




Effects of Maternal Separation and Subsequent Stress on Behaviors and Brain Monoamines in Rats


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.3. Behavioral Tests
2.4. Automated Open Field
2.5. Classical Open Field
2.6. Assessment of the Anxiety–Phobic Level
2.7. Elevated plus Maze
2.8. Social Interaction Test
2.9. Three-Chamber Social Test
2.10. Additional Repeated Stress Exposure of Adult Rats
2.11. Measurement of the Level of Monoamines and Their Metabolites in Brain Structures
2.12. Statistical Processing of the Results
3. Results
3.1. Body Weight
3.2. Locomotor and Exploratory Activity
3.3. Anxiety-like Behavior
3.4. Social Behavior
3.5. Monoamines
3.5.1. Norepinephrine
3.5.2. Dopamine
3.5.3. 5-Hydroxytryptamine
4. Discussion
4.1. Body Weight
4.2. Locomotor Activity, Exploratory Activity, and Anxiety-like Behavior
4.3. Social Behavior
4.4. Monoamines
4.5. Additional repeated Stress in Adult Rats
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Singh, J.; Prakash, J.; Yadav, P.; Bharti, A.; Chatterjee, K. Complex psychological trauma. Ind. Psychiatry J. 2021, 30 (Suppl. 1), S305–S307. [Google Scholar] [CrossRef]
- Jowett, S.; Karatzias, T.; Shevlin, M.; Hyland, P. Psychological trauma at different developmental stages and ICD-11 CPTSD: The role of dissociation. J. Trauma Dissociation 2022, 23, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.; Palmier-Claus, J.; Branitsky, A.; Mansell, W.; Warwick, H.; Varese, F. Childhood adversity and borderline personality disorder: A meta-analysis. Acta Psychiatr. Scand. 2020, 141, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.; Nasa, A.; Cullinane, D.; Raajakesary, K.; Gazzaz, A.; Sooknarine, V.; Haines, M.; Roman, E.; Kelly, L.; O’Neill, A.; et al. Childhood Trauma, the HPA Axis and Psychiatric Illnesses: A Targeted Literature Synthesis. Front. Psychiatry 2022, 13, 748372. [Google Scholar] [CrossRef] [PubMed]
- Cruz, D.; Lichten, M.; Berg, K.; George, P. Developmental trauma: Conceptual framework, associated risks and comorbidities, and evaluation and treatment. Front. Psychiatry 2022, 13, 800687. [Google Scholar] [CrossRef]
- Hyland, P.; Karatzias, T.; Shevlin, M.; Cloitre, M. Examining the discriminant validity of complex posttraumatic stress disorder and borderline personality disorder symptoms: Results from a United Kingdom population sample. J. Trauma. Stress 2019, 32, 855–863. [Google Scholar] [CrossRef]
- Ehrenthal, J.C.; Levy, K.N.; Scott, L.N.; Granger, D.A. Attachment-related regulatory processes moderate the impact of adverse childhood experiences on stress reaction in borderline personality disorder. J. Pers. Disord. 2018, 32, 93–114. [Google Scholar] [CrossRef]
- Owczarek, M.; Karatzias, T.; McElroy, E.; Hyland, P.; Cloitre, M.; Kratzer, L.; Knefel, M.; Grandison, G.; Ho, G.W.K.; Morris, D.; et al. Borderline Personality Disorder (BPD) and Complex Posttraumatic Stress Disorder (CPTSD): A Network Analysis in a Highly Traumatized Clinical Sample. J. Pers. Disord. 2023, 37, 112–129. [Google Scholar] [CrossRef]
- Jowett, S.; Karatzias, T.; Shevlin, M.; Albert, I. Differentiating symptom profiles of ICD-11 PTSD complex PTSD, and borderline personality disorder: A latent class analysis in a multiply traumatized sample. Pers. Disord 2020, 11, 36–45. [Google Scholar] [CrossRef]
- Ford, J.D.; Courtois, C.A. Complex PTSD and borderline personality disorder. Bord. Pers. Disord. Emot. Dysregulation 2021, 8, 16. [Google Scholar] [CrossRef]
- Stahl, S.M. Stahl’s Essential Psychopharmacology: Neuroscientific Basis and Practical Applications; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Pinggal, E.; Dockree, P.M.; O’Connell, R.G.; Bellgrove, M.A.; Andrillon, T. Pharmacological Manipulations of Physiological Arousal and Sleep-Like Slow Waves Modulate Sustained Attention. J. Neurosci. 2022, 42, 8113–8124. [Google Scholar] [CrossRef] [PubMed]
- Silk, K.R. Borderline personality disorder: Overview of biological factors. Psychiatr. Clin. N. Am. 2000, 23, 61–76. [Google Scholar] [CrossRef]
- Rosell, D.R.; Slifstein, M.; Thompson, J.; Xu, X.; Perez-Rodriguez, M.M.; McClure, M.M.; Hazlett, E.A.; New, A.S.; Nabulsi, N.; Huang, Y.; et al. Serotonin transporter availability in physically aggressive personality disordered patients: Associations with trait and state aggression, and response to fluoxetine. Psychopharmacology 2023, 240, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Rylands, A.J.; Hinz, R.; Jones, M.; Holmes, S.E.; Feldmann, M.; Brown, G.; McMahon, A.W.; Talbot, P.S. Pre-and postsynaptic serotonergic differences in males with extreme levels of impulsive aggression without callous unemotional traits: A positron emission tomography study using (11)C-DASB and (11)C-MDL100907. Biol. Psychiatry 2012, 72, 1004–1011. [Google Scholar] [CrossRef]
- van de Giessen, E.; Rosell, D.R.; Thompson, J.L.; Xu, X.; Girgis, R.R.; Ehrlich, Y.; Slifstein, M.; Abi-Dargham, A.; Siever, L.J. Serotonin transporter availability in impulsive aggressive personality disordered patients: A PET study with [11C]DASB. J. Psychiatr. Res. 2014, 58, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, M.D.; Bridges, J.; Sinnerton, R.; Nakamura, A.; Underwood, J.F.G.; Slater, A.; Lee, M.R.D.; Clarke, L.; Lewis, C.; Roberts, N.P.; et al. Pharmacological therapy for post-traumatic stress disorder: A systematic review and meta-analysis of monotherapy, augmentation and head-to-head approaches. Eur. J. Psychotraumatology 2021, 12, 1802920. [Google Scholar] [CrossRef]
- Paret, C.; Niedtfeld, I.; Lotter, T.; Wunder, A.; Grimm, S.; Mennes, M.; Okell, T.; Beckmann, C.; Schmahl, C. Single-Dose Effects of Citalopram on Neural Responses to Affective Stimuli in Borderline Personality Disorder: A Randomized Clinical Trial. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 837–845. [Google Scholar] [CrossRef]
- Hendrickson, R.C.; Raskind, M.A. Noradrenergic dysregulation in the pathophysiology of PTSD. Exp. Neurol. 2016, 284, 181–195. [Google Scholar] [CrossRef]
- Buckholtz, J.W.; Treadway, M.T.; Cowan, R.L.; Woodward, N.D.; Li, R.; Ansari, M.S.; Baldwin, R.M.; Schwartzman, A.N.; Shelby, E.S.; Smith, C.E.; et al. Dopaminergic network differences in human impulsivity. Science 2010, 329, 532. [Google Scholar] [CrossRef]
- Ross, J.A.; Van Bockstaele, E.J. The role of catecholamines in modulating responses to stress: Sex-specific patterns, implications, and therapeutic potential for post-traumatic stress disorder and opiate withdrawal. Eur. J. Neurosci. 2020, 52, 2429–2465. [Google Scholar] [CrossRef]
