

Quantification of Non-Motor Symptoms in Parkinsonian Cynomolgus Monkeys
 and
and
Abstract
:1. Background
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Behavioral Tests
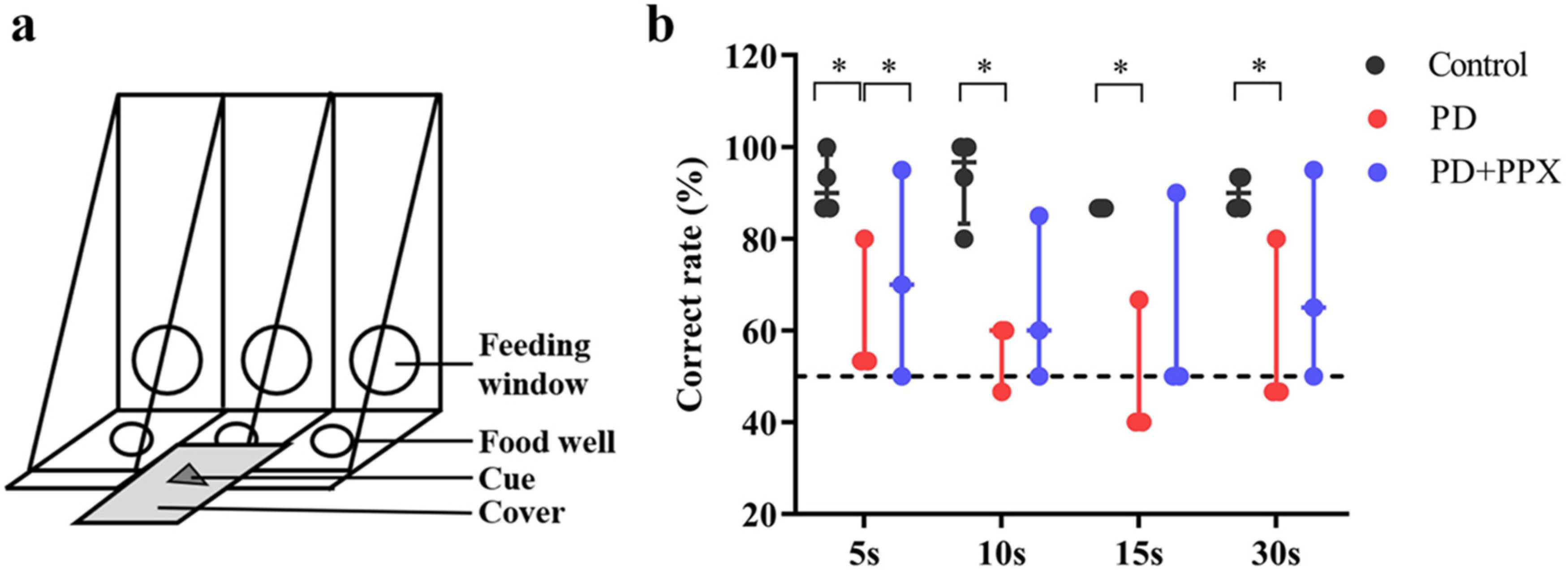
2.3.1. Delayed Matching-To-Sample Task
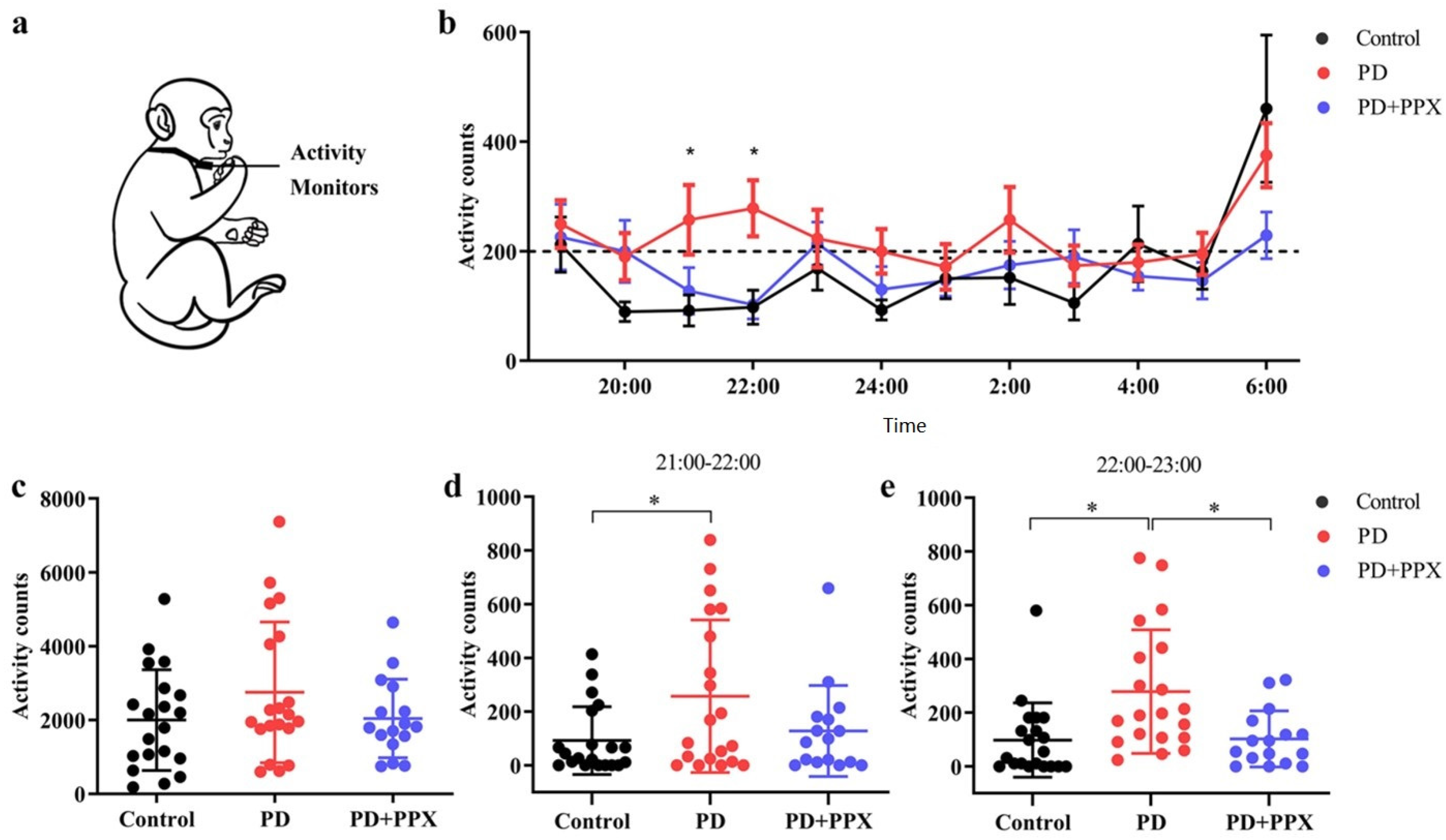
2.3.2. Physical Activity Monitor
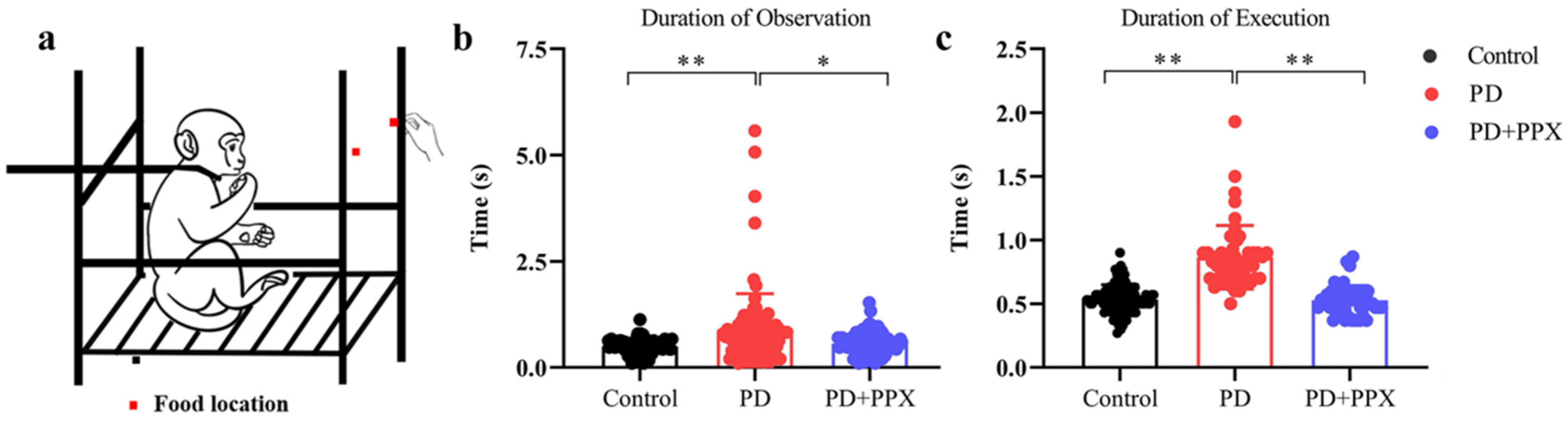
2.3.3. Apathy Feeding Task
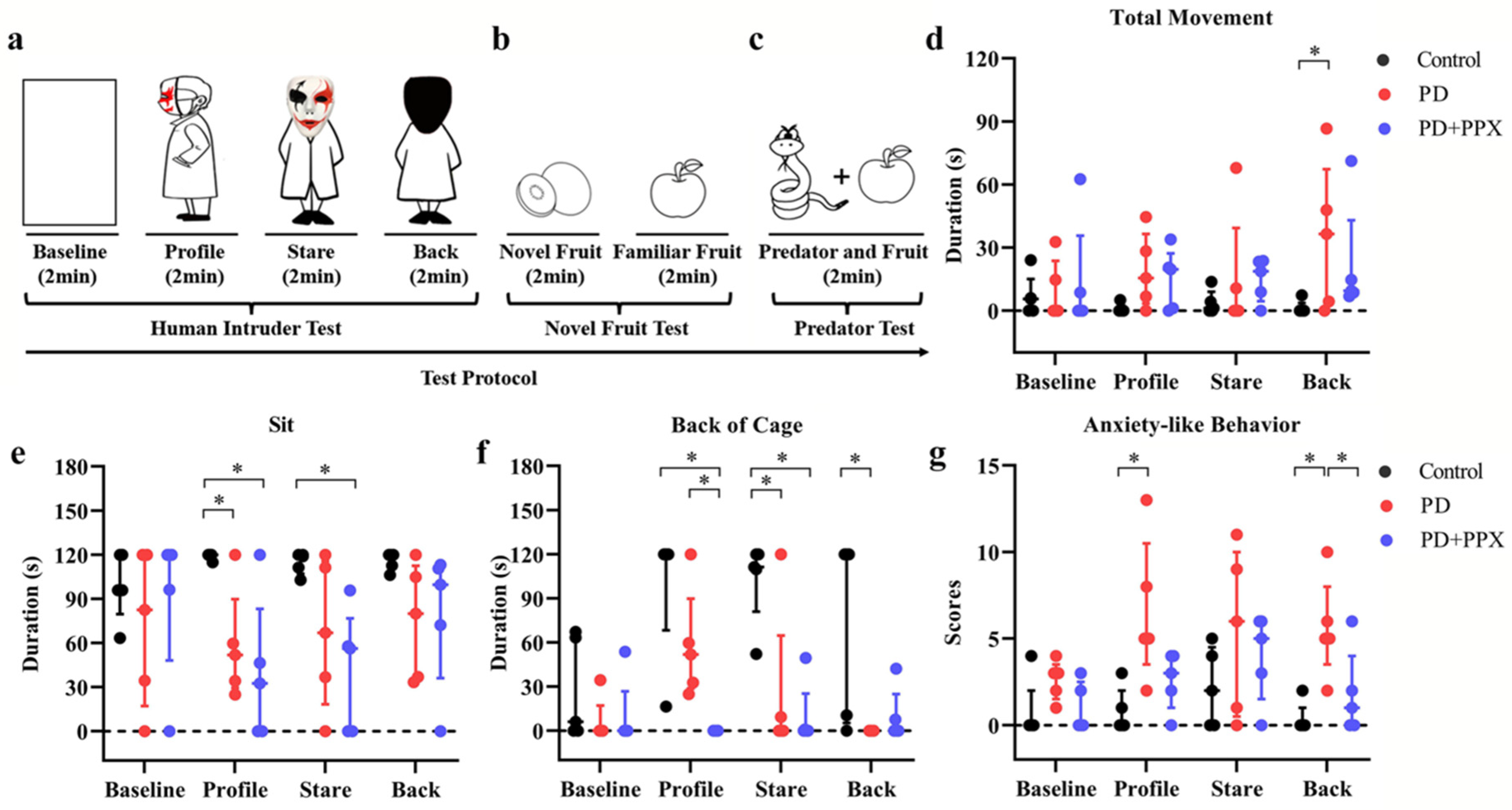
2.3.4. Behavioral Tests for Anxiety
Human Intruder Test
Novel Fruit Test
Predator Confrontation Test
2.4. Statistical Analysis
3. Results
3.1. PD Monkeys Showed Working Memory Deficits in DMTS Task
3.2. PD Monkeys Showed More Nocturnal Activity
3.3. PD Monkeys Showed Deficits in AFT Task
3.4. PPX Reduced Anxiety-Like Behaviors in PD Monkeys
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of Parkinson’s disease: Risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [Google Scholar] [CrossRef]
- Chia, S.J.; Tan, E.K.; Chao, Y.X. Historical Perspective: Models of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 2464. [Google Scholar] [CrossRef] [Green Version]
- Prediger, R.D.; Matheus, F.C.; Schwarzbold, M.L.; Lima, M.M.; Vital, M.A. Anxiety in Parkinson’s disease: A critical review of experimental and clinical studies. Neuropharmacology 2012, 62, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-motor features of Parkinson disease. Nature reviews. Neuroscience 2017, 18, 435–450. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 Parkinson’s Disease Collaborators. Global, regional, and national burden of Parkinson’s disease, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef] [Green Version]
- van Wamelen, D.J.; Martinez-Martin, P.; Weintraub, D.; Schrag, A.; Antonini, A.; Falup-Pecurariu, C.; Odin, P.; Ray Chaudhuri, K.; International, P.; Movement Disorder Society Parkinson’s Disease Non-Motor Study Group. The Non-Motor Symptoms Scale in Parkinson’s disease: Validation and use. Acta Neurol. Scand. 2021, 143, 3–12. [Google Scholar] [CrossRef]
