
Altered Estrous Cyclicity and Feeding Neurocircuitry, but Not Cardiovascular Indices in Female Offspring from Dams with Previous Vertical Sleeve Gastrectomy Surgery

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Assurance
2.2. Animals
2.2.1. Surgical Procedures
2.2.2. Post-Operative Care
2.2.3. Blood Pressure (BP) Determination
2.2.4. Vaginal Lavage and Determination of Estrus Cyclicity
2.2.5. Novel Diet-Induction Test
2.2.6. Immunohistochemistry
2.2.7. Microscopy and ImageJ Cell Counting
2.2.8. Fresh Tissue Harvesting
2.2.9. Triglyceride Assay
2.2.10. RNA Processing and Real-Time PCR
2.2.11. Statistics
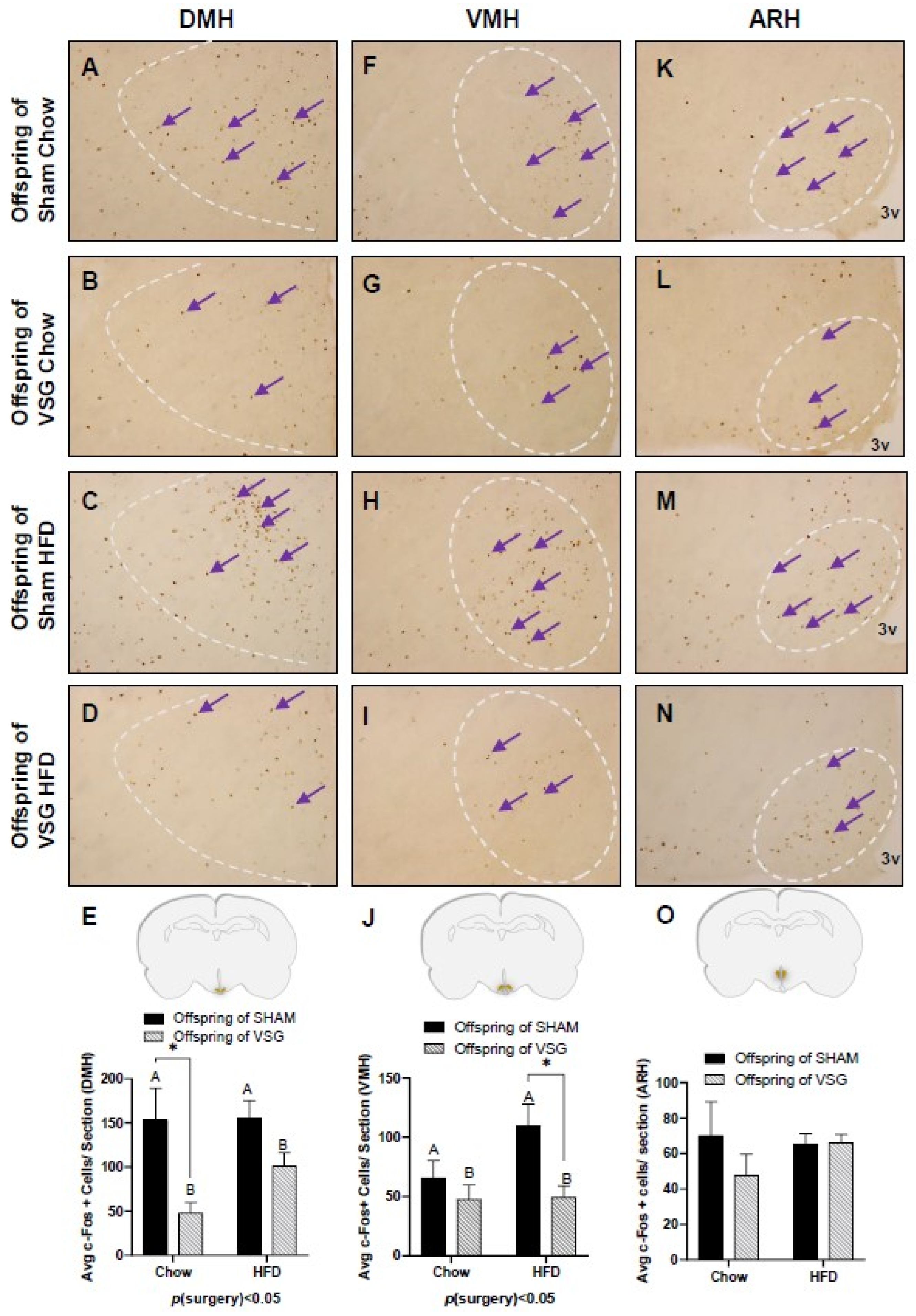
3. Results
4. Discussion
5. Caveats and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Stroh, C.; Weiner, R.; Wolff, S.; Knoll, C.; Manger, T. Influences of gender on complication rate and outcome after Roux-en-Y gastric bypass: Data analysis of more than 10,000 operations from the German Bariatric Surgery Registry. Obes. Surg. 2014, 24, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Courcoulas, A.P.; Christian, N.J.; Belle, S.H.; Berk, P.D.; Flum, D.R.; Garcia, L.; Horlick, M.; Kalarchian, M.A.; King, W.C.; Mitchell, J.E.; et al. Weight change and health outcomes at 3 years after bariatric surgery among individuals with severe obesity. JAMA 2013, 310, 2416–2425. [Google Scholar] [CrossRef] [PubMed]
- Casella, G.; Soricelli, E.; Castagneto-Gissey, L.; Redler, A.; Basso, N.; Mingrone, G. Changes in insulin sensitivity and secretion after sleeve gastrectomy. Br. J. Surg. 2015, 103, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Coleman, K.J.; Huang, Y.C.; Hendee, F.; Watson, H.L.; Casillas, R.A.; Brookey, J. Three-year weight outcomes from a bariatric surgery registry in a large integrated healthcare system. Surg. Obes. Relat. Dis. Off. J. Am. Soc. Bariatr. Surg. 2014, 10, 396–403. [Google Scholar] [CrossRef]
- Buchwald, H.; Estok, R.; Fahrbach, K.; Banel, D.; Jensen, M.D.; Pories, W.J.; Bantle, J.P.; Sledge, I. Weight and type 2 diabetes after bariatric surgery: Systematic review and meta-analysis. Am. J. Med. 2009, 122, 248–256.e5. [Google Scholar] [CrossRef]
- Teitelman, M.; Grotegut, C.; Williams, N.; Lewis, J. The Impact of Bariatric Surgery on Menstrual Patterns. Obes. Surg. 2006, 16, 1457–1463. [Google Scholar] [CrossRef]
- Sarwer, D.B.; Spitzer, J.C.; Wadden, T.A.; Mitchell, J.E.; Lancaster, K.; Courcoulas, A.; Gourash, W.; Rosen, R.C.; Christian, N.J. Changes in sexual functioning and sex hormone levels in women following bariatric surgery. JAMA Surg. 2014, 149, 26–33. [Google Scholar] [CrossRef]
