression in Glioblastoma Tumors between Sexes
In glioblastoma tumors, specifically in the tumor core, women exhibit lower expression of SLC27A1 and SLC27A3, but higher expression of SLC27A5 than men. This suggests differences in fatty acid metabolism between sexes. However, there is a lack of research on sex differences in SLC27 expression in the brain or even in other organs, and therefore, we cannot refer to the existing literature.
SLC27 shows similar activity to specific fatty acids [
13]. However, SLC27A3 and SLC27A5 demonstrate weaker fatty acid activation properties compared to other SLC27 [
13]. Moreover, SLC27A3, SLC27A5, and SLC27A6 show weak or no uptake properties of fatty acids [
13]. SLC27A1 participates in the transport of fatty acids across the BBB [
22], indicating that in women, the uptake of fatty acids from the bloodstream by tumor core cells in glioblastoma may be less intense than in men.
SLC27A3 participates in ceramide synthesis, as demonstrated in experiments on U-87 MG cells [
32]. This suggests that the production of ceramides in glioblastoma tumors may be less intense in women. Ceramides play a critical role in the tumorigenesis processes in glioblastoma [
32,
66,
67] and the higher expression of SLC27A3 is associated with a worse prognosis [
32]. Therefore, this may explain the better prognosis for women with glioblastoma [
68].
4.3. Correlations between the Expression of Various SLC27
Analyses of the correlation between different
SLC27 in various studied regions indicate certain dependencies in fatty acid metabolism. A positive correlation of
SCL27A4 expression was found between the tumor core and enhancing tumor region but not with the peritumoral area. A negative correlation of
SLC27A1 expression was observed between the tumor core and the peritumoral area.
SCL27A1 and
SCL27A4 are important in fatty acid uptake through BBB [
22]. This suggests that fatty acid uptake through BBB is similar in the two studied regions of the glioblastoma tumor, but differs significantly from the peritumoral area. Among the other
SLC27, a positive correlation of
SLC27A3 expression was found between all studied regions of glioblastoma. SLC27A3 is responsible for the production of ceramides, as demonstrated in experiments on U-87 MG cells [
32]. This correlation indicates that ceramide synthesis may be similar between tumors and the rest of the brain. However, the lack of correlation of other
SLC27 suggests that lipid metabolism significantly differs between the tumor and peritumoral area.
In our study, we found a negative correlation between the expression of
SLC27A3 and the expression of
SLC27A4 and
SLC27A5, and a positive correlation between the expression of
SLC27A3 and
SLC27A1. SLC27A3 is a protein that does not participate in fatty acid uptake [
13]. It is responsible for channeling fatty acids to produce ceramides, as demonstrated in experiments on U-87 MG cells [
32]. SLC27A1 is responsible for the synthesis of triglycerides [
16,
17,
18], ceramides [
9], and β-oxidation [
18,
19], as demonstrated in other models. Meanwhile, SLC27A4 is responsible for the production of triglycerides [
17,
36], cholesterol esters [
36], and ceramide [
9]. SCL27A1 and SCL27A4 are important in fatty acid uptake through BBB [
22]. SLC27A5 may be responsible for the production of triglycerides, glycerophospholipids, and cholesteryl esters [
39]. The observed correlations suggest that ceramide biosynthesis may occur in glioblastoma, which precludes the use of fatty acids for the production of other lipids. Ceramide production is also positively correlated with the expression and uptake of fatty acids by SLC27A1. Moreover, a positive correlation for expression was found between
SLC27A4,
SLC27A5, and
SLC27A6, while a negative correlation was observed between
SLC27A1 and
SLC27A5 expression. This suggests that the production of triglycerides, glycerophospholipids, and cholesteryl esters may depend on the uptake of fatty acids through BBB, depending on SLC27A4 but not SLC27A1.
In the tumor core, but not in the enhancing tumor region, there was a positive correlation between the expression of
SLC27A1 and
SLC27A4, both of which are involved in fatty acid uptake through the BBB [
22]. This suggests that fatty acid uptake in the tumor core is mediated by these two proteins, whose expression levels are positively correlated. In contrast, there was no correlation between the expression of
SLC27A1 and
SLC27A4 in the enhancing tumor region, suggesting that fatty acid uptake in this region may depend on one of these proteins.