- Del Casale, A.; Bonanni, L.; Bargagna, P.; Novelli, F.; Fiaschè, F.; Paolini, M.; Forcina, F.; Anibaldi, G.; Cortese, F.N.; Iannuccelli, A.; et al. Current Clinical Psychopharmacology in Borderline Personality Disorder. Curr. Neuropharmacol. 2021, 19, 1760–1779. [Google Scholar] [CrossRef]
- Wagner, G.; Krause-Utz, A.; de la Cruz, F.; Schumann, A.; Schmahl, C.; Bär, K.J. Resting-state functional connectivity of neurotransmitter producing sites in female patients with borderline personality disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 83, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, A.M.; Pineles, S.L. Neurotransmitter, Peptide, and Steroid Hormone Abnormalities in PTSD: Biological Endophenotypes Relevant to Treatment. Curr. Psychiatry Rep. 2018, 20, 52. [Google Scholar] [CrossRef] [PubMed]
- Gurvits, I.G.; Koenigsberg, H.W.; Siever, L.J. Neurotransmitter dysfunction in patients with borderline personality disorder. Psychiatr. Clin. N. Am. 2000, 23, 27–40. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual for Mental Disorders, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Al Jowf, G.I.; Ahmed, Z.T.; Reijnders, R.A.; de Nijs, L.; Eijssen, L.M.T. To Predict, Prevent, and Manage Post-Traumatic Stress Disorder (PTSD): A Review of Pathophysiology, Treatment, and Biomarkers. Int. J. Mol. Sci. 2023, 24, 5238. [Google Scholar] [CrossRef]
- Dunsmoor, J.E.; Cisler, J.M.; Fonzo, G.A.; Creech, S.K.; Nemeroff, C.B. Laboratory models of post-traumatic stress disorder: The elusive bridge to translation. Neuron 2022, 110, 1754–1776. [Google Scholar] [CrossRef]
- Neigh, G.N.; Ritschel, L.A.; Kilpela, L.S.; Harrell, C.S.; Bourke, C.H. Translational reciprocity: Bridging the gap between preclinical studies and clinical treatment of stress effects on the adolescent brain. Neuroscience 2013, 249, 139–153. [Google Scholar] [CrossRef]
- Cattane, N.; Vernon, A.C.; Borsini, A.; Scassellati, C.; Endres, D.; Capuron, L.; Tamouza, R.; Benros, M.E.; Leza, J.C.; Pariante, C.M.; et al. Preclinical animal models of mental illnesses to translate findings from the bench to the bedside: Molecular brain mechanisms and peripheral biomarkers associated to early life stress or immune challenges. European College of Neuropsychopharmacology (ECNP) ImmunoNeuroPsychiatry Thematic Working Group. Eur. Neuropsychopharmacol. 2022, 58, 55–79. [Google Scholar] [CrossRef] [PubMed]
- Nishi, M. Effects of Early-Life Stress on the Brain and Behaviors: Implications of Early Maternal Separation in Rodents. Int. J. Mol. Sci. 2020, 21, 7212. [Google Scholar] [CrossRef]
- Corniquel, M.B.; Koenigsberg, H.W.; Likhtik, E. Toward an animal model of borderline personality disorder. Psychopharmacology 2019, 236, 2485–2500. [Google Scholar] [CrossRef]
- Nia, A.B.; Eveleth, M.C.; Gabbay, J.M.; Hassan, Y.J.; Zhang, B.; Pere-Rodriguez, M.M. Past, present, and future of genetic research in borderline personality disorder. Curr. Opin. Psychol. 2018, 21, 60–68. [Google Scholar] [CrossRef]
- Teng, T.; Fan, L.; Yan, W.; Li, X.; Zhang, Y.; Xiang, Y.; Jiang, Y.; Yuan, K.; Yin, B.; Shi, L.; et al. A diathesis-stress rat model induced suicide-implicated endophenotypes and prefrontal cortex abnormalities in the PKA and GABA receptor signaling pathways. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 116, 110538. [Google Scholar] [CrossRef]
- Hackenberg, T.D.; Vanderhooft, L.; Huang, J.; Wagar, M.; Aexander, J.; Tan, L. Social preference in rats. J. Exp. Anal. Behav. 2021, 115, 634–649. [Google Scholar] [CrossRef]
- Schuweiler, D.R.; Rao, M.; Pribut, H.J.; Roesch, M.R. Rats delay gratification during a time-based diminishing returns task. J. Exp. Psychol. Anim. Learn. Cogn. 2021, 47, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Kestering-Ferreira, E.; Tractenberg, S.G.; Lumertz, F.S.; Orso, R.; Creutzberg, K.C.; Wearick-Silva, L.E.; Viola, T.W.; Grassi-Oliveira, R. Long-term Effects of Maternal Separation on Anxiety-Like Behavior and Neuroendocrine Parameters in Adult Balb/c Mice. Chronic Stress 2021, 5, 24705470211067181. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Cao, K.; Lin, H.; Cui, S.; Shen, C.; Wen, W.; Mo, H.; Dong, Z.; Bai, S.; Yang, L.; et al. Early-Life Stress Induces Depression-Like Behavior and Synaptic-Plasticity Changes in a Maternal Separation Rat Model: Gender Difference and Metabolomics Study. Front. Pharmacol. 2020, 11, 102. [Google Scholar] [CrossRef]
- Colorado, R.A.; Shumake, J.; Conejo, N.M.; Gonzalez-Pardo, H.; Gonzalez-Lima, F. Effects of maternal separation, early handling, and standard facility rearing on orienting and impulsive behavior of adolescent rats. Behav. Process. 2006, 71, 51–58. [Google Scholar] [CrossRef]
- Kwak, H.R.; Lee, J.W.; Kwon, K.J.; Kang, C.D.; Cheong, I.Y.; Chun, W.; Kim, S.S.; Lee, H.J. Maternal social separation of adolescent rats induces hyperactivity and anxiolytic behavior. Korean J. Physiol. Pharmacol. 2009, 13, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Yu, Z.; Hung, Q.; Xu, H. Attachment Insecurity in Rats Subjected to Maternal Separation and Early Weaning: Sex Differences. Front. Behav. Neurosci. 2021, 15, 637678. [Google Scholar] [CrossRef]
- Wang, D.; Levine, J.L.S.; Avila-Quintero, V.; Bloch, M.; Kaffman, A. Systematic review and meta-analysis: Effects of maternal separation on anxiety-like behavior in rodents. Transl. Psychiatry 2020, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- González-Pardo, H.; Arias, J.L.; Gómez-Lázaro, E.; Taboada, I.L.; Conejo, N.M. Sex-Specific Effects of Early Life Stress on Brain Mitochondrial Function, Monoamine Levels and Neuroinflammation. Brain Sci. 2020, 10, 447. [Google Scholar] [CrossRef]
- Matthews, K.; Dalley, J.W.; Matthews, C.; Tsai, T.H.; Robbins, T.W. Periodic maternal separation of neonatal rats produces region-and gender-specific effects on biogenic amine content in postmortem adult brain. Synapse 2001, 40, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Novak, G.; Fan, T.; O’Dowd, B.F.; George, S.R. Postnatal maternal deprivation and pubertal stress have additive effects on dopamine D2 receptor and CaMKII beta expression in the striatum. Int. J. Dev. Neurosci. 2013, 31, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Kiss Von Soly, S.; Ratnayake, U.; Klug, M.; Binder, M.D.; Hannan, A.J.; van den Buuse, M. Long-term effects of combined neonatal and adolescent stress on brain-derived neurotrophic factor and dopamine receptor expression in the rat forebrain. Biochim. Biophys Acta 2014, 1842, 2126–2135. [Google Scholar] [CrossRef] [PubMed]
