
How Freely Moving Mind Wandering Relates to Creativity: Behavioral and Neural Evidence

Abstract
1. Introduction
2. Methods
2.1. Participants
2.2. Behavioral Data
2.2.1. Freely Moving Mind Wandering Scale (FMMW-S)
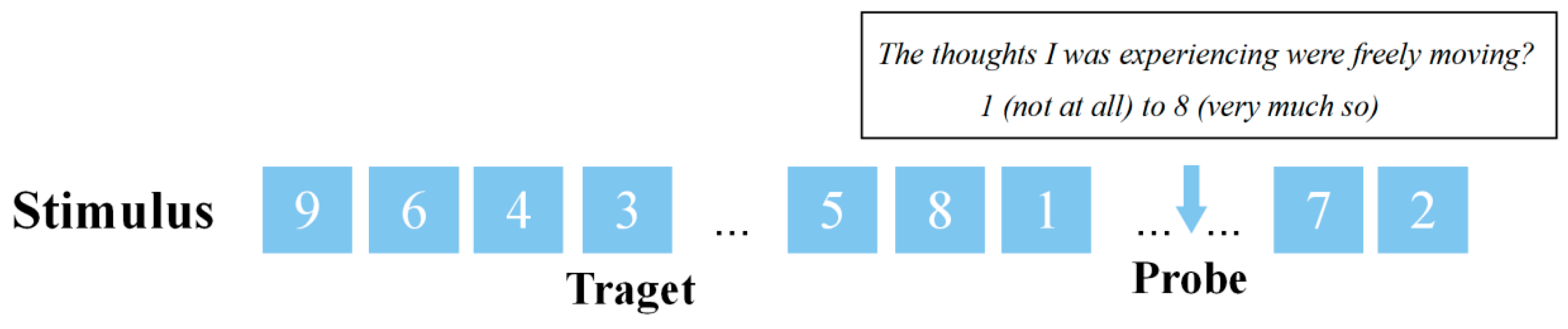
2.2.2. Sustained Attention to Response Task (SART)
2.2.3. Positive and Negative Affect Scale (PANAS)
2.2.4. Alternative Uses Task (AUT)
2.2.5. The Inventory of Creative Activities and Achievements (ICAA)
2.3. Image Acquisition and Preprocessing
2.4. Resting-State Time Series Extraction
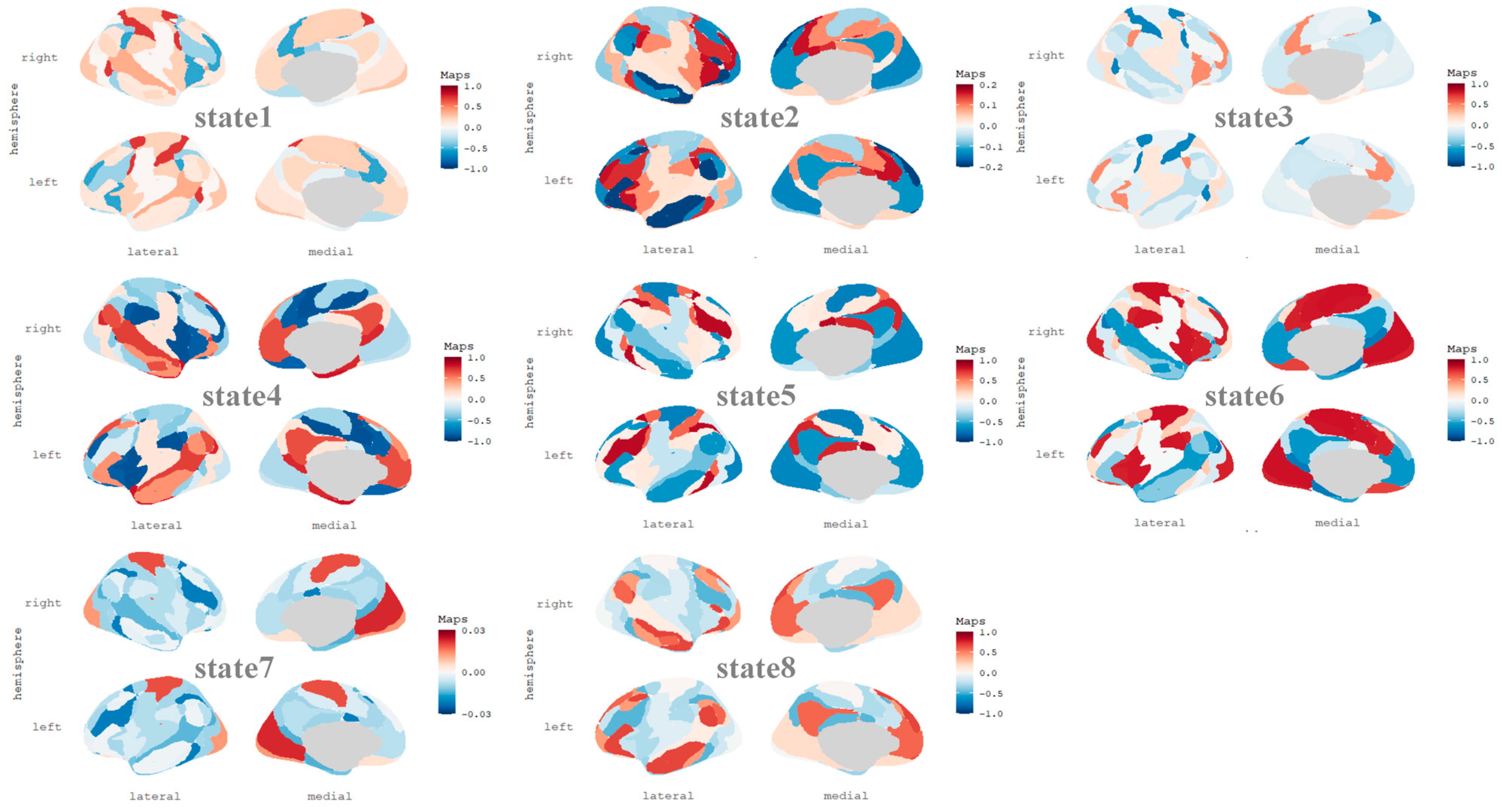
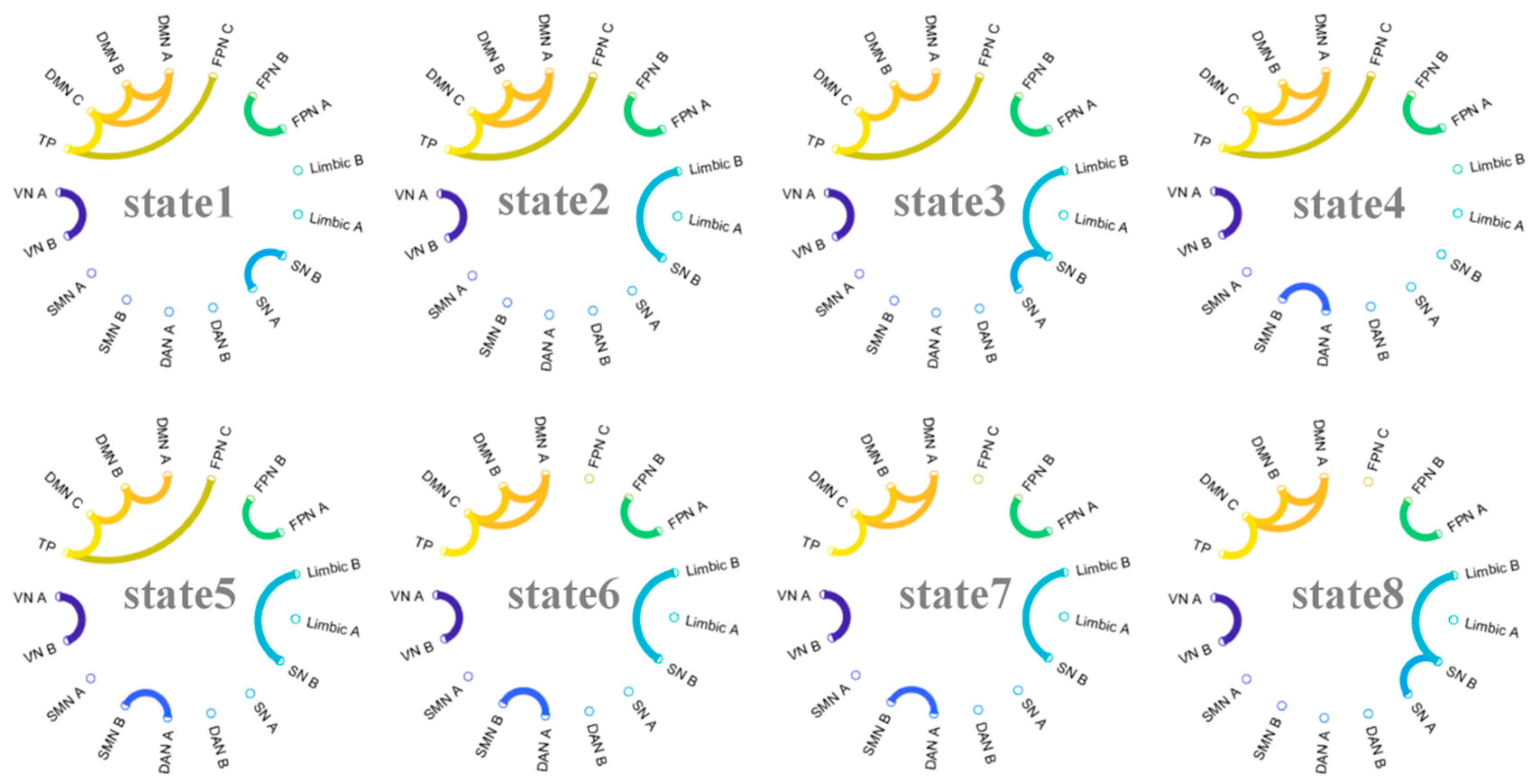
2.5. Hidden Markov Model (HMM)
2.6. Mediation Analysis
3. Results
3.1. Descriptive Analyses
3.2. Construct Validity
3.3. Relationship Between FMMW and Creativity
3.4. Brain State Lifetime Is Associated with Dynamic Features of FMMW
3.5. Mediation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Runco, M.A.; Jaeger, G.J. The Standard Definition of Creativity. Creat. Res. J. 2012, 24, 92–96. [Google Scholar] [CrossRef]
- Conner, T.S.; Silvia, P.J. Creative Days: A Daily Diary Study of Emotion, Personality, and Everyday Creativity. Psychol. Aesthet. Creat. Arts 2015, 9, 463–470. [Google Scholar] [CrossRef]
- Giannouli, V. Are Sex Differences in Self-estimated Intelligence an Elusive Phenomenon? Exploring the Role of Working Memory, Creativity, and Other Psychological Correlates in Young and Older Adults. Brain Behav. 2023, 13, e2857. [Google Scholar] [CrossRef] [PubMed]
- Beaty, R.E.; Silvia, P.J.; Nusbaum, E.C.; Jauk, E.; Benedek, M. The Roles of Associative and Executive Processes in Creative Cognition. Mem. Cogn. 2014, 42, 1186–1197. [Google Scholar] [CrossRef]
- Giannouli, V.; Tragantzopoulou, P. Creativity Is Not Related to Higher Cognitive Functioning: An Exploration Among Healthcare Professionals and Students During the COVID-19 Omicron Wave. SVOANE 2023, 4, 71–78. [Google Scholar] [CrossRef]
- Baird, B.; Smallwood, J.; Mrazek, M.D.; Kam, J.W.Y.; Franklin, M.S.; Schooler, J.W. Inspired by Distraction: Mind Wandering Facilitates Creative Incubation. Psychol. Sci. 2012, 23, 1117–1122. [Google Scholar] [CrossRef]