- Rochester, D.; Jain, A.; Polotsky, A.J.; Polotsky, H.; Gibbs, K.; Isaac, B.; Zeitlian, G.; Hickmon, C.; Feng, S.; Santoro, N. Partial recovery of luteal function after bariatric surgery in obese women. Fertil. Steril. 2009, 92, 1410–1415. [Google Scholar] [CrossRef]
- Tan, O.; Carr, B.R. The impact of bariatric surgery on obesity-related infertility and in vitro fertilization outcomes. Semin. Reprod. Med. 2012, 30, 517–528. [Google Scholar] [CrossRef]
- Johansson, K.; Cnattingius, S.; Naslund, I.; Roos, N.; Trolle Lagerros, Y.; Granath, F.; Stephansson, O.; Neovius, M. Outcomes of pregnancy after bariatric surgery. N. Engl. J. Med. 2015, 372, 814–824. [Google Scholar] [CrossRef]
- Galazis, N.; Docheva, N.; Simillis, C.; Nicolaides, K.H. Maternal and neonatal outcomes in women undergoing bariatric surgery: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 181, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, A.Y.; Levy, A.; Levi, I.; Mazor, M.; Wiznitzer, A.; Sheiner, E. Effect of bariatric surgery on pregnancy outcome. Int. J. Gynaecol. Obstet. Off. Organ. Int. Fed. Gynaecol. Obstet. 2008, 103, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Cianflone, K.; Biron, S.; Hould, F.S.; Lebel, S.; Marceau, S.; Lescelleur, O.; Biertho, L.; Simard, S.; Kral, J.G.; et al. Effects of Bariatric Surgical Weight Loss in Mothers on Intergenerational Transmission of Obesity. Endocrinology 2009, 150, 4816. [Google Scholar] [CrossRef]
- Sheiner, E.; Levy, A.; Silverberg, D.; Menes, T.S.; Levy, I.; Katz, M.; Mazor, M. Pregnancy after bariatric surgery is not associated with adverse perinatal outcome. Am. J. Obs. Gynecol. 2004, 190, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Rottenstreich, A.; Elchalal, U.; Kleinstern, G.; Beglaibter, N.; Khalaileh, A.; Elazary, R. Maternal and Perinatal Outcomes After Laparoscopic Sleeve Gastrectomy. Obstet. Gynecol. 2018, 131, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Chevrot, A.; Kayem, G.; Coupaye, M.; Lesage, N.; Msika, S.; Mandelbrot, L. Impact of bariatric surgery on fetal growth restriction: Experience of a perinatal and bariatric surgery center. Am. J. Obs. Gynecol. 2016, 214, 655.e1–655.e7. [Google Scholar] [CrossRef]
- Akhter, Z.; Rankin, J.; Ceulemans, D.; Ngongalah, L.; Ackroyd, R.; Devlieger, R.; Vieira, R.; Heslehurst, N. Pregnancy after bariatric surgery and adverse perinatal outcomes: A systematic review and meta-analysis. PLoS Med. 2019, 16, e1002866. [Google Scholar] [CrossRef]