- Hely, M.A.; Morris, J.G.; Reid, W.G.; Trafficante, R. Sydney Multicenter Study of Parkinson’s disease: Non-L-dopa-responsive problems dominate at 15 years. Mov. Disord. Off. J. Mov. Disord. Soc. 2005, 20, 190–199. [Google Scholar] [CrossRef]
- Villalba, R.M.; Smith, Y. Loss and remodeling of striatal dendritic spines in Parkinson’s disease: From homeostasis to maladaptive plasticity? J. Neural. Transm. 2018, 125, 431–447. [Google Scholar] [CrossRef] [PubMed]
- Vezoli, J.; Fifel, K.; Leviel, V.; Dehay, C.; Kennedy, H.; Cooper, H.M.; Gronfier, C.; Procyk, E. Early presymptomatic and long-term changes of rest activity cycles and cognitive behavior in a MPTP-monkey model of Parkinson’s disease. PLoS ONE 2011, 6, e23952. [Google Scholar] [CrossRef] [Green Version]
- Han, N.R.; Kim, Y.K.; Ahn, S.; Hwang, T.Y.; Lee, H.; Park, H.J. A Comprehensive Phenotype of Non-motor Impairments and Distribution of Alpha-Synuclein Deposition in Parkinsonism-Induced Mice by a Combination Injection of MPTP and Probenecid. Front. Aging Neurosci. 2020, 12, 599045. [Google Scholar] [CrossRef] [PubMed]
- Tardif, S.D.; Coleman, K.; Hobbs, T.R.; Lutz, C. IACUC review of nonhuman primate research. ILAR J. 2013, 54, 234–245. [Google Scholar] [CrossRef] [Green Version]
- Przedborski, S.; Jackson-Lewis, V.; Yokoyama, R.; Shibata, T.; Dawson, V.L.; Dawson, T.M. Role of neuronal nitric oxide in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced dopaminergic neurotoxicity. Proc. Natl. Acad. Sci. USA 1996, 93, 4565–4571. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Sun, Y.Q.; Teoh, H.C. Comparison of nocturnal symptoms in advanced Parkinson’s disease patients with sleep disturbances: Pramipexole sustained release versus immediate release formulations. Drug Des. Dev. Ther. 2018, 12, 2017–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, T.; Ye, R.; Zhang, B. Efficacy and safety of pramipexole extended-release in Parkinson’s disease: A review based on meta-analysis of randomized controlled trials. Eur. J. Neurol. 