- Knight, P.; Chellian, R.; Wilson, R.; Behnood-Rod, A.; Panunzio, S.; Bruijnzeel, A.W. Sex differences in the elevated plus-maze test and large open field test in adult Wistar rats. Pharmacol. Biochem. Behav. 2021, 204, 173168. [Google Scholar] [CrossRef]
- Rodina, V.I.; Krupina, N.A.; Kryzhanovskii, G.N.; Oknina, N.B. A new method of evaluating anxiety states and phobias in rats. Bull. Exp. Biol. Med. 1992, 114, 916–920. [Google Scholar] [CrossRef]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef]
- Ari, C.; D’Agostino, D.P.; Diamond, D.M.; Kindy, M.; Park, C.; Kovács, Z. Elevated Plus Maze Test Combined with Video Tracking Software to Investigate the Anxiolytic Effect of Exogenous Ketogenic Supplements. J. Vis. Exp. 2019, 143, e58396. [Google Scholar] [CrossRef]
- Niesink, R.J.M.; Van Ree, J.M. Involvement of opioid and dopaminergic systems in isolation-induced pinning and social grooming of young rats. Neuropharmacology 1989, 28, 411–418. [Google Scholar] [CrossRef]
- McKibben, C.E.; Reynolds, G.P.; Jenkins, T.A. Analysis of sociability and preference for social novelty in the acute and subchronic phencyclidine rat. J. Psychopharmacol. 2014, 28, 955–963. [Google Scholar] [CrossRef]
- Scott, L.N.; Levy, K.N.; Granger, D.A. Biobehavioral reactivity to social evaluative stress in women with borderline personality disorder. Pers. Disord. 2013, 4, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Niedtfeld, I. Experimental investigation of cognitive and affective empathy in borderline personality disorder: Effects of ambiguity in multimodal social information processing. Psychiatry Res. 2017, 253, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, M.S.; Imani, A.; Meknatkhah, S.; Riazi, G. Correlation between adolescent chronic emotional stress and incidence of adult cardiovascular disease in female rats. Iran J. Basic Med. Sci. 2019, 22, 1179–1185. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2007; p. 456. [Google Scholar]
- Chiu, K.; Lau, W.M.; Lau, H.T.; So, K.F.; Chang, R.C. Micro-dissection of rat brain for RNA or protein extraction from specific brain region. J. Vis. Exp. 2007, 7, 269. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Royal Statistical. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Li, M.; Xue, X.; Shao, S.; Shao, F.; Wang, W. Cognitive, emotional and neurochemical effects of repeated maternal separation in adolescent rats. Brain Res. 2013, 1518, 82–90. [Google Scholar] [CrossRef]
- Jin, S.; Zhao, Y.; Jiang, Y.; Wang, Y.; Li, C.; Zhang, D.; Lian, B.; Du, Z.; Sun, H.; Sun, L. Anxiety-like behaviour assessments of adolescent rats after repeated maternal separation during early life. NeuroReport 2018, 29, 643–649. [Google Scholar] [CrossRef]
- Khalifeh, S.; Khodagholi, F.; Moghtadaei, M.; Behvarmanesh, A.; Kheradmand, A.; Ghazvini, H. Effects of Maternal Deprivation on Anxiety, Depression, and Empathy in Male and Female Offspring of Wistar Rats in the Face of Novel Objects. Galen Med. J. 2019, 8, e1093. [Google Scholar] [CrossRef]
- Lee, N.; Oh, S.J.; Park, J.W.; Nam, K.R.; Kang, K.J.; Lee, K.C.; Lee, Y.J.; Choi, J.S.; Seok, J.H.; Choi, J.Y. Evaluation of the Effects of Developmental Trauma on Neurotransmitter Systems Using Functional Molecular Imaging. Int. J. Mol. Sci. 2021, 22, 2522. [Google Scholar] [CrossRef]
- Oh, S.J.; Lee, N.; Nam, K.R.; Kang, K.J.; Lee, K.C.; Lee, Y.J.; Seok, J.H.; Choi, J.Y. Effect of developmental stress on the in vivo neuronal circuits related to excitation-inhibition balance and mood in adulthood. Front. Psychiatry 2023, 14, 1086370. [Google Scholar] [CrossRef]
- Ohta, K.; Miki, T.; Warita, K.; Suzuki, S.; Kusaka, T.; Yakura, T.; Liu, J.Q.; Tamai, M.; Takeuchi, Y. Prolonged maternal separation disturbs the serotonergic system during early brain development. Int. J. Dev. Neurosci. 2014, 33, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, G.S.; Girardi, C.E.; de Oliveira, A.S.; Monteiro Longo, B.; Suchecki, D. Maternal deprivation alters growth, food intake, and neuropeptide Y in the hypothalamus of adolescent male and female rats. Dev Psychobiol. 2016, 58, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.A.; Almeida, L.C.D.A.; Tavares, G.A.; Falcão, L.D.A.L.; Beltrão, L.C.; Costa, F.C.O.; de Souza, F.L.; da Silva, M.C.; de Souza, S.L. Dual exposure to stress in different stages of development affects eating behavior of male Wistar rats. Physiol. Behav. 2020, 214, 112769. [Google Scholar] [CrossRef] [PubMed]
- de Souza, J.A.; da Silva, M.C.; Costa, F.C.O.; de Matos, R.J.B.; Campina, R.C.D.F.; Almeida, L.C.D.A.; da Silva, A.A.M.; Cavalcante, T.C.F.; Tavares, G.A.; de Souza, S.L. Early life stress induced by maternal separation during lactation alters the eating behavior and serotonin system in middle-aged rat female offspring. Pharm. Biochem. Behav. 2020, 192, 172908. [Google Scholar] [CrossRef]
- Kuma, H.; Miki, T.; Matsumoto, Y.; Gu, H.; Li, H.P.; Kusaka, T.; Satriotomo, I.; Okamoto, H.; Yokoyama, T.; Bedi, K.S.; et al. Early maternal deprivation induces alterations in brain-derived neurotrophic factor expression in the developing rat hippocampus. Neurosci. Lett. 2004, 372, 68–73. [Google Scholar] [CrossRef]
- Gołyszny, M.; Zieliński, M.; Paul-Samojedny, M.; Filipczyk, Ł.; Pałasz, A.; Obuchowicz, E. Escitalopram alters the hypotha-lamic OX system but does not affect its up-regulation induced by early-life stress in adult rats. Neurosci. Res. 2022, 180, 58–71. [Google Scholar] [CrossRef]
- Yadav, V.K.; Oury, F.; Tanaka, K.F.; Thomas, T.; Wang, Y.; Cremers, S.; Hen, R.; Krust, A.; Chambon, P.; Karsenty, G. Leptin-dependent serotonin control of appetite: Temporal specificity, transcriptional regulation, and therapeutic implications. J. Exp. Med. 2011, 208, 41–52. [Google Scholar] [CrossRef] [PubMed]
- de Lima, R.M.S.; Dos Santos Bento, L.V.; di Marcello Valladão Lugon, M.; Barauna, V.G.; Bittencourt, A.S.; Dalmaz, C.; de Vasconcellos Bittencourt, A.P.S. Early life stress and the programming of eating behavior and anxiety: Sex-specific relation-ships with serotonergic activity and hypothalamic neuropeptides. Behav. Brain Res. 2020, 379, 112399. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Sex differences in the physiology of eating. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1215–R1267. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, Y.; Li, X.; Liu, X.; Qin, C. Early-Life Neglect Alters Emotional and Cognitive Behavior in a Sex-Dependent Manner and Reduces Glutamatergic Neuronal Excitability in the Prefrontal Cortex. Front. Psychiatry 2021, 11, 572224. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.J.; Yang, Y.; Wang, L.P.; Xu, L.; Mao, R.R. Maternal separation exaggerates spontaneous recovery of extinguished contextual fear in adult female rats. Behav. Brain Res. 2014, 269, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, W.M.; Riley, E.P.; Ehlers, C.L. Behavioral and electrophysiological effects of early repeated maternal separation. Depression 1994, 2, 43–53. [Google Scholar] [CrossRef]
