
Review of EEG Affective Recognition with a Neuroscience Perspective

Abstract
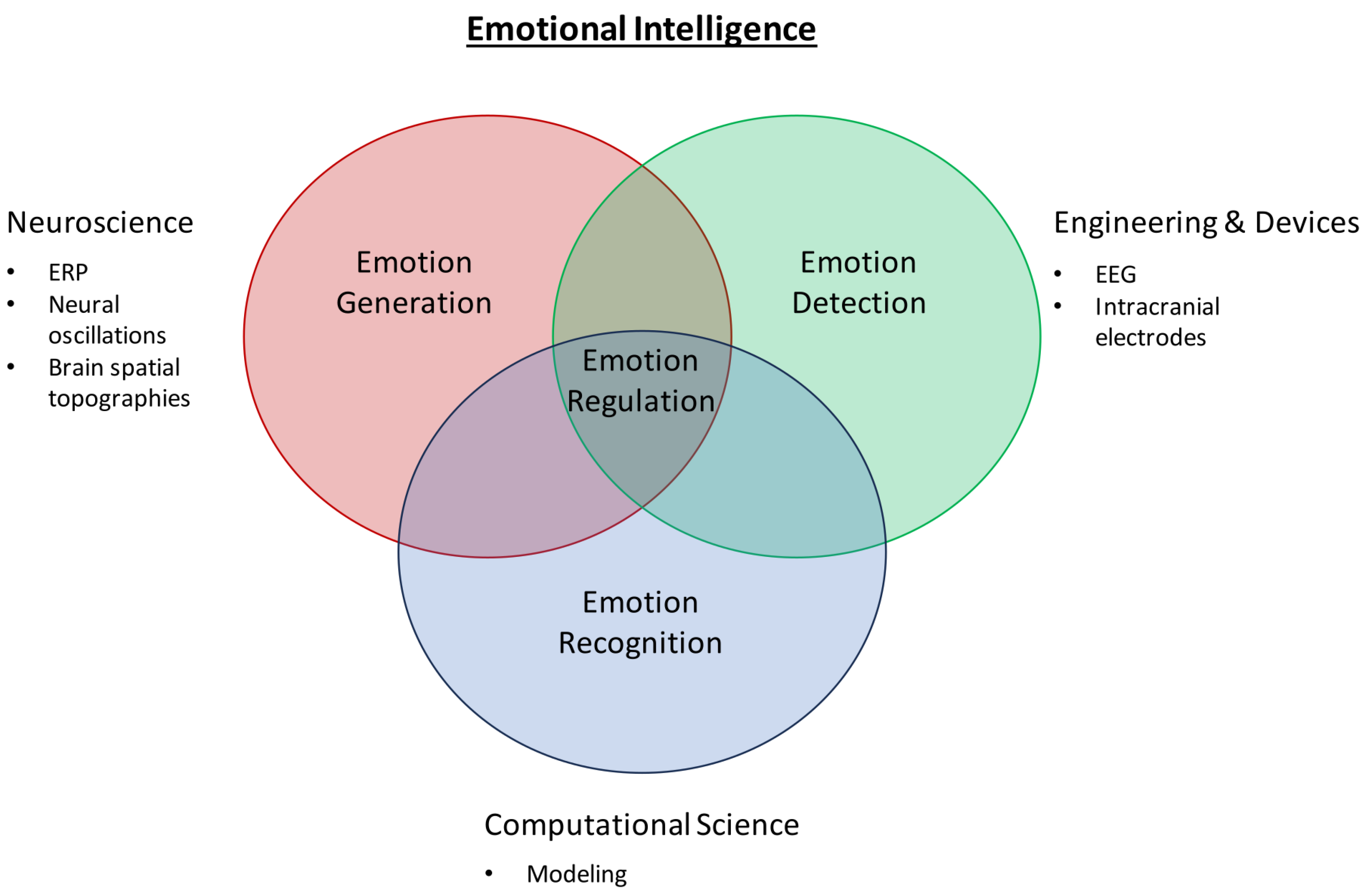
1. Introduction
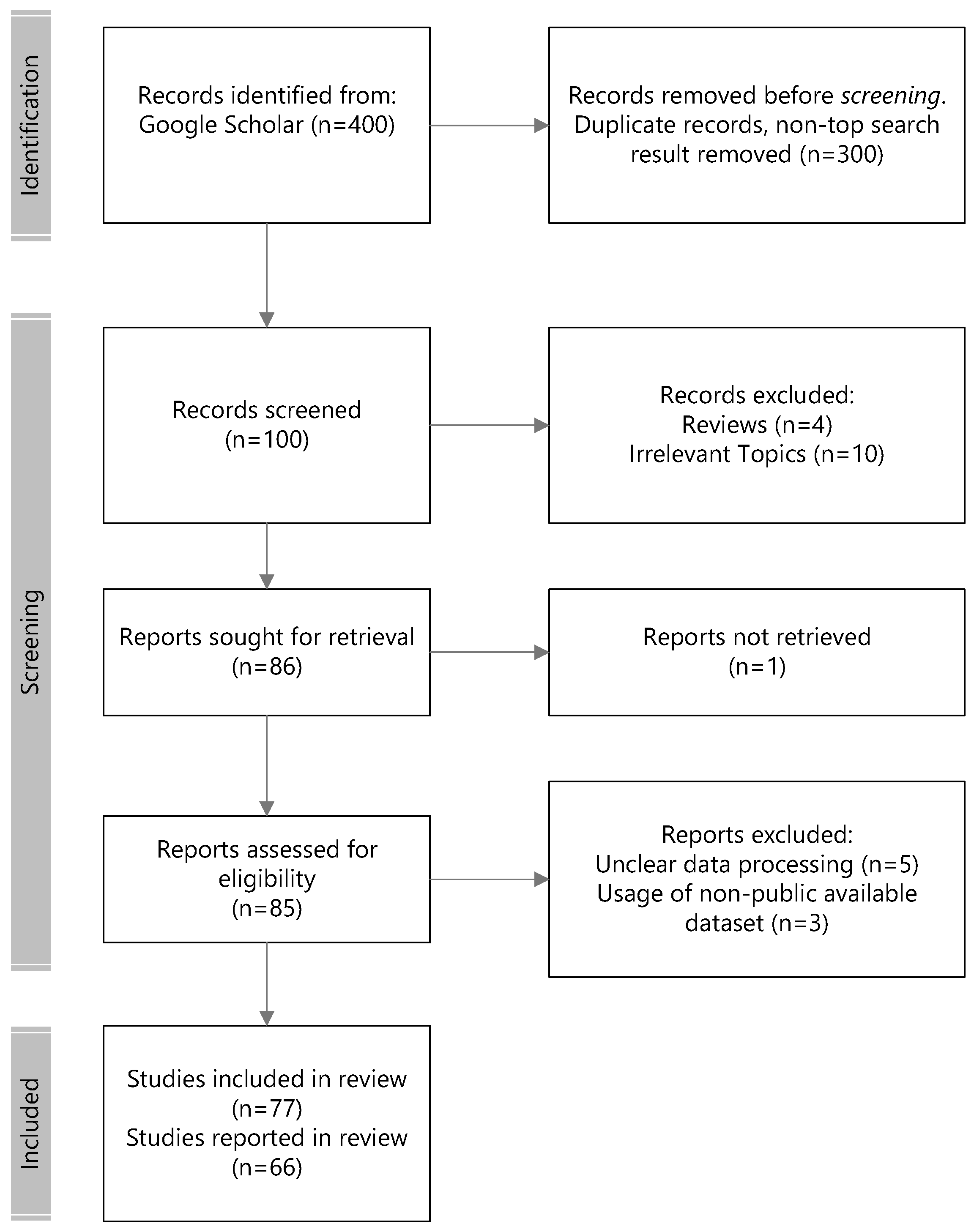
2. Methodology of the Review
3. Components of Emotions
3.1. Spatial Specificity of Emotions
3.2. Spatiotemporal Specificity of Emotions
3.2.1. How Are Brain Signals Generated?
3.2.2. What Information Is Transmitted in Brain Signals?
Insights from ERPs for Emotion Recognition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Data Acquisition Modality | Extracted Biomarker(s) | EEG Channels & Associated Brain Region(s) | Stimulus Modality for Facial Expressions | Emotion | Investigated Emotion Dimension(s) | Major Finding |
|---|---|---|---|---|---|---|---|
| Proverbio et al., 2020 [102] | EEG | P450, late anterior negativity response, late positivity (LPP) | RIGHT and LEFT frontal regions | Audio | Positive, Negative valence | Valence | P450 and LP enhanced by positive valence content; anterior negativity was enhanced by negative content; negative speech activated right temporo/parietal areas; positive speech activated left homologous and inferior frontal areas |
| Bondy et al., 2017 [103] | EEG | late positive potential (LPP) | Parietal region | Static images | Positive, Negative valence | Valence | Greater LPP for emotional images relative to neutral; negative valence images elicited greater LPP than positive valence images; greater LPP in younger subjects than older subjects |
| Dzhelyova et al., 2016 [100] | EEG | P100, N170, P300 | RIGHT dorsal occipito-temporal region (happiness), RIGHT ventral occipito-temporal region (fear and disgust) | Static images | Happy, Fear, Disgust, Neutral | Type of emotion, expression change detection | Unique spatial topography for each emotion type: dorsal occipito-temporal region (happiness), and ventral occipito-temporal region (fear and disgust); time-domain EEG markers apparent across all expression changes |
| Schirmer et al., 2012 [104] | EEG | P200 | Frontal, central, parietal regions | Visual words, and audio | Sad, Neutral | Word valence | Increased P200 amplitudes to sad intonation during encoding; voice-related retrieval effects observed in P200 using test words during retrieval |
| Schrammen et al., 2020 [105] | EEG | N170, alpha | Temporo-parietal region (N170), frontal region (alpha) | Static images | Happy, Anger | Valence | More negative amplitudes in the dominant hemisphere symmetrical to handedness; frontal alpha asymmetry is an index of inhibitory control; right hemisphere dominance for negative valence, left hemisphere dominance for positive valence |