2017, 24, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, F.A.; Egner, T.; Buckley, M.J. Monitoring Demands for Executive Control: Shared Functions between Human and Nonhuman Primates. Trends Neurosci. 2017, 40, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonini, A.; Barone, P.; Ceravolo, R.; Fabbrini, G.; Tinazzi, M.; Abbruzzese, G. Role of pramipexole in the management of Parkinson’s disease. CNS Drugs 2010, 24, 829–841. [Google Scholar] [CrossRef]
- Sgambato, V.; Tremblay, L. Pathophysiology of dyskinesia and behavioral disorders in non-human primates: The role of serotonergic fibers. Neural Transm 2018, 125, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.Y.; Park, H.J.; Ahn, Y.H.; Lee, P.H. Neuroprotective effect of L-dopa on dopaminergic neurons is comparable to pramipexol in MPTP-treated animal model of Parkinson’s disease: A direct comparison study. J. Neurochem. 2009, 111, 1042–1050. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, X.; Zhang, P.; Ma, Y.; Wang, L.; Xu, H.; Sui, D. Neuroprotective effects of pramipexole transdermal patch in the MPTP-induced mouse model of Parkinson’s disease. J. Pharmacol. Sci. 2018, 138, 31–37. [Google Scholar] [CrossRef]
- Patterson, T.A.; Li, M.; Hotchkiss, C.E.; Mauz, A.; Eddie, M.; Greischel, A.; Stierstorfer, B.; Deschl, U.; Paule, M.G. Toxicity assessment of pramipexole in juvenile rhesus monkeys. Toxicology 2010, 276, 164–171. [Google Scholar] [CrossRef]
- Yue, F.; Chan, P.; Wan, H.; Weeks, R.; Grondin, R.; Than, Z. Similarities and Differences in Cognitive Deficits and Responsiveness to L-Dopa Between Aged and MPTP-Treated Cynomolgus Monkeys Tested on the Same Tasks. Cell Transpl. 2014, 23, 790. [Google Scholar]