- Irving, Z.C.; McGrath, C.; Flynn, L.; Glasser, A.; Mills, C. The Shower Effect: Mind Wandering Facilitates Creative Incubation during Moderately Engaging Activities. Psychol. Aesthet. Creat. Arts 2022. Advance Online Publication. [Google Scholar] [CrossRef]
- Smeekens, B.A.; Kane, M.J. Working Memory Capacity, Mind Wandering, and Creative Cognition: An Individual-Differences Investigation into the Benefits of Controlled versus Spontaneous Thought. Psychol. Aesthet. Creat. Arts 2016, 10, 389–415. [Google Scholar] [CrossRef]
- Smith, A.P.; Brosowsky, N.; Murray, S.; Daniel, R.; Meier, M.E.; Seli, P. Fixation, Flexibility, and Creativity: The Dynamics of Mind Wandering. J. Exp. Psychol. Hum. Percept. Perform. 2022, 48, 689–710. [Google Scholar] [CrossRef]
- Steindorf, L.; Hammerton, H.A.; Rummel, J. Mind Wandering Outside the Box—About the Role of off-Task Thoughts and Their Assessment during Creative Incubation. Psychol. Aesthet. Creat. Arts 2021, 15, 584–595. [Google Scholar] [CrossRef]
- Christoff, K.; Irving, Z.C.; Fox, K.C.R.; Spreng, R.N.; Andrews-Hanna, J.R. Mind-Wandering as Spontaneous Thought: A Dynamic Framework. Nat. Rev. Neurosci. 2016, 17, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Girn, M.; Mills, C.; Roseman, L.; Carhart-Harris, R.L.; Christoff, K. Updating the Dynamic Framework of Thought: Creativity and Psychedelics. NeuroImage 2020, 213, 116726. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.; Raffaelli, Q.; Irving, Z.C.; Stan, D.; Christoff, K. Is an Off-Task Mind a Freely-Moving Mind? Examining the Relationship between Different Dimensions of Thought. Conscious. Cogn. 2018, 58, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, J.; Schooler, J.W. The Restless Mind. Psychol. Bull. 2006, 132, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Beaty, R.E.; Benedek, M.; Barry Kaufman, S.; Silvia, P.J. Default and Executive Network Coupling Supports Creative Idea Production. Sci. Rep. 2015, 5, 10964. [Google Scholar] [CrossRef]
- Beaty, R.E.; Benedek, M.; Silvia, P.J.; Schacter, D.L. Creative Cognition and Brain Network Dynamics. Trends Cogn. Sci. 2016, 20, 87–95. [Google Scholar] [CrossRef]
- Kam, J.W.Y.; Irving, Z.C.; Mills, C.; Patel, S.; Gopnik, A.; Knight, R.T. Distinct Electrophysiological Signatures of Task-Unrelated and Dynamic Thoughts. Proc. Natl. Acad. Sci. USA 2021, 118, e2011796118. [Google Scholar] [CrossRef]