| Chen et al., 2016 [101] | EEG | N100, N200, P300, alpha, beta, theta | Whole brain | Static images, audio, audio-visual pairing of static face image to audio sentence | Angry, Neutral | Type of emotion | Bimodal emotional changes detected with higher accuracy and faster reaction times; all emotional changes across modalities induced greater theta synchronization, enhanced amplitudes in N200/P300 complex; P300 and theta oscillations are important for emotional change detection |
| Martini et al., 2012 [108] | EEG | P300, LPP, gamma | LEFT/RIGHT temporal and frontal (negative valence), LEFT temporal and RIGHT parieto-temporal (neutral) | Static images of scenes | Negative valence, Neutral | Valence | Increase in P300 and LPP components for negative valence pictures across right hemisphere; enhanced gamma for negative valence pictures; increased gamma power for negative valence pictures; early strong between-region gamma synchronizations for negative valence pictures; late strong between-region gamma synchronization for neutral pictures |
| Eimer et al., 2003 [106] | EEG | N170, P200 | Frontocentral region (P200) | Static images | Angry, Disgusted, Fearful, Happy, Sad, Surprised, Neutral | Valence | N170 amplitudes displayed neither a main effect for valence or task in response to neutral vs emotional faces; sustained P200 elicited in response to emotional faces at frontocentral sites and then a broadly distributed P300 thereafter |
Insights from Evoked Oscillations for Emotion Recognition
| Study | Data Acquisition Modality | Extracted Biomarker(s) | EEG Channels & Associated Brain Region(s) | Stimulus Modality for Facial Expressions | Emotion | Investigated Emotion Dimension(s) | Major Finding |
|---|---|---|---|---|---|---|---|
| Schrammen et al., 2020 [105] | EEG | N170, alpha | Temporo-parietal region (N170), frontal region (alpha) | Static images | Happy, Anger | Valence | More negative amplitudes over the projected hemisphere according to the visual field for which the No-Go stimulus was presented; more negative amplitude in the dominant hemisphere symmetrical to handedness; frontal alpha asymmetry is an index of inhibitory control, frontal asymmetry not related to handedness; right hemisphere dominance for negative valence, left hemisphere dominance for positive valence |
| Chen et al., 2016 [101] | EEG | N100, N200, P300, alpha, beta, theta | Whole brain | Static images, audio, audio-visual pairing of static face image to audio sentence | Angry, Neutral | Type of emotion | Bimodal emotional changes detected with higher accuracy and faster reaction times than unimodal; all emotional changes across modalities induced greater theta synchronization, enhanced amplitudes in the N200/P300 complex, P300 amplitudes; P300 and theta oscillations crucial for emotional change detection; facial and bimodal change caused significant N100 enhancement and larger alpha/beta desynchronization |