- Brown, C.A.; Campbell, M.C.; Karimi, M.; Tabbal, S.D.; Loftin, S.K.; Tian, L.L.; Moerlein, S.M.; Perlmutter, J.S. Dopamine pathway loss in nucleus accumbens and ventral tegmental area predicts apathetic behavior in MPTP-lesioned monkeys. Exp. Neurol. 2012, 236, 190–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalin, N.H.; Shelton, S.E. Defensive behaviors in infant rhesus monkeys: Environmental cues and neurochemical regulation. Science 1989, 243, 1718–1721. [Google Scholar] [CrossRef] [PubMed]
- Hamel, A.; Lutz, C.; Coleman, K.; Worlein, J.; Peterson, E.; Rosenberg, K.; Novak, M.; Meyer, J. Responses to the Human Intruder Test are related to hair cortisol phenotype and sex in rhesus macaques (Macaca mulatta). Am. J. Primatol. 2017, 79, e22526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celano, C.M.; Daunis, D.J.; Lokko, H.N.; Campbell, K.A.; Huffman, J.C. Anxiety Disorders and Cardiovascular Disease. Curr. Psychiatry Rep. 2016, 18, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maestripieri, D. Gestural communication in three species of macaques (Macaca mulatta, M. nemestrina, M. arctoides). In Gestural Communication in Nonhuman and Human Primates; John Benjamins Publishing Company: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Machado, C.J.; Kazama, A.M.; Bachevalier, J. Impact of amygdala, orbital frontal, or hippocampal lesions on threat avoidance and emotional reactivity in nonhuman primates. Emotion 2009, 9, 147–163. [Google Scholar] [CrossRef] [Green Version]
- Williamson, D.E.; Coleman, K.; Bacanu, S.A.; Devlin, B.J.; Rogers, J.; Ryan, N.D.; Cameron, J.L. Heritability of fearful-anxious endophenotypes in infant rhesus macaques: A preliminary report. Biol. Psychiatry 2003, 53, 284–291. [Google Scholar] [CrossRef]