- Wei, J.-N.; Sung, F.-C.; Li, C.-Y.; Chang, C.-H.; Lin, R.-S.; Lin, C.-C.; Chiang, C.-C.; Chuang, L.-M. Low Birth Weight and High Birth Weight Infants Are Both at an Increased Risk to Have Type 2 Diabetes Among Schoolchildren in Taiwan. Diabetes Care 2003, 26, 343–348. [Google Scholar] [CrossRef]
- Pettitt, D.; Jovanovic, L. Birth weight as a predictor of type 2 diabetes mellitus: The U-shaped curve. Curr. Diabetes Rep. 2001, 1, 78–81. [Google Scholar] [CrossRef]
- Huang, R.C.; Burke, V.; Newnham, J.P.; Stanley, F.J.; Kendall, G.E.; Landau, L.I.; Oddy, W.H.; Blake, K.V.; Palmer, L.J.; Beilin, L.J. Perinatal and childhood origins of cardiovascular disease. Int. J. Obes. 2006, 31, 236–244. [Google Scholar] [CrossRef]
- Lawson, W.J.; Shirey, K.; Spann, R.A.; Zamarripa, C.A.; Hosler, J.P.; Grayson, B.E. Vertical sleeve gastrectomy improves indices of metabolic disease in rodent model of surgical menopause. Menopause 2017, 24, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Spann, R.A.; Lawson, W.J.; Bidwell, G.L.; Zamarripa, C.A.; Maranon, R.O.; Bandyopadhyay, S.; Taylor, E.R.; Reckelhoff, J.F.; Garrett, M.R.; Grayson, B.E. Rodent Vertical Sleeve Gastrectomy Alters Maternal Immune Health and Feto-placental Development. Clin. Sci. 2017, 132, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Grayson, B.E.; Schneider, K.M.; Woods, S.C.; Seeley, R.J. Improved rodent maternal metabolism but reduced intrauterine growth after vertical sleeve gastrectomy. Sci. Transl. Med. 2013, 5, 199ra112. [Google Scholar] [CrossRef] [PubMed]
- Deer, E.M.; Welch, B.; Hernandez, L.L.; Seeley, R.J.; Grayson, B.E. Nutrient and hormone composition of milk is altered in rodent dams post-bariatric surgery. J. Dev. Orig. Health Dis. 2020, 11, 71–77. [Google Scholar] [CrossRef]
- Spann, R.A.; Welch, B.A.; Grayson, B.E. Ghrelin signalling is dysregulated in male but not female offspring in a rat model of maternal vertical sleeve gastrectomy. J. Neuroendocr. 2021, 33, e12913. [Google Scholar] [CrossRef]
- Deer, E.M.; Phillips, C.L.; Welch, B.A.; Himel, A.R.; Duncan, B.C.; Spann, R.A.; Grayson, B.E. Dysregulated appetitive leptin signaling in male rodent offspring from post-bariatric dams. Curr. Res. Physiol. 2020, 3, 50–58. [Google Scholar] [CrossRef]
- Spann, R.A.; Taylor, E.B.; Welch, B.A.; Grayson, B.E. Altered immune system in offspring of rat maternal vertical sleeve gastrectomy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R852–R863. [Google Scholar] [CrossRef]