| Zheng et al., 2019 [119] | Intracranial electrodes | theta, alpha | Amygdala and hippocampus (contralateral or outside of epileptogenic region in patients) | Static images of scenes | Positive, Negative valence | Valence | Incorrect response trials enhanced alpha power from amygdala modulates a decrease in both amygdala and hippocampal theta power and a decrease in high-frequency activity in the hippocampus; correct response trials decreased alpha power and enhanced theta power from amygdala synchronized with similarly enhanced theta power in hippocampus with high-frequency activity in both regions modulated by theta phase of the other; amygdala exerted a unidirectional influence on the hippocampus for memory retrieval via changes in alpha and theta band power |
| Tang et al., 2011 [120] | EEG | gamma (low and high) | LEFT/RIGHT/MIDDLE frontal, LEFT/RIGHT/MIDDLE centro-parietal, LEFT/RIGHT/MIDDLE occipito-temporal regions | Static images | Positive, Negative valence | Face-in-the-crowd expression search task among neutral faces | Early gamma activity (100–200 ms) decreased with increasing detection difficulty; late gamma activity (after 400 ms) increased with increasing detection difficulty |
| Schubring & Harald, 2020 [121] | EEG | alpha, beta | Occipitoparietal region | Static images of scenes | Positive, Negative valence | Arousal level | Both positive and negative valence pictures of high arousal levels associated with decreased alpha and lower beta band power; decrease in band power is associated with ERD response; a late ERS response observed for high-arousal negative pictures |
| Martini et al., 2012 [108] | EEG | P300, LPP, gamma | LEFT/RIGHT temporal and frontal (negative valence), LEFT temporal and RIGHT parieto-temporal (neutral) | Static images of scenes | Negative valence, Neutral | Valence | Increase in P300 and LPP components for negative valence across right hemisphere; enhanced gamma for negative valence; increased gamma power for negative valence with early low gamma and late gamma peaks across scalp over the whole duration; early strong between-region gamma synchronizations for negative valence between left and right temporal and frontal regions; late strong between-region gamma synchronization for neutral pictures between left temporal and right parieto-temporal regions |
| Güntekin & Basar, 2007 [117] | EEG | alpha, beta | LEFT posterior areas (T5, P3, O2), LEFT frontal areas (F3, CZ, C3) | Static images | Angry, Happy, Neutral | Type of emotion | Only for pictures with highest valence ratings from subjects: higher amplitude of alpha power for angry than happy expression at mostly left posterior hemisphere; higher amplitude of beta power for angry than happy expression at mostly left frontal hemisphere |
4. Affective Computing for EEG-Based Emotion Recongition
4.1. Experimental Protocols for Affective Computing