- Barros, M.; Tomaz, C. Non-human primate models for investigating fear and anxiety. Neurosci. Biobehav. Rev. 2002, 26, 187–201. [Google Scholar] [CrossRef]
- Boulet, S.; Mounayar, S.; Poupard, A.; Bertrand, A.; Jan, C.; Pessiglione, M.; Hirsch, E.C.; Feuerstein, C.; Francois, C.; Feger, J.; et al. Behavioral recovery in MPTP-treated monkeys: Neurochemical mechanisms studied by intrastriatal microdialysis. J. Neurosci. 2008, 28, 9575–9584. [Google Scholar] [CrossRef] [Green Version]
- Barraud, Q.; Lambrecq, V.; Forni, C.; McGuire, S.; Hill, M.; Bioulac, B.; Balzamo, E.; Bezard, E.; Tison, F.; Ghorayeb, I. Sleep disorders in Parkinson’s disease: The contribution of the MPTP non-human primate model. Exp. Neurol. 2009, 219, 574–582. [Google Scholar] [CrossRef]
- Delamarre, A.; MacSweeney, C.; Suzuki, R.; Brown, A.J.; Li, Q.; Pioli, E.Y.; Bezard, E. Gastrointestinal and metabolic function in the MPTP-treated macaque model of Parkinson’s disease. Heliyon 2020, 6, e05771. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal death and implications for therapeutic intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madetko, N.; Migda, B.; Alster, P.; Turski, P.; Koziorowski, D.; Friedman, A. Platelet-to-lymphocyte ratio and neutrophil-tolymphocyte ratio may reflect differences in PD and MSA-P neuroinflammation patterns. Neurol. Neurochir. Pol. 2022, 56, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- McGeer, P.L.; McGeer, E.G. History of innate immunity in neurodegenerative disorders. Front. Pharmacol. 2011, 2, 77. [Google Scholar] [CrossRef] [Green Version]
- McGeer, P.L.; Schwab, C.; Parent, A.; Doudet, D. Presence of reactive microglia in monkey substantia nigra years after 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine administration. Ann. Neurol. 2003, 54, 599–604. [Google Scholar] [CrossRef]
- Schneider, J.S. Chronic exposure to low doses of MPTP. II. Neurochemical and pathological consequences in cognitively-impaired, motor asymptomatic monkeys. Brain Res. 1990, 534, 25–36. [Google Scholar] [CrossRef]