- Himel, A.R.; Taylor, E.B.; Phillips, C.L.; Welch, B.A.; Spann, R.A.; Bandyopadhyay, S.; Grayson, B.E. Splenectomy fails to attenuate immuno-hematologic changes after rodent vertical sleeve gastrectomy. Exp. Biol. Med. 2019, 244, 1125–1135. [Google Scholar] [CrossRef]
- Whitaker, K.L.; Jarvis, M.J.; Beeken, R.J.; Boniface, D.; Wardle, J. Comparing maternal and paternal intergenerational transmission of obesity risk in a large population-based sample. Am. J. Clin. Nutr. 2010, 91, 1560–1567. [Google Scholar] [CrossRef]
- Roseboom, T.J.; van der Meulen, J.H.P.; Ravelli, A.C.J.; Osmond, C.; Barker, D.J.P.; Bleker, O.P. Effects of Prenatal Exposure to the Dutch Famine on Adult Disease in Later Life: An Overview. Twin Res. 2001, 4, 293–298. [Google Scholar] [CrossRef]
- Moxthe, L.C.; Sauls, R.; Ruiz, M.; Stern, M.; Gonzalvo, J.; Gray, H.L. Effects of Bariatric Surgeries on Male and Female Fertility: A Systematic Review. J. Reprod. Infertil. 2020, 21, 71–86. [Google Scholar] [PubMed]
- Shah, D.K.; Ginsburg, E.S. Bariatric surgery and fertility. Curr. Opin. Obstet. Gynecol. 2010, 22, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Spann, R.A.; Grayson, B.E. Curbing Obesity from One Generation to Another: The Effects of Bariatric Surgery on the In Utero Environment and Beyond. Reprod. Sci. 2020, 27, 1821–1833. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measure | Offspring of Sham-Chow | Offspring of VSG-Chow | Offspring of Sham-HFD | Offspring of VSG-HFD | Statistics | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | ± | SEM | Mean | ± | SEM | Mean | ± | SEM | Mean | ± | SEM | 2W ANOVA | |
| Body Weight (g) | 335.60 | ± | 5.74 | 314.70 | ± | 10.32 | 328.60 | ± | 11.31 | 318.90 | ± | 30.11 | NS |
| Body Fat (g) | 81.36 | ± | 6.18 | 75.76 | ± | 11.36 | 80.06 | ± | 10.22 | 87.58 | ± | 22.31 | NS |
| Fat Mass Percent (%) | 24.04 | ± | 1.52 | 23.55 | ± | 2.99 | 23.91 | ± | 2.59 | 26.41 | ± | 4.75 | NS |
| Lean Mass (g) | 176.70 | ± | 2.51 | 168.50 | ± | 5.47 | 169.50 | ± | 4.78 | 160.20 | ± | 13.69 | NS |
| Lean Mass Percent (%) | 52.86 | ± | 1.51 | 54.02 | ± | 2.69 | 51.82 | ± | 1.76 | 50.42 | ± | 1.03 | NS |
| Gene Name | Sham-Chow | VSG-Chow | Sham-HFD | VSG-HFD | Statistics | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | ± | SEM | Mean | ± | SEM | Mean | ± | SEM | Mean | ± | SEM | 2W-ANOVA | |
| Agouti-related peptide (AGRP) | 1.00 | ± | 0.15 | 0.99 | ± | 0.12 | 0.48 | ± | 0.18 | 1.73 | ± | 0.42 | ** p(surgery) < 0.01 |