- Kucyi, A.; Esterman, M.; Riley, C.S.; Valera, E.M. Spontaneous Default Network Activity Reflects Behavioral Variability Independent of Mind-Wandering. Proc. Natl. Acad. Sci. USA 2016, 113, 13899–13904. [Google Scholar] [CrossRef]
- Mason, M.F.; Norton, M.I.; Van Horn, J.D.; Wegner, D.M.; Grafton, S.T.; Macrae, C.N. Wandering Minds: The Default Network and Stimulus-Independent Thought. Science 2007, 315, 393–395. [Google Scholar] [CrossRef]
- Smallwood, J.; Schooler, J.W. The Science of Mind Wandering: Empirically Navigating the Stream of Consciousness. Annu. Rev. Psychol. 2015, 66, 487–518. [Google Scholar] [CrossRef]
- Christoff, K.; Gordon, A.M.; Smallwood, J.; Smith, R.; Schooler, J.W. Experience Sampling during fMRI Reveals Default Network and Executive System Contributions to Mind Wandering. Proc. Natl. Acad. Sci. USA 2009, 106, 8719–8724. [Google Scholar] [CrossRef] [PubMed]
- Fox, K.C.R.; Spreng, R.N.; Ellamil, M.; Andrews-Hanna, J.R.; Christoff, K. The Wandering Brain: Meta-Analysis of Functional Neuroimaging Studies of Mind-Wandering and Related Spontaneous Thought Processes. NeuroImage 2015, 111, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Kucyi, A.; Esterman, M.; Capella, J.; Green, A.; Uchida, M.; Biederman, J.; Gabrieli, J.D.E.; Valera, E.M.; Whitfield-Gabrieli, S. Prediction of Stimulus-Independent and Task-Unrelated Thought from Functional Brain Networks. Nat. Commun. 2021, 12, 1793. [Google Scholar] [CrossRef] [PubMed]
- Kucyi, A. Just a Thought: How Mind-Wandering Is Represented in Dynamic Brain Connectivity. NeuroImage 2018, 180, 505–514. [Google Scholar] [CrossRef]
- Mittner, M.; Boekel, W.; Tucker, A.M.; Turner, B.M.; Heathcote, A.; Forstmann, B.U. When the Brain Takes a Break: A Model-Based Analysis of Mind Wandering. J. Neurosci. 2014, 34, 16286–16295. [Google Scholar] [CrossRef]
- Kucyi, A.; Davis, K.D. Dynamic Functional Connectivity of the Default Mode Network Tracks Daydreaming. NeuroImage 2014, 100, 471–480. [Google Scholar] [CrossRef]
- Wang, X. Semantic Associative Abilities and Executive Control Functions Predict Novelty and Appropriateness of Idea Generation. Commun. Biol. 2024, 7, 703. [Google Scholar] [CrossRef]