- Marmendal, M.; Eriksson, C.J.; Fahlke, C. Early deprivation increases exploration and locomotion in adult male Wistar offspring. Pharm. Biochem. Behav. 2006, 85, 535–544. [Google Scholar] [CrossRef]
- Alves, R.L.; Oliveira, P.; Lopes, I.M.; Portugal, C.C.; Alves, C.J.; Barbosa, F.; Summavielle, T.; Magalhães, A. Early-life stress affects drug abuse susceptibility in adolescent rat model independently of depression vulnerability. Sci. Rep. 2020, 10, 13326. [Google Scholar] [CrossRef] [PubMed]
- Kent, M.; Bardi, M.; Hazelgrove, A.; Sewell, K.; Kirk, E.; Thompson, B.; Trexler, K.; Terhune-Cotter, B.; Lambert, K. Profiling coping strategies in male and female rats: Potential neurobehavioral markers of increased resilience to depressive symptoms. Horm. Behav. 2017, 95, 33–43. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, J.W.; Yim, S.V.; Kim, M.J.; Kim, S.A.; Kim, Y.J.; Kim, C.J.; Chung, J.H. Fluoxetine enhances cell proliferation and prevents apoptosis in dentate gyrus of maternally separated rats. Mol. Psychiatry 2001, 6, 725–728. [Google Scholar] [CrossRef]
- Lundberg, S.; Martinsson, M.; Nylander, I.; Roman, E. Altered corticosterone levels and social play behavior after prolonged maternal separation in adolescent male but not female Wistar rats. Horm. Behav. 2017, 87, 137–144. [Google Scholar] [CrossRef]
- Veenema, A.H.; Neumann, I.D. Maternal separation enhances offensive play-fighting, basal corticosterone and hypothalamic vasopressin mRNA expression in juvenile male rats. Psychoneuroendocrinology 2009, 34, 463–467. [Google Scholar] [CrossRef]
- Veenema, A.H.; Blume, A.; Niederle, D.; Buwalda, B.; Neumann, I.D. Effects of early life stress on adult male aggression and hypothalamic vasopressin and serotonin. Eur. J. Neurosci. 2006, 24, 1711–1720. [Google Scholar] [CrossRef]
- Ferris, C.F.; Stolberg, T.; Kulkarni, P.; Murugavel, M.; Blanchard, R.; Blanchard, D.C.F.; Febo, M.; Brevard, M.; Simon, N.G. Imaging the neural circuitry and chemical control of aggressive motivation. BMC Neurosci. 2008, 9, 111. [Google Scholar] [CrossRef]
- van Erp, A.M.; Miczek, K.A. Increased accumbal dopamine during daily alcohol consumption and subsequent aggressive behavior in rats. Psychopharmacology 2007, 191, 679–688. [Google Scholar] [CrossRef]
- Diamantopoulou, A.; Raftogianni, A.; Stamatakis, A.; Alikaridis, F.; Oitzl, M.S.; Stylianopoulou, F. Denial of reward in the neonate shapes sociability and serotonergic activity in the adult rat. PLoS ONE 2012, 7, e33793. [Google Scholar] [CrossRef] [PubMed]
- Mintz, M.; Rüedi-Bettschen, D.; Feldon, J.; Pryce, C.R. Early social and physical deprivation leads to reduced social motivation in adulthood in Wistar rats. Behav. Brain Res. 2005, 156, 311–320. [Google Scholar] [CrossRef]
- Hulshof, H.J.; Novati, A.; Sgoifo, A.; Luiten, P.G.; de Boer, J.A.; Meerlo, P. Maternal separation decreases adult hippocampal cell proliferation and impairs cognitive performance but has little effect on stress sensitivity and anxiety in adult Wistar rats. Behav. Brain Res. 2011, 216, 552–560. [Google Scholar] [CrossRef]
- Karen, C.; Rajan, K.E. Social Behaviour and Epigenetic Status in Adolescent and Adult Rats: The Contribution of Early-Life Stressful Social Experience. Cell Mol. Neurobiol. 2019, 39, 371–385. [Google Scholar] [CrossRef]
- Kambali, M.Y.; Anshu, K.; Kutty, B.M.; Muddashetty, R.S.; Laxmi, T.R. Effect of early maternal separation stress on attention, spatial learning and social interaction behaviour. Exp. Brain Res. 2019, 237, 1993–2010. [Google Scholar] [CrossRef]
- Oades, R.D.; Sadile, A.G.; Sagvolden, T.; Viggiano, D.; Zuddas, A.; Devoto, P.; Aase, H.; Johansen, E.B.; Ruocco, L.A.; Russell, V.A. The control of responsiveness in ADHD by catecholamines: Evidence for dopaminergic, noradrenergic and interactive roles. Dev. Sci. 2005, 8, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Huppertz-Kessler, C.J.; Poeschl, J.; Hertel, R.; Unsicker, K.; Schenkel, J. Effects of a new postnatal stress model on monoaminergic neurotransmitters in rat brains. Brain Dev. 2012, 34, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Pagava, K.I.; Gogberishvili, K. Differences in modification of stress mechanisms in rat pups exposed to continuous and intermittent maternal deprivation. Bull. Exp. Biol. Med. 2004, 138, 202–204. [Google Scholar] [CrossRef]
- Adinolfi, A.; Zell, S.; Leo, D.; Carbone, C.; Mus, L.; Illiano, P.; Alleva, E.; Gainetdinov, R.R.; Adriani, W. Behavioral characterization of DAT-KO rats and evidence of asocial-like phenotypes in DAT-HET rats: The potential involvement of norepinephrine system. Behav. Brain Res. 2019, 359, 516–527. [Google Scholar] [CrossRef]
- Sonuga-Barke, E.J.S. Psychological heterogeneity in AD/HD--a dual pathway model of behavior and cognition. Behav. Brain Res. 2002, 130, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Chocyk, A.; Majcher-Maślanka, I.; Przyborowska, A.; Maćkowiak, M.; Wędzony, K. Early-life stress increases the survival of midbrain neurons during postnatal development and enhances reward-related and anxiolytic-like behaviors in a sex-dependent fashion. Int. J. Dev. Neurosci. 2015, 44, 33–47. [Google Scholar] [CrossRef]
- Sagvolden, T.; Johansen, E.B.; Aase, H.; Russell, V.A. A dynamic developmental theory of Attention-Deficit/Hyperactivity Disorder (ADHD) predominantly hyperactive/ impulsive subtype. Behav. Brain Sci. 2005, 28, 397–419. [Google Scholar] [CrossRef]
- Gondré-Lewis, M.C.; Bassey, R.; Blum, K. Pre-clinical models of reward deficiency syndrome: A behavioral octopus. Neurosci. Biobehav. Rev. 2020, 115, 164–188. [Google Scholar] [CrossRef] [PubMed]
- Viggiano, D.; Vallon, D.; Sadile, A. Dysfunctions in dopamine systems and ADHD: Evidence from animals and modeling. Neural Plast. 2004, 11, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Bacqué-Cazenave, J.; Bharatiya, R.; Barrière, G.; Delbecque, J.-P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdère, P. Serotonin in Animal Cognition and Behavior. Int. J. Mol. Sci. 2020, 21, 1649. [Google Scholar] [CrossRef]