- Schneider, J.S.; Kovelowski, C.J., 2nd. Chronic exposure to low doses of MPTP. I. Cognitive deficits in motor asymptomatic monkeys. Brain Res. 1990, 519, 122–128. [Google Scholar] [CrossRef]
- Fernandez-Ruiz, J.; Doudet, D.J.; Aigner, T.G. Long-term cognitive impairment in MPTP-treated rhesus monkeys. Neuroreport 1995, 7, 102–104. [Google Scholar] [CrossRef]
- Konnova, E.A.; Swanberg, M. Animal Models of Parkinson’s Disease. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; Stoker, T.B., Greenland, J.C., Eds.; Exon Publications: Brisbane, AU, Australia, 2018. [Google Scholar]
- Vuckovic, M.G.; Wood, R.I.; Holschneider, D.P.; Abernathy, A.; Togasaki, D.M.; Smith, A.; Petzinger, G.M.; Jakowec, M.W. Memory, mood, dopamine, and serotonin in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-lesioned mouse model of basal ganglia injury. Neurobiol. Dis. 2008, 32, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Titova, N.; Schapira, A.H.V.; Chaudhuri, K.R.; Qamar, M.A.; Katunina, E.; Jenner, P. Nonmotor Symptoms in Experimental Models of Parkinson’s Disease. Int. Rev. Neurobiol. 2017, 133, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Visanji, N.P.; Gomez-Ramirez, J.; Johnston, T.H.; Pires, D.; Voon, V.; Brotchie, J.M.; Fox, S.H. Pharmacological characterization of psychosis-like behavior in the MPTP-lesioned nonhuman primate model of Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2006, 21, 1879–1891. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.A.; Ross, C.N.; Spross, J.; Cheng, C.J.; Izquierdo, A.; Biju, K.C.; Chen, C.; Li, S.; Tardif, S.D. Behavioral phenotypes associated with MPTP induction of partial lesions in common marmosets (Callithrix jacchus). Behav. Brain Res. 2017, 325, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, C.; Souaiby, L.; Advenier-Iakovlev, E.; Gaillard, R. Pramipexole and Electroconvulsive Therapy in Treatment-Resistant Depression. Clin. Neuropharmacol. 