- Fan, L.; Zhuang, K.; Wang, X.; Zhang, J.; Liu, C.; Gu, J.; Qiu, J. Exploring the Behavioral and Neural Correlates of Semantic Distance in Creative Writing. Psychophysiology 2023, 60, e14239. [Google Scholar] [CrossRef]
- Mashal, N.; Faust, M.; Hendler, T.; Jung-Beeman, M. An fMRI Investigation of the Neural Correlates Underlying the Processing of Novel Metaphoric Expressions. Brain Lang. 2007, 100, 115–126. [Google Scholar] [CrossRef]
- Beaty, R.E.; Kenett, Y.N.; Christensen, A.P.; Rosenberg, M.D.; Benedek, M.; Chen, Q.; Fink, A.; Qiu, J.; Kwapil, T.R.; Kane, M.J.; et al. Robust Prediction of Individual Creative Ability from Brain Functional Connectivity. Proc. Natl. Acad. Sci. USA 2018, 115, 1087–1092. [Google Scholar] [CrossRef]
- Mills, C.; Porter, A.R.; Andrews-Hanna, J.R.; Christoff, K.; Colby, A. How Task-Unrelated and Freely Moving Thought Relate to Affect: Evidence for Dissociable Patterns in Everyday Life. Emotion 2021, 21, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Vidaurre, D.; Quinn, A.J.; Baker, A.P.; Dupret, D.; Tejero-Cantero, A.; Woolrich, M.W. Spectrally Resolved Fast Transient Brain States in Electrophysiological Data. NeuroImage 2016, 126, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.H.; Warkentin, M.; Wallace, L. So Many Ways for Assessing Outliers: What Really Works and Does It Matter? J. Bus. Res. 2021, 132, 530–543. [Google Scholar] [CrossRef]
- Wei, L.; Hu, X.; Zhu, Y.; Yuan, Y.; Liu, W.; Chen, H. Aberrant Intra- and Internetwork Functional Connectivity in Depressed Parkinson’s Disease. Sci. Rep. 2017, 7, 2568. [Google Scholar] [CrossRef] [PubMed]
- Carriere, J.S.A.; Seli, P.; Smilek, D. Wandering in Both Mind and Body: Individual Differences in Mind Wandering and Inattention Predict Fidgeting. Can. J. Exp. Psychol./Rev. Can. De Psychol. Expérimentale 2013, 67, 19–31. [Google Scholar] [CrossRef]
- Robertson, I.H.; Manly, T.; Andrade, J.; Baddeley, B.T.; Yiend, J. ‘Oops!’: Performance Correlates of Everyday Attentional Failures in Traumatic Brain Injured and Normal Subjects. Neuropsychologia 1997, 35, 747–758. [Google Scholar] [CrossRef]
- Watson, D.; Anna, L.; Tellegen, A. Development and Validation of Brief Measures of Positive and Negative Affect: The PANAS Scales. J. Personal. Soc. Psychol. 1988, 54, 1063. [Google Scholar]