| Glucocorticoid Receptor (NR3C1) | 1.00 | ± | 0.16 | 1.16 | ± | 0.10 | 1.13 | ± | 0.27 | 0.72 | ± | 0.21 | * p(diet) < 0.05 |
| Glucose Transporter 1 (GLUT-1) | 1.00 | ± | 0.16 | 1.07 | ± | 0.06 | 0.97 | ± | 0.30 | 0.51 | ± | 0.16 | NS |
| Glucose Transporter 2 (GLUT-2) | 1.00 | ± | 0.16 | 1.16 | ± | 0.10 | 1.13 | ± | 0.27 | 0.72 | ± | 0.21 | NS |
| Glucose Transporter 4 (GLUT-4) | 1.00 | ± | 0.09 | 0.97 | ± | 0.07 | 0.93 | ± | 0.14 | 0.75 | ± | 0.19 | NS |
| Insulin Growth Factor 2 Receptor (IGF2R) | 1.00 | ± | 0.14 | 1.01 | ± | 0.06 | 0.88 | ± | 0.14 | 0.50 | ± | 0.14 | * p(diet) < 0.05 |
| Interleukin 6 (IL-6) | 1.00 | ± | 0.08 | 1.17 | ± | 0.13 | 0.62 | ± | 0.17 | 1.30 | ± | 0.14 | ** p(surgery) < 0.01 |
| Neuropeptide Y (NPY) | 1.00 | ± | 0.15 | 0.96 | ± | 0.11 | 0.42 | ± | 0.13 | 1.31 | ± | 0.26 | * p(surgery) < 0.05 |
| Pro-opiomelanacortin (POMC) | 1.00 | ± | 0.23 | 0.98 | ± | 0.22 | 0.31 | ± | 0.16 | 0.92 | ± | 0.23 | NS |
| Very long-chain acyl-CoA synthetase (SLC27A) | 1.00 | ± | 0.15 | 1.07 | ± | 0.07 | 0.88 | ± | 0.23 | 0.49 | ± | 0.14 | * p(diet) < 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, S.; Welch, T.N.; Aravindan, N.; Spann, R.A.; Welch, B.A.; Grayson, B.E. Altered Estrous Cyclicity and Feeding Neurocircuitry, but Not Cardiovascular Indices in Female Offspring from Dams with Previous Vertical Sleeve Gastrectomy Surgery. Brain Sci. 2023, 13, 1218. https://doi.org/10.3390/brainsci13081218
Johnson S, Welch TN, Aravindan N, Spann RA, Welch BA, Grayson BE. Altered Estrous Cyclicity and Feeding Neurocircuitry, but Not Cardiovascular Indices in Female Offspring from Dams with Previous Vertical Sleeve Gastrectomy Surgery. Brain Sciences. 2023; 13(8):1218. https://doi.org/10.3390/brainsci13081218
Chicago/Turabian StyleJohnson, Seth, Taylor N. Welch, Nandini Aravindan, Redin A. Spann, Bradley A. Welch, and Bernadette E. Grayson. 2023. "Altered Estrous Cyclicity and Feeding Neurocircuitry, but Not Cardiovascular Indices in Female Offspring from Dams with Previous Vertical Sleeve Gastrectomy Surgery" Brain Sciences 13, no. 8: 1218. https://doi.org/10.3390/brainsci13081218
APA StyleJohnson, S., Welch, T. N., Aravindan, N., Spann, R. A., Welch, B. A., & Grayson, B. E. (2023). Altered Estrous Cyclicity and Feeding Neurocircuitry, but Not Cardiovascular Indices in Female Offspring from Dams with Previous Vertical Sleeve Gastrectomy Surgery. Brain Sciences, 13(8), 1218. https://doi.org/10.3390/brainsci13081218