- Llorente, R.; O’Shea, E.; Gutierrez-Lopez, M.D.; Llorente-Berzal, A.; Colado, M.I.; Viveros, M.P. Sex-dependent maternal deprivation effects on brain monoamine content in adolescent rats. Neurosci. Lett. 2010, 479, 112–117. [Google Scholar] [CrossRef]
- Rentesi, G.; Antoniou, K.; Marselos, M.; Fotopoulos, A.; Alboycharali, J.; Konstandi, M. Long-term consequences of early maternal deprivation in serotonergic activity and HPA function in adult rat. Neurosci. Lett. 2010, 480, 7–11. [Google Scholar] [CrossRef]
- Arborelius, L.; Eklund, M.B. Both long and brief maternal separation produces persistent changes in tissue levels of brain monoamines in middle-aged female rats. Neuroscience 2007, 145, 738–750. [Google Scholar] [CrossRef]
- Kosten, T.A.; Zhang, X.Y.; Kehoe, P. Infant rats with chronic neonatal isolation experience show decreased extracellular serotonin levels in ventral striatum at baseline and in response to cocaine. Dev. Brain Res. 2004, 152, 19–24. [Google Scholar] [CrossRef]
- Xue, X.; Shao, S.; Li, M.; Shao, F.; Wang, W. Maternal separation induces alterations of serotonergic system in different aged rats. Brain Res. Bull. 2013, 95, 15–20. [Google Scholar] [CrossRef]
- Dutcher, E.G.; Lopez-Cruz, L.; Pama, E.A.C.; Lynall, M.E.; Bevers, I.C.R.; Jones, J.A.; Khan, S.; Sawiak, S.J.; Milton, A.L.; Clatworthy, M.R.; et al. Early-life stress biases responding to negative feedback and increases amygdala volume and vulnerability to later-life stress. Transl. Psychiatry 2023, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Patki, G.; Salvi, A.; Liu, H.; Salim, S. Witnessing traumatic events and post-traumatic stress disorder: Insights from an animal model. Neurosci. Lett. 2015, 600, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Kaneyuki, H.; Yokoo, H.; Tsuda, A.; Yoshida, M.; Mizki, Y.; Yamada, M.; Tanaka, M. Psychological stress increases dopamine turnover selectively in mesoprefrontal dopamine neurons of rats: Reversal by diazepam. Brain Res. 1991, 557, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.F. Exposure to the stressor environment prevents the temporal dissipation of behavioral depression/learned helplessness. Biol. Psychiatry 2001, 49, 763–773. [Google Scholar] [CrossRef]
- Deutch, A.Y.; Tam, S.-Y.; Roth, R.H. Footshock and conditioned stress increase 3,4-dihydroxyphenylacetic acid (DOPAC) in the ventral tegmental area but not substantia nigra. Brain Res. 1985, 333, 143–146. [Google Scholar] [CrossRef]
- Inoue, T.; Koyama, T.; Yamashita, I. Effect of conditioned fear stress on serotonin metabolism in the rat brain. Pharmacol. Biochem. Behav. 1993, 44, 371–374. [Google Scholar] [CrossRef]
- Roth, R.H.; Tam, S.-Y.; Ida, Y.; Yang, J.-X.; Deutch, A.Y. Stress and the mesocorticolimbic dopamine systems. Ann. N. Y. Acad. Sci. 1988, 537, 138–147. [Google Scholar] [CrossRef]
- Kim, J.; Park, M.; Lee, C.; Ha, J.J.; Choi, J.S.; Kim, C.H.; Seok, J.H. Maladaptive Alterations of Defensive Response Following Developmental Complex Stress in Rats. Clin. Psychopharmacol. Neurosci. 2020, 18, 412–422. [Google Scholar] [CrossRef]
- Daniels, W.M.; Pietersen, C.Y.; Carstens, M.E.; Stein, D.J. Maternal separation in rats leads to anxiety-like behavior and a blunted ACTH response and altered neurotransmitter levels in response to a subsequent stressor. Metab. Brain Dis. 2004, 19, 3–14. [Google Scholar] [CrossRef]
- Oosterhof, C.A.; El Mansari, M.; Merali, Z.; Blier, P. Altered monoamine system activities after prenatal and adult stress: A role for stress resilience? Brain Res. 2016, 1642, 409–418. [Google Scholar] [CrossRef]
- Macrì, S.; Würbel, H. Developmental plasticity of HPA and fear responses in rats: A critical review of the maternal mediation hypothesis. Horm. Behav. 2006, 50, 667–680. [Google Scholar] [CrossRef]
- Li, M.; Fu, X.; Xie, W.; Guo, W.; Li, B.; Cui, R.; Yang, W. Effect of Early Life Stress on the Epigenetic Profiles in Depression. Front. Cell Dev. Biol. 2020, 8, 867. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Fanibunda, S.E.; Kapri, D.; Vasaya, S.; Pati, S.; Vaidya, V.A. GPCR signaling: Role in mediating the effects of early adversity in psychiatric disorders. FEBS J. 2021, 288, 2602–2621. [Google Scholar] [CrossRef] [PubMed]
- Johnson, F.K.; Kaffman, A. Early life stress perturbs the function of microglia in the developing rodent brain: New insights and future challenges. Brain Behav. Immun. 2018, 69, 18–27. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
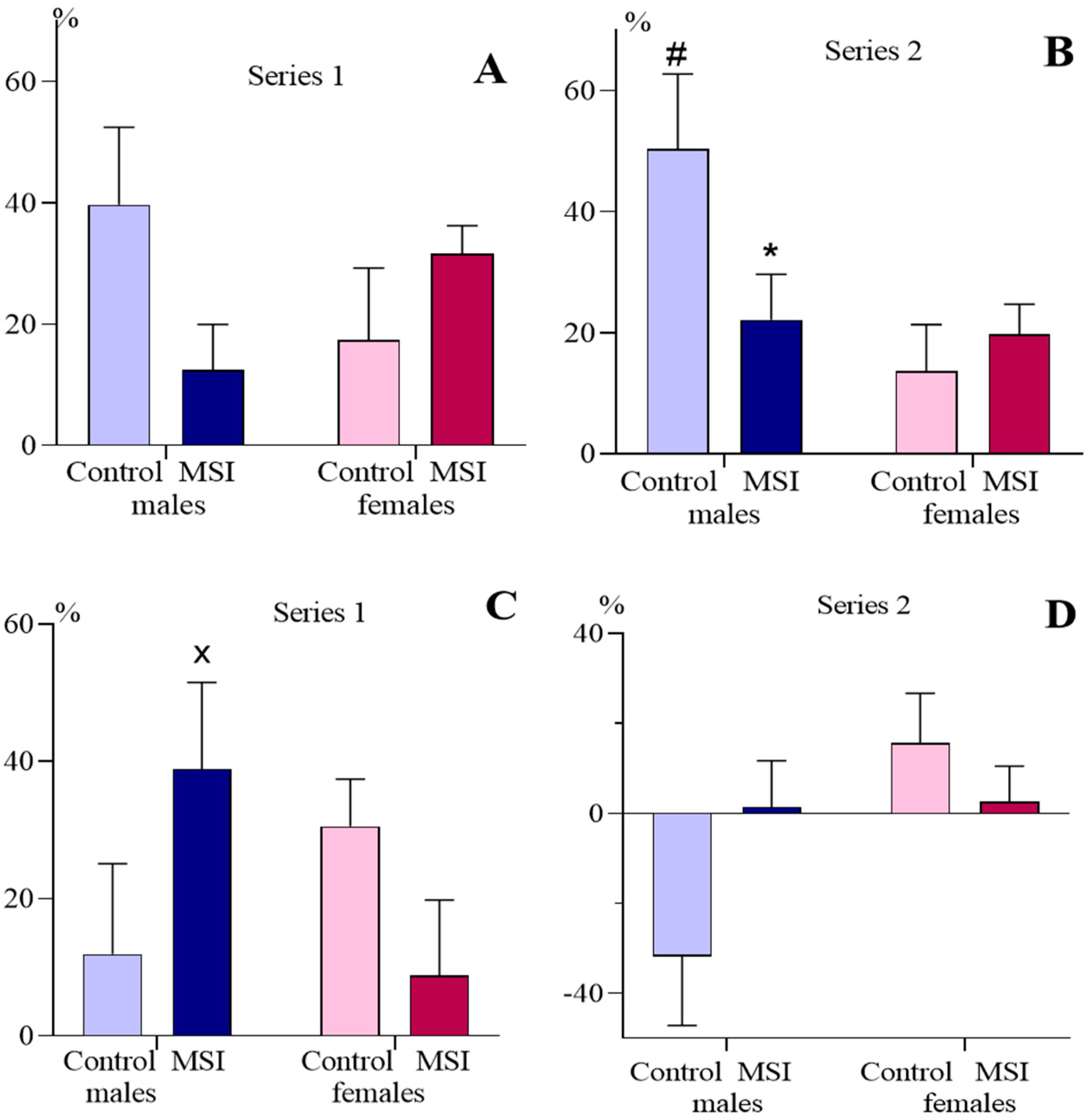{kind=link}
| Control, m | MSI, m | Control, f | MSI, f | |||
|---|---|---|---|---|---|---|
| Series 1 | ||||||
| P28 | n | 10 | 11 | 9 | 10 | |
| weight | 33.0 ± 0.7 | 30.7 ± 2.4 | 32.3 ± 1.1 | 31.3 ± 1.8 | ||
| ANOVA | Sex main effect: F (1, 36) = 0.001, p = 0.975; Rearing main effect: F (1, 36) = 0.869, p = 0.357 | |||||
| P56 | n | 12 | 16 | 13 | 12 | |
| weight | 160.6 ± 6.4 | 149.4 ± 6.2 | 146.9 ± 7.1 | 149.0 ± 5.2 | ||
| ANOVA | Sex main effect: F (1, 36) = 1.636, p = 0.207; Rearing main effect: F (1, 36) = 0.289, p = 0.594 | |||||
| Series 2 | ||||||
| P58 | n | 26 | 24 | 24 | 22 | |
| weigh | 172.7 ± 6.6 | 162.5 ± 5.9 | 159.6 ± 5.8 | 150.1 ± 5.0 | ||
| ANOVA | Sex main effect: F (1, 92) = 4.60, p = 0.035; Rearing main effect: F (1, 92) = 2.781, p = 0.099 | |||||
| P75 | weigh | 310.7 ± 7.3 | 304.9 ± 7.1 | 252.9 ± 5.6 # | 232.1 ± 5.0 * # | |
| ANOVA | Sex main effect: F (1, 92) = 9.798, p = 0.003; Rearing main effect: F (1, 92) = 4.250, p = 0.042 | |||||
| 1 Month | |||||
|---|---|---|---|---|---|
| Control, m | MSI, m | Control, f | MSI, f | ||
| Frontal cortex | 1.79 ± 0.06 | 1.94 ± 0.08 | 1.88 ± 0.14 | 1.89 ± 0.09 | |
| Hippocampus | 1.11 ± 0.06 | 1.38 ± 0.05 * | 1.19 ± 0.09 | 1.27 ± 0.07 | |
| Rearing main effect: F (1, 36) = 6.199, p = 0.017 | |||||
| Striatum | 2.44 ± 0.24 | 2.68 ± 0.22 | 2.49 ± 0.18 | 1.99 ± 0.25 | |
| Hypothalamus | 6.89 ± 0.15 | 7.86 ± 0.24 * # | 6.91 ± 0.24 | 6.46 ± 0.20 | |
| Sex main effect: F (1, 36) = 9.798, p = 0.003; Sex × Rearing interaction: F (1, 36) = 12.312, p = 0.001 | |||||
| 2 months | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Frontal cortex | 2.61 ± 0.12 | 2.62 ± 0.06 | 2.77 ± 0.10 | 2.50 ± 0.12 | |
| Hippocampus | 2.22 ± 0.09 | 1.94 ± 0.08 * | 1.96 ± 0.08 # | 1.81 ± 0.08 | |
| Sex main effect: F (1,49) = 4.757.3, p = 0.034; Rearing main effect: F (1, 49)= 6.078, p = 0.017 * p = 0.063 | |||||
| Striatum | 3.40 ± 0.53 | 4.11 ± 0.32 | 2.98 ± 0.34 | 3.00 ± 0.51 | |
| Hypothalamus | 10.51 ± 0.37 | 10.61 ± 0.30 | 11.47 ± 0.59 | 9.61 ± 0.17 * | |
| Sex × Rearing interaction: F (1, 48) = 4.871, p = 0.032 | |||||