- Fink, A.; Benedek, M.; Grabner, R.H.; Staudt, B.; Neubauer, A.C. Creativity Meets Neuroscience: Experimental Tasks for the Neuroscientific Study of Creative Thinking. Methods 2007, 42, 68–76. [Google Scholar] [CrossRef]
- Diedrich, J.; Jauk, E.; Silvia, P.; Gredlein, J.; Neubauer, A.; Benedek, M. Assessment of Real-Life Creativity: The Inventory of Creative Activities and Achievements (ICAA). Psychol. Aesthet. Creat. Arts 2017, 12, 304. [Google Scholar] [CrossRef]
- Yan, C.; Zang, Y. DPARSF: A MATLAB Toolbox for “Pipeline” Data Analysis of Resting-State fMRI. Front. Syst. Neurosci. 2010, 4, 1377. [Google Scholar] [CrossRef]
- Schaefer, A.; Kong, R.; Gordon, E.M.; Laumann, T.O.; Zuo, X.-N.; Holmes, A.J.; Eickhoff, S.B.; Yeo, B.T.T. Local-Global Parcellation of the Human Cerebral Cortex from Intrinsic Functional Connectivity MRI. Cereb. Cortex 2018, 28, 3095–3114. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A Graph Theoretical Network Analysis Toolbox for Imaging Connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar]
- Vidaurre, D.; Smith, S.M.; Woolrich, M.W. Brain Network Dynamics Are Hierarchically Organized in Time. Proc. Natl. Acad. Sci. USA 2017, 114, 12827–12832. [Google Scholar] [CrossRef] [PubMed]
- Vidaurre, D.; Abeysuriya, R.; Becker, R.; Quinn, A.J.; Alfaro-Almagro, F.; Smith, S.M.; Woolrich, M.W. Discovering Dynamic Brain Networks from Big Data in Rest and Task. NeuroImage 2018, 180, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Hayes, A.F.; Preacher, K.J. Conditional Process Modeling: Using Structural Equation Modeling to Examine Contingent Causal Processes. In Structural Equation Modeling: A Second Course, 2nd ed.; Quantitative Methods in Education and the Behavioral Sciences: Issues, Research, and Teaching; IAP Information Age Publishing: Charlotte, NC, USA, 2013; pp. 219–266. ISBN 978-1-62396-244-9. [Google Scholar]
- Rouder, J.N.; Speckman, P.L.; Sun, D.; Morey, R.D.; Iverson, G. Bayesian t Tests for Accepting and Rejecting the Null Hypothesis. Psychon. Bull. Rev. 2009, 16, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Irving, Z.C. Mind-Wandering Is Unguided Attention: Accounting for the “Purposeful” Wanderer. Philos. Stud. 2016, 173, 547–571. [Google Scholar] [CrossRef]
- Liégeois, R. Resting Brain Dynamics at Different Timescales Capture Distinct Aspects of Human Behavior. Nat. Commun. 2019, 10, 2317. [Google Scholar] [CrossRef]