4.1.1. EEG Dataset
4.1.2. Data Preparation
4.1.3. Evaluation of Classification
4.2. Feature Engineering and Preprocessing Techniques
4.2.1. Feature Processing
4.2.2. Feature Engineering: Time Domain
4.2.3. Feature Engineering: Frequency Domain
4.2.4. Feature Engineering: Time–Frequency Domain
4.2.5. Feature Engineering: Nonlinear Domain
4.2.6. Feature Engineering: Spatial Dynamics Features
4.3. Recognition Models and Algorithm
4.3.1. Spatial Dynamics Recognition Models
4.3.2. Temporal Dynamics Recognition Models
4.3.3. Spatiotemporal Dynamics Recognition Models
4.3.4. Generalized Recognition Models
| Author | Dataset | Methodology | Experiment Results | Neuroscience Attribute |
|---|---|---|---|---|
| Atkinson and Campos, 2016 [24] | DEAP | Feature selection: Minimum-Redundancy-Maximum-Relevance (MRMR), Model: SVM classifier with RBF Kernel | Subject-Independent: DEAP: 2-Class (Valence-73.14%, Arousal-73.06%) | Generalized |
| Thammasan et al., 2016 [192] | EEGs induced by MIDI audio materials | Feature Processing: Fractal dimension (FD), power spectral density (PSD), discrete wavelet transform (DWT); windowing size (1-4/5-8 secs), Model: Deep belief network (DBN) | Subject-Dependent: 2-Class (Valence-88.24%, Arousal-82.42%) | Temporal |
| Li et al., 2016 [23] | DEAP | Feature Processing: CWT and Scalogram on multi-channels, Model: C-RNN; CNN for extracting features and LSTM to predict the emotion | 2 classes (Valence: 0.7206, Arousal: 0.7412) | Spatiotemporal |
| Yin et al., 2017 [197] | DEAP | Feature Processing: Statistical Features and PSD Model: Multi-Fusion-Layer based Ensemble Classifier of Stacked Autoencoder (MESAE) | Subject-Dependent: {(2-Class: (Valence-72.43%, Arousal-69.01%))} | Spatial |
| Li et al., 2018 [198] | SEED, SEED-IV | Feature Processing: DE; Model: Deep Adaptation Network (DAN) using MK-MMD losses | Subject-Independent: {(SEED: (DAN-83.81%, DANN-79.19%)), (SEED-IV: (DAN-58.87%, DANN-54.63%)} | Spatial |
| Yang et al., 2018 [199] | DEAP | Feature Processing: Raw EEG signal built into data mesh based on electrode topology; Model: Parallel Convolutional Recurrent Neural Network | Subject-Dependent: {(DEAP: 2-Class (Valence-90.80%, Arousal-91.03%))} | Spatiotemporal |
| Zhang et al., 2018 [200] | SEED | Feature Processing: DE, Model: Spatial Temporal RNN (STRNN) to learn spatial and temporal dependencies | Subject-Independent: {89.5%} | Spatiotemporal |
| Zheng et al., 2018 [153] | SEED-IV | Feature Processing: Differential Entropy (DE); Model: EmotionMeter | Subject-Dependent: {SEED-IV: 70.58%} | Spatial |
| Wang et al., 2018 [191] | SEED | Feature Processing: Differential Entropy (DE); Model: BDGLS | Subject-Dependent: {SEED: 93.66%} | Spatial |