| 3 months (after double hit) | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Frontal cortex | FS | 1.82 ± 0.07 | 2.01 ± 0.07 | 2.41 ± 0.07 # | 2.28 ± 0.08 # |
| EmS | 1.99 ± 0.07 | 2.03 ± 0.09 | 2.60 ± 0.08 # | 2.38 ± 0.07 * # | |
| Sex main effect: F (1, 88) = 72.170, p < 0.001; Stress main effect: F (1, 88) = 5.150, p = 0.010; Sex × Rearing interaction: F (1, 88) = 6.889, p = 0.018 | |||||
| Hippocampus | FS | 2.33 ± 0.05 | 2.22 ± 0.09 | 2.30 ± 0.09 | 2.14 ± 0.10 |
| EmS | 2.66 ± 0.17 | 2.26 ± 0.07 * | 2.44 ± 0.11 | 2.34 ± 0.13 | |
| Rearing main effect: F (1, 88)= 6.699, p = 0.011; Stress main effect: F (1, 88) = 5.65, p = 0.020 | |||||
| Striatum | FS | 4.18 ± 0.47 | 2.79 ± 0.26 | 2.33 ± 0.33 # | 2.92 ± 0.21 |
| EmS | 3.36 ± 0.20 s | 2.59 ± 0.31 | 2.54 ± 0.28 | 3.17 ± 0.35 | |
| Sex main effect: F (1, 88) = 4.775, p = 0.031; Rearing main effect: F (1, 88) = 14.496, p < 0.001 | |||||
| Hypothalamus | FS | 11.01 ± 0.58 | 10.85 ± 0.46 | 10.43 ± 0.46 | 10.45 ± 0.39 |
| EmS | 12.61 ± 0.97 | 11.19 ± 0.51 | 9.94 ± 0.55 | 9.99 ± 0.31 # | |
| Rearing main effect: F (1, 88) = 8.702, p = 0.004; Sex × stress interaction: F (1, 88) = 3.080, p = 0.083 | |||||
| 1 Month | |||||
|---|---|---|---|---|---|
| Frontal Cortex | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 0.71 ± 0.33 | 0.49 ± 0.12. | 0.54 ± 0.11 | 0.44 ± 0.02 | |
| DOPAC | 0.22 ± 0.09 | 0.17 ± 0.02 | 0.18 ± 0.02 | 0.15 ± 0.01 | |
| HVA | 0.43 ± 0.10 | 0.31 ± 0.04 | 0.47 ± 0.07 | 0.27 ± 0.02 * | |
| Rearing main effect: F (1, 36) = 5.923, p = 0.020 | |||||
| DOPAC/DA | 0.36 ± 0.03 | 0.38 ± 0.03 | 0.35 ± 0.03 | 0.33 ± 0.02 | |
| HVA/DA | 0.84 ± 0.11 | 0.72 ± 0.09 | 0.95 ± 0.11 | 0.62 ± 0.05 * | |
| Rearing main effect: F (1, 36) = 5.303, p = 0.027 | |||||
| Hippocampus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 0.10 ± 0.02 | 0.12 ± 0.04. | 0.15 ± 0.03 | 0.09 ± 0.02 | |
| DOPAC | 0.05 ± 0.00 | 0.05 ± 0.02 | 0.06 ± 0.03 | 0.04 ± 0.00 | |
| HVA | nd | nd | nd | nd | |
| DOPAC/DA | 0.57 ± 0.06 | 0.48 ± 0.05 | 0.49 ± 0.05 | 0.56 ± 0.08 | |
| Striatum | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 26.85 ± 2.89 | 21.02 ± 2.05 | 23.41 ± 3.41 | 27.15 ± 2.54 | |
| DOPAC | 8.97 ± 0.89 | 6.17 ± 0.91 * | 4.06 ± 0.53 # | 5.43 ± 0.41 | |
| Sex main effect: F (1, 36) = 14.393, p < 0.001; Sex × Rearing interaction: F (1, 36) = 7.842, p = 0.008 | |||||
| HVA | 3.88 ± 0.49 | 2.63 ± 0.29 * | 2.57 ± 0.24 # | 2.83 ± 0.18 | |
| Sex × Rearing interaction: F (1, 36) = 5.040, p = 0.031 | |||||
| DOPAC/DA | 0.34 ± 0.02 | 0.30 ± 0.03 | 0.18 ± 0.01 # | 0.20 ± 0.01 # | |
| Sex main effect: F (1, 36) = 40.406, p < 0.001 | |||||
| HVA/DA | 0.15 ± 0.01 | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 | |
| Sex main effect: F (1, 36) = 6.705, p = 0.0138; Rearing main effect: F (1, 36) = 3.908, p = 0.055 | |||||
| Hypothalamus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 2.35 ± 0.18 | 1.71 ± 0.14 * | 1.80 ± 0.19 | 1.87 ± 0.15 | |
| Sex × Rearing interaction: F (1, 36) = 4.673, p = 0.037 | |||||
| DOPAC | 0.53 ± 0.05 | 0.38 ± 0.03 | 0.43 ± 0.04 | 0.52 ± 0.05 | |
| Sex × Rearing interaction: F (1, 36) = 8.010, p = 0.008 | |||||
| HVA | 0.21 ± 0.04 | 0.16 ± 0.01 | 0.13 ± 0.01 | 0.19 ± 0.01 | |
| Sex × Rearing interaction: F (1, 36) = 6.198, p = 0.018 | |||||
| DOPAC/DA | 0.22 ± 0.01 | 0.23 ± 0.01 | 0.24 ± 0.01 | 0.28 ± 0.01 * | |
| Sex main effect: F (1, 36) = 14.823, p < 0.001; Rearing main effect: F (1, 36) = 4.976, p = 0.032; Sex × Rearing interaction: F (1, 36) = 3.555, p = 0.067 | |||||
| HVA/DA | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.08 ± 0.01 | 0.11 ± 0.01 | |
| Rearing main effect: F (1, 36) = 3.384, p = 0.074 | |||||
| 2 months | |||||
| Frontal cortex | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 0.89 ± 0.22 | 0.84 ± 0.09 | 0.66 ± 0.04 | 0.55 ± 0.03 | |
| DOPAC | 0.19 ± 0.03 | 0.20 ± 0.02 | 0.13 ± 0.01 | 0.12 ± 0.01 | |
| HVA | 0.29 ± 0.03 | 0.30 ± 0.03 | 0.21 ± 0.03 | 0.22 ± 0.02 | |
| Sex main effect: F (1, 49) = 8.508, p = 0.005 | |||||
| DOPAC/DA | 0.23 ± 0.03 | 0.24 ± 0.03 | 0.20 ± 0.03 | 0.22 ± 0.02 | |
| Sex main effect: F (1, 49) = 4.800, p = 0.033 | |||||
| HVA/DA | 0.84 ± 0.11 | 0.72 ± 0.09 | 0.95 ± 0.11 | 0.62 ± 0.05 | |
| Hippocampus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 0.16 ± 0.02 | 0.17 ± 0.03 | 0.63 ± 0.49 | 0.14 ± 0.02 | |
| DOPAC | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.14 ± 0.07 | 0.07 ± 0.01 | |
| HVA | nd | nd | nd | nd | |
| DOPAC/DA | 0.26 ± 0.02 | 0.30 ± 0.03 | 0.48 ± 0.05 # | 0.56 ± 0.06 | |
| Sex main effect: F (1, 49) = 41.116, p < 0.001 | |||||
| Striatum | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 37.37 ± 4.19 | 30.01 ± 3.17 | 47.30 ± 3.85 | 35.07 ± 3.26 * | |
| Sex main effect: F (1, 49) = 4.525, p = 0.038; Rearing main effect: F (1, 49) = 7.958, p = 0.007 | |||||
| DOPAC | 7.97 ± 1.00 | 6.46 ± 0.64 | 8.15 ± 0.57 | 9.12 ± 0.73 | |
| Sex × Rearing interaction: F (1, 49) = 3.626, p = 0.063 | |||||
| HVA | 3.25 ± 0.45 | 2.08 ± 0.22 * | 3.43 ± 0.28 | 3.43 ± 0.39 | |
| Sex main effect: F (1, 49) = 5.026, p = 0.029; Rearing main effect: F (1, 49) = 3.142, p = 0.082; Sex × Rearing interaction: F (1, 49) = 3.900, p = 0.054 | |||||
| DOPAC/DA | 0.21 ± 0.01 | 0.22 ± 0.01 | 0.18 ± 0.01 # | 0.27 ± 0.01 * # | |
| Rearing main effect: F (1, 49) = 19.378, p < 0.001; Sex × Rearing interaction: F (1, 49) = 18.895, p < 0.001 | |||||
| HVA/DA | 0.09 ± 0.01 | 0.07 ± 0.01 * | 0.07 ± 0.01 | 0.10 ± 0.01 | |
| Sex main effect: F (1, 49) = 8.221, p = 0.006; Rearing main effect: F (1, 49) = 4.614, p = 0.037; Sex × Rearing interaction: F (1, 49) = 4.568, p = 0.037 | |||||
| Hypothalamus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | 2.40 ± 0.13 | 2.20 ± 0.09 | 2.25 ± 0.19 | 2.15 ± 0.10 | |
| DOPAC | 0.49 ± 0.03 | 0.46 ± 0.03 | 0.56 ± 0.05 | 0.44 ± 0.02 | |
| Rearing main effect: F (1, 49) = 4.853. p = 0.032 | |||||
| HVA | 0.25 ± 0.01 | 0.15 ± 0.01 * | 0.34 ± 0.03 # | 0.22 ± 0.01 * # | |
| Sex main effect: F (1, 49) = 23.242, p < 0.001; Rearing main effect: F (1, 49) = 44.449, p < 0.001 | |||||
| DOPAC/DA | 0.21 ± 0.01 | 0.21 ± 0.01 | 0.25 ± 0.01 # | 0.21 ± 0.01 * | |
| Sex main effect: F (1, 49)= 4.165, p = 0.036; Rearing main effect: F (1, 49) = 4.443, p = 0.040; Sex × Rearing interaction: F (1, 49) = 6.979, p = 0.011 | |||||
| HVA/DA | 0.11 ± 0.01 | 0.07 ± 0.0.01 * | 0.15 ± 0.01 # | 0.11 ± 0.01* # | |
| Sex main effect: F (1, 49)= 33.821, p < 0.001; Rearing main effect: F (1, 49) = 36.799, p < 0.001 | |||||
| 3 months (after double hit) | |||||
| Frontal Cortex | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | FS | 1.83 ± 0.52 | 0.98 ± 0.28 | 1.45 ± 0.36 | 1.82 ± 0.47 |