- Corbetta, M.; Patel, G.; Shulman, G.L. The Reorienting System of the Human Brain: From Environment to Theory of Mind. Neuron 2008, 58, 306–324. [Google Scholar] [CrossRef]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Sowden, P.T.; Pringle, A.; Gabora, L. The Shifting Sands of Creative Thinking: Connections to Dual-Process Theory. Think. Reason. 2015, 21, 40–60. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M | SD | Range | Skewness | Kurtosis | |
|---|---|---|---|---|---|
| FMMW-S | 4.96 | 1.38 | 2.00–7.00 | −0.46 | −0.53 |
| FMMW-SART | 4.78 | 1.67 | 1.00–8.00 | 0.13 | −0.69 |
| PANAS_PA | 28.27 | 7.79 | 10–45 | −0.28 | −0.20 |
| M | SD | Range | Skewness | Kurtosis | |
|---|---|---|---|---|---|
| FMMW-S | 4.95 | 1.22 | 2.00–7.00 | −0.37 | 0 |
| AUT_Flu | 6.00 | 2.16 | 1.40–12.70 | 0.55 | −0.02 |
| AUT_Fle | 4.83 | 1.53 | 1.00–9.83 | 0.37 | −0.09 |
| AUT_N | 15.41 | 6.20 | 2.00–35.67 | 0.62 | 0.07 |
| ICAA | 29.89 | 19.05 | 0–93.00 | 0.74 | 0.11 |
| 1 | 2 | 3 | |
|---|---|---|---|
| FMMW-S | - | ||
| FMMW-SART | 0.30 *** | - | |
| PANAS_PA | 0.21 ** | 0.24 ** | - |
| 1 | 2 | 3 | 4 | 5 | |
|---|---|---|---|---|---|
| FMMW-S | - | ||||
| AUT_Flu | 0.10 *** | - | |||
| AUT_Fle | 0.09 ** | 0.96 *** | - | ||
| AUT_N | 0.10 *** | 0.96 *** | 0.95 ** | - | |
| ICAA | 0.07 * | 0.24 *** | 0.24 ** | 0.24 ** | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Q.; Weng, L.; Geng, L.; Qiu, J. How Freely Moving Mind Wandering Relates to Creativity: Behavioral and Neural Evidence. Brain Sci. 2024, 14, 1122. https://doi.org/10.3390/brainsci14111122
Feng Q, Weng L, Geng L, Qiu J. How Freely Moving Mind Wandering Relates to Creativity: Behavioral and Neural Evidence. Brain Sciences. 2024; 14(11):1122. https://doi.org/10.3390/brainsci14111122
Chicago/Turabian StyleFeng, Qiuyang, Linman Weng, Li Geng, and Jiang Qiu. 2024. "How Freely Moving Mind Wandering Relates to Creativity: Behavioral and Neural Evidence" Brain Sciences 14, no. 11: 1122. https://doi.org/10.3390/brainsci14111122
APA StyleFeng, Q., Weng, L., Geng, L., & Qiu, J. (2024). How Freely Moving Mind Wandering Relates to Creativity: Behavioral and Neural Evidence. Brain Sciences, 14(11), 1122. https://doi.org/10.3390/brainsci14111122