4.4. Correlation of SLC27 Expression with Patient Characteristics
The correlation of
SLC27 gene expression with patient characteristics was analyzed, including age, BMI, height, and smoking history. Negative correlations were observed between the expression of
SLC27A4,
SLC27A5, and
SLC27A6 and BMI in the peritumoral area of the patients, with
SLC27A5 and
SLC27A6 showing this correlation in both genders. Additionally, the expression of
SLC27A1 was positively correlated with BMI in all patients. SLC27A1 and SLC27A4 are involved in the uptake of fatty acids by the BBB [
22], with SLC27A1 specifically participating in the transport of DHA C22:6n-3 [
23,
24]. This suggests that in obese patients, there may be a lower expression of
SLC27A4 and higher expression of
SLC27A1 in the brain, indicating greater uptake of DHA C22:6n-3 by the brain. The results also indicate that lipid metabolism in the brain may be altered in obese individuals, but further research is needed to fully understand the significance of SLC27 in obesity. Currently, no experimental studies are available on this topic.
In women, there was a negative correlation between the expression of
SLC27A1 and weight and BMI in the enhancing tumor region and a positive correlation with weight in the tumor core. This suggests differences in the uptake and metabolism of fatty acids between these two regions of glioblastoma tumors in lean and obese women. In the peritumoral area, there was also a negative correlation of
SLC27 with weight in women. According to one available study [
69], obese women have lower expression of
SLC27A1 in muscle tissue compared to lean women, which may explain the negative correlation between
SLC27A1 expression and BMI and weight in the enhancing tumor region of glioblastoma tumors and the peritumoral area in women. However, the cited study [
69] shows no difference in
SLC27A1 expression between obese and lean men. In this study, a positive correlation was found between BMI and
SLC27A1 expression in the enhancing tumor region of glioblastoma tumors in men, which is inconsistent with the cited study that examined expression in muscle tissue. In men with glioblastoma tumors, there was also a negative correlation between the expression of
SLC27A4,
SLC27A5,
SLC27A6, and a positive correlation between
SLC27A1 and
SLC27A3 expression with BMI. This indicates significant differences in fatty acid metabolism between obese and lean men, with greater uptake of fatty acids by the BBB involving SLC27A1 and less involving SLC27A4 in obese men [
22]. This may result in increased uptake of DHA C22:6n-3 by glioblastoma tumors in obese men [
23,
24]. Obese men may also have higher production of ceramides [
32] and less intensive metabolism of other types of lipids [
39] compared to lean men.
In women, there was a negative correlation between the expression of the tested SLC27 genes and age in the peritumoral area, whereas no such relationship was observed in men. These results may indicate changes in fatty acid metabolism in the healthy brain that occur with age, especially in women. However, there is a lack of experimental studies in this direction. In the tumor core of women, there was a negative correlation between the expression of SLC27A5 and a positive correlation between the expression of SLC27A4 and SLC27A1 with age. Additionally, in the enhancing tumor region, there was a positive correlation between the expression of SLC27A6 and age. In men, there was a negative correlation between the expression of SLC27A5 and a positive correlation between the expression of SLC27A3 with age in the enhancing tumor region. These findings also indicate that the patient’s age and sex significantly affect fatty acid metabolism in glioblastoma tumors.
In women, there was a positive correlation between the expression of SLC27A4, SLC27A5, and SLC27A6 with cigarette smoking in all investigated regions of the glioblastoma tumor and in the peritumoral area, whereas no such relationship was observed in men. These data suggest that cigarette smoking in women with glioblastoma may affect carcinogenesis and, consequently, may influence fatty acid metabolism in the tumor. However, there is a lack of available studies on the influence of cigarette smoking on the expression of SLC27 enzymes in tumors, including lung cancer. Therefore, further research is needed to investigate the association between cigarette smoking and fatty acid metabolism in tumors in women.
4.5. Correlation with Enzymes Involved in Fatty Acid Synthesis
We have demonstrated a positive correlation between the expression of SLC27A1 and SLC27A3 and the expression of ELOVL6 in both the glioblastoma tumor and the peritumoral area. Furthermore, we have identified a positive correlation between the expression of SLC27A4, SLC27A5, and SLC27A6 and the expression of SCD and FADS2 in the peritumoral area. These findings shed some light on the metabolic pathways of fatty acids in glioblastoma tumors.