| EmS | 1.43 ±0.43 | 1.72 ± 0.38 | 1.18 ± 0.19 | 1.20 ± 0.25 | |
| DOPAC | FS | 0.22 ± 0.05 | 0.16 ± 0.04 | 0.21 ± 0.04 | 0.26 ± 0.05 |
| EmS | 0.20 ± 0.07 | 0.23 ± 0.05 | 0.16 ± 0.03 | 0.14 ± 0.02 | |
| HVA | FS | 0.21 ± 0.03 | 0.21 ± 0.03 | 0.22 ±0.04 | 0.28 ± 0.04 |
| EmS | 0.24 ± 0.06 | 0.23 ± 0.04 | 0.19 ± 0.03 | 0.23 ± 0.02 | |
| DOPAC/DA | FS | 0.14 ± 0.01 | 0.17 ± 0.01 | 0.16 ± 0.01 | 0.17 ± 0.02 |
| EmS | 0.14 ± 0.01 | 0.13 ± 0.01 (s) | 0.13 ± 0.01 | 0.13 ± 0.01 s | |
| Stress main effect: F (1, 88) = 14.194, p < 0.001; Rearing × stress interaction: F (1, 88) = 3.781, p = 0.055 | |||||
| HVA/DA | FS | 0.18 ± 0.03 | 0.24 ± 0.03 | 0.21 ± 0.05 | 0.21 ± 0.03 |
| EmS | 0.20 ± 0.02 | 0.16 ± 0.02 | 0.17 ± 0.02 | 0.23 ± 0.03 | |
| Sex × Rearing × stress interaction: F (1, 88) = 5.189, p = 0.025 | |||||
| Hippocampus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | FS | 0.17 ± 0.02 | 0.30 ± 0.05 | 0.19 ± 0.02 | 0.28 ± 0.05 |
| EmS | 0.26 ± 0.06 | 0.27 ± 0.05 | 0.16 ± 0.01 | 0.25 ± 0.03 | |
| DOPAC | FS | 0.04 ± 0.00 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.06 ± 0.01 |
| EmS | 0.07 ± 0.01 | 0.08 ± 0.01 | 0.05 ± 0.00 | 0.06 ± 0.01 | |
| Rearing main effect: F (1, 88) = 5.294, p = 0.015 | |||||
| DOPAC/DA | FS | 0.26 ± 0.01 | 0.30 ± 0.03 | 0.30 ± 0.01 | 0.27 ± 0.03 |
| EmS | 0.31 ± 0.02 | 0.31 ± 0.04 | 0.33 ± 0.02 | 0.25 ± 0.02 | |
| Sex × Rearing interaction: F (1, 88) = 4.532, p = 0.036 | |||||
| Striatum | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | FS | 33.67 ± 5.70 | 31.64± 4.25 | 47.73 ± 5.38 | 40.09 ± 7.27 |
| EmS | 43.53 ± 3.94 | 38.87± 5.38 | 53.99 ± 4.66 | 46.29 ± 4.92 | |
| Sex main effect: F (1, 88) = 7.440, p = 0.008; Stress main effect: F (1, 88) = 3.982, p = 0.041 | |||||
| DOPAC | FS | 3.30 ± 0.55 | 3.38 ± 0.50 | 4.47 ± 0.47 | 3.61 ± 0.64 |
| EmS | 4.53 ± 0.47 | 4.01 ± 0.57 | 5.40 ± 0.57 | 4.38 ± 0.51 | |
| Stress main effect: F (1, 88) = 5.472, p = 0.021 | |||||
| HVA | FS | 1.94 ± 0/30 | 1.87 ± 0.27 | 2.62 ± 0.26 | 2.43 ± 0.33 |
| EmS | 2.62 ± 0.26 | 2.28 ± 0.32 | 2.83 ± 0.20 | 3.13 ± 0.39 | |
| Sex main effect: F (1, 88) = 7.780, p = 0.006; Stress main effect: F (1, 88) = 5.951, p = 0.017 | |||||
| DOPAC/DA | FS | 0.10 ± 0.00 | 0.11 ± 0.00 | 0.09 ± 0.00 | 0.09 ± 0.00 |
| EmS | 0.10 ± 0.00 | 0.10 ± 0.00 | 0.10 ± 0.01 | 0.09 ± 0.00 | |
| Sex main effect: F (1, 88) = 9.319, p = 0.003 | |||||
| HVA/DA | FS | 0.07 ± 0.02 | 0.07 ± 0.01 | 0.05 ± 0.00 | 0.09 ± 0.02 |
| EmS | 0.06 ± 0.00 | 0.06 ± 0.00 | 0.05 ± 0.00 | 0.07 ± 0.00 | |
| Sex × Rearing interaction: F (1, 88) = 3.692, p = 0.058 | |||||
| Hypothalamus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| Dopamine | FS | 2.27 ± 0.22 | 1.92 ± 0.13 | 2.23 ± 0.41 | 2.11 ± 0.12 |
| EmS | 2.26 ± 0.26 | 2.27 ± 0.25 | 2.48 ± 0.80 | 1.93 ± 0.12 | |
| DOPAC | FS | 0.41 ± 0.06 | 0.39 ± 0.02 | 0.53 ± 0.09 | 0.41 ± 0.02 |
| EmS | 0.41 ± 0.05 | 0.40 ± 0.05 | 0.49 ± 0.11 | 0.37 ± 0.02 | |
| HVA | FS | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.25 ± 0.02 | 0.18 ± 0.02 * |
| EmS | 0.12 ± 0.01 | 0.10 ± 0.01 | 0.21 ± 0.02 | 0.18 ± 0.02 | |
| Sex main effect: F (1, 88) = 85.001, p < 0.001; Rearing main effect: F (1, 88) = 7.048, p = 0.009; Sex × Rearing interaction: F (1, 88) = 3.048, p = 0.084 | |||||
| DOPAC/DA | FS | 0.18 ± 0.01 | 0.21 ± 0.01 | 0.24 ± 0.01 | 0.20 ± 0.01 |
| EmS | 0.18 ± 0.01 | 0.18 ± 0.01 | 0.22 ± 0.01 | 0.20 ± 0.01 | |
| Sex main effect: F (1, 88) = 17.931, p < 0.001;Rearing main effect: F (1, 88) = 3.259, p = 0.074; Stress main effect: F (1, 88) = 4.076, p = 0.046; Sex × Rearing interaction: F (1, 88) = 12.503, p < 0.001; Sex × Rearing × stress interaction: F (1, 88) = 2.962, p = 0.088 | |||||
| HVA/DA | FS | 0.04 ± 0.00 | 0.05 ± 0.01 | 0.13 ± 0.01 | 0.09 ± 0.01 |
| EmS | 0.06 ± 0.00 | 0.04 ± 0.00 | 0.12 ± 0.01 | 0.10 ± 0.01 | |
| Sex main effect: F (1, 88) = 76.719, p < 0.001; Rearing main effect: F (1, 88) = 3.364, p = 0.023; Sex × Rearing interaction: F (1, 88) = 12.503, p < 0.001 | |||||
| 1 Months | |||||
|---|---|---|---|---|---|
| Frontal Cortex | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 2.50 ± 0.01 | 2.42 ± 0.06 | 2.62 ± 0.09 | 2.53 ± 0.04 | |
| HIAA | 2.00 ± 0.13 | 1.65 ± 0.18 | 1.87 ± 0.08 | 1.79 ± 0.08 | |
| HIAA/5-HT | 0.8 ± 0.03 | 0.68 ± 0.07 | 0.72 ± 0.04 | 0.71 ± 0.04 | |
| Hippocampus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 2.25 ± 0.11 | 2.94 ± 0.40 | 2.71 ± 0.14 | 2.86 ± 0.09 | |
| Rearing main effect: F (1, 36) = 4.175, p = 0.034 | |||||
| HIAA | 2.24 ± 0.10 | 2.56 ± 0.12 | 2.71 ± 0.14 | 2.57 ± 0.10 | |
| Sex main effect: F (1, 36) = 4.271, p = 0.046; Sex × Rearing interaction: F (1, 36) = 4.010, p = 0.053 | |||||
| HIAA/5-HT | 1.00 ± 0.0 | 0.93 ± 0.06 | 1.01 ± 0.06 | 0.90 ± 0.02 | |
| Striatum | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 2.06 ± 0.07 | 2.26 ± 0.17 | 2.79 ± 0.20 # | 2.82 ± 0.20 # | |
| Sex main effect: F (1, 36) = 15.365, p < 0.001 | |||||
| HIAA | 4.55 ± 0.19 | 4.47 ± 0.27 | 4.80 ± 0.16 | 4.53 ± 0.31 | |
| HIAA/5-HT | 2.23 ± 0.11 | 2.06 ± 0.18 | 1.79 ± 0.13 # | 1.62 ± 0.06 | |
| Sex main effect: F (1, 36) = 12.703, p = 0.001 | |||||
| Hypothalamus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 6.25 ± 0.13 | 5.58 ± 0.22 * | 5.46 ± 0.20 # | 5.34 ± 0.12 | |
| Sex main effect: F (1, 36) = 9.347, p = 0.004; Rearing main effect: F (1, 36) = 5.467, p = 0.025 | |||||
| HIAA | 5.30 ± 0.19 | 4.48 ± 0.28 * | 4.97 ± 0.11 | 5.18 ± 0.23 | |
| Sex × Rearing interaction: F (1, 36) = 5.624, p = 0.023 | |||||
| HIAA/5-HT | 0.85 ± 0.03 | 0.79 ± 0.03 | 0.92 ± 0.03 | 0.97 ± 0.03 | |
| 2 months | |||||
| Frontal cortex | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 2.78 ± 0.08 | 3.28 ± 0.09 * | 3.11 ± 0.10 # | 2.41 ± 0.15 * # | |
| Sex main effect: F (1, 49) = 5.980, p = 0.018; Sex × Rearing interaction: F (1, 49) = 32.555, p < 0.001 | |||||
| HIAA | 1.38 ± 0.06 | 1.62 ± 0.06 * | 1.44 ± 0.05 | 1.05 ± 0.06 * | |
| Sex main effect: F (1, 49) = 18.435, p < 0.001; Sex × Rearing interaction: F (1, 49) = 31.328, p < 0.001 | |||||
| HIAA/5-HT | 0.5 ± 0.02 | 0.49 ± 0.01 | 0.47 ± 0.02 | 0.44 ± 0.02 | |
| Sex main effect: F (1, 49) = 6.116, p = 0.017 | |||||
| Hippocampus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 3.37 ± 0.08 | 3.37 ± 0.27 | 2.74 ± 0.14 | 2.59 ± 0.11 | |
| Sex main effect: F (1, 49) = 17.219, p < 0.001 | |||||
| HIAA | 1.67 ± 0.05 | 1.66 ± 0.06 | 1.77 ± 0.10 | 1.86 ± 0.06 | |
| Sex main effect: F (1, 49) = 4.856, p = 0.033 | |||||
| HIAA/5-HT | 0.50 ± 0.02 | 0.51 ± 0.03 | 0.66 ± 0.04 | 0.72 ± 0.02 | |