2017, 40, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Iravani, M.M.; Haddon, C.O.; Cooper, J.M.; Jenner, P.; Schapira, A.H. Pramipexole protects against MPTP toxicity in non-human primates. J. Neurochem. 2006, 96, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Silindir, M.; Ozer, A.Y. The benefits of pramipexole selection in the treatment of Parkinson’s disease. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2014, 35, 1505–1511. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Evaluation Indicator |
|---|---|
| Grind teeth | Less than 30 s, for 1 point 30–60 s, for 2 points 60–90 s, for 3 points 90 s or more, for 4 points |
| Open mouth | Less than 10 s, for 1 point 10–30 s, for 2 points 30–60 s, for 3 points 60 s or more, for 4 points |
| Tactile/oral exploration, scratch, freeze | Less than 2 s, for 1 point 2–5 s, for 2 points 5–10 s, for 3 points 10 s or more, for 4 points |
| Yawn, pounce, over one’s eyes, turn around, fear grimace, cage shake | 1 time, for 1 point 2 times, for 2 points 3 times, for 3 points 4 times or more, for 4 points |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Y.; Gan, C.; Chen, Z.; Qi, Z.; Meng, Z.; Yue, F. Quantification of Non-Motor Symptoms in Parkinsonian Cynomolgus Monkeys. Brain Sci. 2023, 13, 1153. https://doi.org/10.3390/brainsci13081153
Bao Y, Gan C, Chen Z, Qi Z, Meng Z, Yue F. Quantification of Non-Motor Symptoms in Parkinsonian Cynomolgus Monkeys. Brain Sciences. 2023; 13(8):1153. https://doi.org/10.3390/brainsci13081153
Chicago/Turabian StyleBao, Yu, Chaoning Gan, Zuyue Chen, Zhongquan Qi, Zhiqiang Meng, and Feng Yue. 2023. "Quantification of Non-Motor Symptoms in Parkinsonian Cynomolgus Monkeys" Brain Sciences 13, no. 8: 1153. https://doi.org/10.3390/brainsci13081153