ELOVL6 participates in the synthesis of stearic acid C18:0 from palmitic acid C16:0 [
6], while SCD and FADS2 are involved in the production of MUFAs and PUFAs, respectively. SLC27 proteins transport fatty acids across the cell membrane and activate the transported fatty acids [
13,
28,
35]. These proteins exhibit activity towards various fatty acids from different groups [
13]. Specifically, they activate SFAs (palmitic acid C16:0, stearic acid C18:0, lignoceric acid C24:0, and cerotic acid C26:0 [
12,
13,
37]), MUFAs (oleic acid C18:1n-9) [
13], and PUFAs (arachidonic acid C20:4n-6) [
13]. However, the possibility of transporting and activating other fatty acids (such as linoleic acid C18:2n-6) has not been studied. The high activity of SLC27 towards various fatty acids suggests that other fatty acids may also be transported and activated by these proteins.
SLC27A1 and SLC27A4 are the most important SLC27 proteins involved in the transport of fatty acids across the BBB [
22]. Our data suggest a correlation between the transport of palmitic acid C16:0 by SLC27A1 across the BBB and the elongation of this fatty acid. This provides insights into the potential effects of drugs targeting SLC27A1. SLC27A3 exhibits similar properties, although it does not account for fatty acid uptake [
13]. It may be crucial for intracellular fatty acid metabolism, particularly in the synthesis of ceramides, as evidenced by experiments on U-87 MG cells [
32]. Moreover, fatty acids transported by SLC27A4 across the BBB undergo different modifications in the peritumoral area. SFAs (palmitic acid C16:0 and stearic acid C18:0) are desaturated, i.e., transformed into MUFAs. In contrast, 18-carbon PUFAs are activated and transformed into longer PUFAs, such as arachidonic acid C20:4n-6, eicosapentaenoic acid C20:5n-3, and docosahexaenoic acid C22:6n-3. Interestingly, the observed correlations indicate that SLC27A5 and SLC27A6 exhibit similar activities to SLC27A4, although a correlation with
FADS2 was only observed in the peritumoral area. This suggests that such a correlation does not occur in the glioblastoma tumor itself.
In women, a positive correlation was observed between the expression of ELOVL1 and SLC27A4, SLC27A5, and SLC27A6 in the tumor core. This finding suggests that very-long-chain SFAs may be taken up from the blood by SLC27A4 and subsequently elongated by ELOVL1 to be incorporated into triglycerides, glycerophospholipids, and cholesteryl esters in the tumor core. In contrast, in men, a positive correlation was found between the expression of ELOVL1 and SLC27A1 and SLC27A3, and a negative correlation with SLC27A5 and SLC27A6, which is opposite to the correlation observed in women. This indicates that the metabolism of very-long-chain SFAs differs between sexes. In men, very-long-chain SFAs may be elongated and incorporated into ceramides after uptake by SLC27A1.
4.6. Clinical Significance of the Obtained Results
Analyses on the GEPIA portal show that the expression level of any
SLC27 gene is not significantly associated with prognosis for patients with glioblastoma [
43]. However, comparing the highest and lowest quartiles of
SLC27 expression, a trend towards worse outcomes with higher expression of
SLC27A4 (
p = 0.062) and
SLC27A3 (
p = 0.083) was observed for glioblastoma patients [
43]. This suggests that these two SLC27 representatives may have important clinical significance as a therapeutic target.
According to Kolar et al., analyzing The Chinese Glioma Genome Atlas, higher expression of
SLC27A3 in glioblastoma tumors is associated with worse outcomes [
32]. This suggests that this protein plays a significant role in the tumorigenic processes in this cancer type. This enzyme activates fatty acids and channels them towards ceramide production [
32]. SLC27A3 is essential in the stemness and self-renewal of glioblastoma stem cells [
33], as well as in the proliferation and anchorage-independent growth of glioma cells [
60]. Currently, no published studies on the significance of SLC27A4 in glioblastoma tumorigenesis are available on the PubMed browser (
https://pubmed.ncbi.nlm.nih.gov/, accessed 25 February 2022). However, SLC27A4 is known to play a crucial role in brain physiology by participating in the transport of fatty acids across the BBB, together with SLC27A1 [
22]. This suggests that SLC27A4 may also be important in the uptake of fatty acids from the bloodstream by glioblastoma cells. Nonetheless, SLC27A4 may also be significant in the tumorigenesis of other cancers. Higher expression of
SLC27A4 in breast tumors is associated with worse prognosis, particularly with distant metastasis-free survival [
70]. In breast cancer cells, SLC27A4 plays a role in proliferation and epithelial-to-mesenchymal transition (EMT) [
70]. Similarly, studies on hepatocellular carcinoma (HCC) cells have demonstrated that SLC27A4 is crucial for the proliferation and invasion of these cells [
71]. In lung cancer cells, SLC27A4 is responsible for chemoresistance [
72].