| Sex main effect: F (1, 49) = 55.357, p < 0.001 | |||||
| Striatum | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 2.70 ± 0.18 | 2.92 ± 0.14 | 2.45 ± 0.13 # | 2.42 ± 0.13 # | |
| Sex main effect: F (1, 49) = 5.210, p = 0.027 | |||||
| HIAA | 3.55 ± 0.15 | 3.87 ± 0.19 | 3.51 ± 0.14 | 3.85 ± 0.22 | |
| Rearing main effect: F (1, 49) = 3.577, p = 0.064 | |||||
| HIAA/5-HT | 1.37 ± 0.11 | 1.35 ± 0.06 | 1.49 ± 0.10 # | 1.64 ± 0.12 # | |
| Sex main effect: F (1, 49) = 4.094, p = 0.048 | |||||
| Hypothalamus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | 6.81 ± 0.45 | 5.95 ± 0.19 * | 6.51 ± 0.45 # | 6.46 ± 0.13 | |
| Sex × Rearing interaction: F (1, 49) = 3.289, p = 0.075 | |||||
| HIAA | 5.30 ± 0.19 | 4.48 ± 0.28 * | 4.97 ± 0.11 | 5.18 ± 0.23 | |
| Sex main effect: F (1, 49)= 10.471, p = 0.002; Rearing main effect: F (1, 49) = 6.302, p = 0.015 | |||||
| HIAA/5-HT | 0.58 ± 0.02 | 0.57 ± 0.02 | 0.69 ± 0.02 | 0.63 ± 0.02 * # | |
| Sex main effect: F (1, 49) = 22.276, p < 0.001; Rearing main effect: F (1, 49) = 4.053, p = 0.050; Sex × Rearing interaction: F (1, 49) = 3.686, p = 0.060 | |||||
| 3 months (after double hit) | |||||
| Frontal Cortex | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | FS | 3.51 ± 0.14 | 3.37 ± 0.14 | 3.70 ± 0.19 | 3.71 ± 0.13 |
| EmS | 3.21 ± 0.13 | 3.59 ± 0.13 | 3.62 ± 0.09 | 3.86 ± 0.09 | |
| Sex main effect: F (1, 88) = 9.933, p = 0.002; Rearing × stress interaction: F (1, 88) = 3.707, p = 0.057 | |||||
| HIAA | FS | 1.63 ± 0.09 | 1.43 ± 0.09 | 1.66 ± 0.05 | 1.43 ± 0.09 |
| EmS | 1.64 ± 0.10 | 1.63 ± 0.07 | 1.60 ± 0.07 | 1.44 ± 0.08 | |
| Rearing main effect: F (1, 88) = 6.884, p = 0.010 | |||||
| HIAA/5-HT | FS | 0.46 ± 0.01 | 0.42 ± 0.02 | 0.46 ± 0.02 | 0.39 ± 0.03 |
| EmS | 0.51 ± 0.02 | 0.46 ± 0.01 | 0.44 ± 0.02 | 0.38 ± 0.02 | |
| Sex main effect: F (1, 88) = 10.51, p = 0.002; Rearing main effect: F (1, 88) = 15.93, p < 0.001; Sex ×stress interaction: F (1, 88) = 3.32, p = 0.072 | |||||
| Hippocampus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | FS | 2.67 ± 0.26 | 2.96 ± 0.14 | 2.80 ± 0.26 | 3.48 ± 0.40 |
| EmS | 2.82 ± 0.10 | 3.21 ± 0.13 | 2.86 ± 0.25 | 2.95 ± 0.11 | |
| Rearing main effect: F (1, 88) = 5.156, p = 0.026 | |||||
| HIAA | FS | 1.42 ± 0.04 | 1.42 ± 0.06 | 1.78 ± 0.06 | 2.01 ± 0.26 |
| EmS | 1.74 ± 0.06 s | 1.62 ± 0.06 | 1.87 ± 0.13 | 1.78 ± 0.05 | |
| Sex main effect: F (1, 88) = 16.510, p < 0.001 Sex × stress interaction: F (1, 88) = 4.63, p = 0.034 | |||||
| HIAA/5-HT | FS | 0.57 ± 0.04 | 0.49 ± 0.02 | 0.69 ± 0.05 | 0.60 ± 0.05 |
| EmS | 0.62 ± 0.02 | 0.51 ± 0.02 | 0.67 ± 0.03 | 0.61 ± 0.01 | |
| Sex main effect: F (1, 88) = 16.759, p < 0.001; Rearing main effect: F (1, 88) = 14.452, p < 0.001 | |||||
| Striatum | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | FS | 4.07 ± 0.20 | 3.53 ± 0.18 | 3.66 ± 0.17 | 4.15 ± 0.25 |
| EmS | 3.82 ± 0.18 | 3.61 ± 0.20 | 3.54 ± 0.14 | 4.44 ± 0.29 | |
| Sex × Rearing interaction: F (1, 88) = 13.859, p < 0.001 | |||||
| HIAA | FS | 2.99 ± 0.16 | 2.46 ± 0.14 | 3.08 ± 0.10 | 2.82 ± 0.15 |
| EmS | 3.07 ± 0.17 | 2.59 ± 0.16 | 2.86 ± 0.15 | 3.02 ± 0.19 | |
| Rearing main effect: F (1, 88) = 6.273, p = 0.014; Sex × Rearing interaction: F (1, 88) = 4.259, p = 0.042 | |||||
| HIAA/5-HT | FS | 0.74 ± 0.03 | 0.70 ± 0.03 | 0.86 ± 0.04 | 0.69 ± 0.02 |
| EmS | 0.80 ± 0.03 | 0.72 ± 0.02 | 0.82 ± 0.04 | 0.69 ± 0.02 | |
| Rearing main effect: F (1, 88) = 24.97, p < 0.001; Sex × Rearing interaction: F (1, 88) = 3.967, p = 0.049 | |||||
| Hypothalamus | |||||
| Control, m | MSI, m | Control, f | MSI, f | ||
| 5-HT | FS | 6.13 ± 0.30 | 5.59 ± 0.20 | 5.62 ± 0.23 | 6.19 ± 0.18 |
| EmS | 6.81 ± 0.61 | 5.88 ± 0.24 | 5.16 ± 0.28 | 6.02 ± 0.16 | |
| Sex × Rearing interaction: F (1, 88) = 10.115, p = 0.002; Sex × stress interaction: F (1, 88) = 3.110, p = 0.081 | |||||
| HIAA | FS | 2.83 ± 0.13 | 2.50 ± 0.11 | 3.14 ± 0.10 | 3.05 ± 0.08 |
| EmS | 3.35 ± 0.25 | 2.75 ± 0.13 s | 2.92 ± 0.15 | 3.11 ± 0.09 | |
| Rearing main effect: F (1, 88) = 3.889, p = 0.052; Sex × Rearing interaction: F (1, 88) = 6.165, p = 0.015; Sex × stress interaction: F (1, 88) = 5.110, p = 0.026 | |||||
| HIAA/5-HT | FS | 0.47 ± 0,02 | 0.45 ± 0.01 | 0.57 ± 0.02 | 0.50 ± 0.02 |
| EmS | 0.50 ± 0.01 | 0.47 ± 0.01 | 0.57 ± 0.01 | 0.52 ± 0.02 | |
| Sex main effect: F (1, 88) = 35.432, p < 0.001; Rearing main effect: F (1, 88) = 13.028, p < 0.001; Stress main effect: F (1, 88) = 2.978, p = 0.088 | |||||
| ma | NE | DA | DOPAC | HVA | DOPAC/DA | HVA/ DA | 5-HT | HIAA | HIAA/ 5-HT | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sex | m | f | m | f | m | f | m | f | m | f | m | f | m | f | m | f | m | f |
| 1 month | ||||||||||||||||||
| FC | = | = | = | = | = | = | ↓ | ↓ * | = | = | ↓ | ↓ * | = | = | = | = | = | = |
| Hip | ↑ * | ↑ | = | = | = | = | nd | nd | = | = | = | = | ↑ | ↑ | = | = | = | = |
| Str | = | = | = | = | ↓ * | = | ↓ * | = | = | = | ↓ | ↓ | = | = | = | = | = | = |
| Hyp | ↑ * | = | ↓ * | = | = | = | = | = | ↑ | ↑ * | ↑ | ↑ | ↓ * | ↓ | ↓ * | = | = | = |
| 2 months | ||||||||||||||||||
| FC | = | = | = | = | = | = | = | = | = | = | = | = | ↑ * | ↓ * | ↑ * | ↓ * | = | = |
| Hip | ↓ * | ↓ | = | = | = | = | = | = | = | = | = | = | = | = | = | = | = | = |
| Str | = | = | ↓ | ↓ * | ↓ * | = | ↑ | ↑ * | ↓ * | = | = | = | ↑ | ↑ * | = | = | ||
| Hyp | = | ↓ * | = | = | ↓ | ↓ | ↓ * | ↓ * | = | ↓ * | ↓ * | ↓ * | ↓ * | = | ↓ * | = | = | ↓ * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavrenkova, P.V.; Khlebnikova, N.N.; Alchinova, I.B.; Demorzhi, M.S.; Shoibonov, B.B.; Karganov, M.Y. Effects of Maternal Separation and Subsequent Stress on Behaviors and Brain Monoamines in Rats. Brain Sci. 2023, 13, 956. https://doi.org/10.3390/brainsci13060956
Mavrenkova PV, Khlebnikova NN, Alchinova IB, Demorzhi MS, Shoibonov BB, Karganov MY. Effects of Maternal Separation and Subsequent Stress on Behaviors and Brain Monoamines in Rats. Brain Sciences. 2023; 13(6):956. https://doi.org/10.3390/brainsci13060956
Chicago/Turabian StyleMavrenkova, Polina V., Nadezhda N. Khlebnikova, Irina B. Alchinova, Marina S. Demorzhi, Batozhab B. Shoibonov, and Mikhail Yu. Karganov. 2023. "Effects of Maternal Separation and Subsequent Stress on Behaviors and Brain Monoamines in Rats" Brain Sciences 13, no. 6: 956. https://doi.org/10.3390/brainsci13060956
APA StyleMavrenkova, P. V., Khlebnikova, N. N., Alchinova, I. B., Demorzhi, M. S., Shoibonov, B. B., & Karganov, M. Y. (2023). Effects of Maternal Separation and Subsequent Stress on Behaviors and Brain Monoamines in Rats. Brain Sciences, 13(6), 956. https://doi.org/10.3390/brainsci13060956