The results of this study have shown that the expression of
SLC27A3 was higher in the glioblastoma tumors of men than of women. Additionally, the expression of
SLC27A3 in the enhancing tumor region of men was positively correlated with the age and BMI of the patient. Furthermore, the expression of
SLC27A3 in men was positively correlated with the expression of
ELOVL1 in this region of the glioblastoma tumor.
ELOVL1 is an enzyme responsible for the elongation of SFA [
6], and the expression of this enzyme is also associated with poor patient outcomes in glioblastoma [
43]. These observations may be useful for the development of personalized therapies, especially for obese men who are much older than the average glioblastoma patient. Such therapies should target the enzymes responsible for the synthesis of ceramides containing very-long-chain SFAs (either the studied SLC27A3 or ELOVL1) or enzymes directly responsible for ceramide synthesis.
Another enzyme described in this study that may affect outcomes in glioblastoma patients is SLC27A4. We have shown that the expression of SLC27A4 was the same in both sexes. It was negatively correlated with BMI and height in men, and positively correlated with age and the intensity of cigarette smoking in women. In women, but not in men, the expression of SLC27A4 was positively correlated with the expression of ELOVL1, indicating a different function of this enzyme between sexes. Drugs targeting SLC27A4 could be helpful in treating glioblastoma in low-weight men who are lean or thin. In female glioblastoma patients, such drugs could have a therapeutic effect in old age, and in cases when the glioblastoma developed as a result of intense cigarette smoking.
4.7. Limitations
The objective of this study was to investigate the expression of SLC27 in glioblastoma tumors and to explore the correlation between the expression of the investigated proteins and the key enzymes involved in fatty acid biosynthesis, as well as patient characteristics such as sex, age, BMI, and smoking. However, there are several limitations to the study. The expression levels of the investigated genes in glioblastoma tumors were compared to those in the peritumoral area rather than healthy brain tissue due to ethical issues with obtaining samples of healthy brain tissue. While epilepsy surgery brain samples are sometimes used as a control, they may be more similar to brain tumors than healthy brain tissue [
50]. Therefore, using the peritumoral area as a control seems to be the most reasonable option. Another limitation is the small sample size of patients, all of whom come from a single population. Nevertheless, the obtained results are consistent with the available literature data, suggesting that the conclusions drawn from the study may provide valuable data for scientific research. Additionally, the study did not investigate the correlation between the investigated genes and the expression of lipid biosynthesis enzymes such as sphingolipids, glycerophospholipids, and triacylglycerols. As a result, the significance of SLC27 in the entire lipid metabolism of glioblastoma tumors was not fully demonstrated, indicating a potential direction for further research into lipid metabolism in glioblastomas.
5. Conclusions
Several conclusions can be drawn from the results of this study. The expression of SLC27, responsible for transporting fatty acids through the BBB, is reduced in glioblastoma tumors. This suggests a lower uptake of fatty acids by the tumor compared to non-tumor brain tissue. Therefore, it can be inferred that glioblastoma tumors exhibit a more intense biosynthesis of fatty acids than their uptake from the bloodstream. This metabolic difference might set them apart from healthy brain tissue. Thus, a therapeutic approach aimed at de novo synthesis of fatty acids may be more effective than targeting the transport of fatty acids.
Furthermore, SLC27 is associated with the de novo synthesis of fatty acids. Specifically, SLC27A1 and SLC27A3 are associated with stearic acid synthesis, and SLC27A4, SLC27A5, and SLCA6 with very-long-chain-fatty-acid synthesis. There are differences in the expression of these genes between genders, which showed a positive correlation in women and negative correlation in men.
Lastly, the transport and activation of fatty acids in glioblastoma tumors may depend on the intensity of cigarette smoking in women and BMI in men. This factor may influence the development of a personalized therapy approach for each patient.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}