| Li et al., 2018 [201] | SEED, SEED-IV | Feature Processing: DE; Model: Bi-hemispheres domain adversarial neural network (BiDANN) | Subject-Dependent: {92.38%}, Subject-Independent: {83.28%} | Spatiotemporal |
| Li et al., 2019 [202] | SEED | Feature Processing: DE; Model: R2G-STNN, based on BiLSTM to learn the spatial and temporal features | Subject-Dependent: {93.38%}, Subject-Independent: {84.16%} | Spatiotemporal |
| Zhong et al., 2020 [188] | SEED | Feature Processing: Differential Entropy (DE) smoothed by linear dynamic systems(LDS), Model: Regularized GNN (RGNN) with Emotion DL | Subject-Dependent: (SEED: 92.24%), (SEED-IV: 79.37%), Subject-independent modeling: (SEED: 85.30%), (SEED-IV: 73.84%) | Spatial |
| Song et al., 2020 [189] | SEED, DREAMER | Feature Processing: Using handcrafted features such as DE, PSD, DASM, RASM and DCAU based on frequency bands, Model: Dynamical graph convolutional neural network (DGCNN) | subject-dependent modeling (SEED: 90.4%, DREAMER-Valence: 86.23%, DREAMER-Arousal: 84.54%), subject-independent modeling (SEED: 79.95%) | Spatial |
| Lew et al., 2020 [28] | DEAP, SEED-IV | Feature Processing: RODAN extracted features; Model: RODAN, A spatial–temporal model with a domain adversarial network. | Subject-Dependent: {DEAP: 2-Class (Valence: 62.93%, Arousal: 63.97%), 4-Class (VA: 38.16%); SEED-IV: 70.28%}, Subject-Independent: {DEAP: 2-Class (Valence: 56.78%, Arousal: 56.60%), 4-Class (VA: 31.84%); SEED-IV: 60.75%} | Spatiotemporal |
| Duan et al., 2020 [203] | DEAP | Feature Processing: None. No handcrafted features; Model: CNN with Meta Update Strategy | Subject-Independent: {(DEAP:2 Class(Arousal: 66.5% )} | Generalized |
| Duan et al., 2020 [204] | SEED, DEAP | Feature Processing: No handcrafted Features. Features generated from CNN network; Models: MLCL that adopts meta learning approach | Subject-Independent: {(DEAP: 2-Class(Arousal 67.5%)), SEED: 3-Class( 78.6%))} | Spatiotemporal |
| Li et al., 2020 [205] | SEED, SEED-IV, MPED | Feature Processing: DE for SEED and SEED-IV, STFT for MPED, Model: Bi-hemispheric discrepancy model(BiHDM) capture the asymmetric characteristics between hemispheres | Subject-Dependent: {(SEED: 93.12%), (SEED-IV: 74.35%), ((MPED:40.34%) }, Subject-Independent: {(SEED: 85.4%, SEED-IV: 69.3%, (MPED: 28.27%)} | Spatiotemporal |
| Ding et al., 2021 [206] | SEED | Feature Processing: DE; Model: Task-specific Domain Adversarial Neural Network (T-DANN) an adversarial training method that transfer information inter- and intra-subject for prediction | Subject-Independent: { (cross-subject: 74.19%, cross-phase: 85.13%)} | Spatial |
| Wang et al., 2021 [207] | SEED, DEAP | Feature Processing: DE; Model: Few-label adversarial domain adaption (FLADA) | Subject-Independent: {(DEAP:68.0%), (SEED: 89.32%)} | Spatial |
| Miranda-Correa et al., 2021 [155] | AMIGOS | Feature Processing: PSD, PCA; Model: Gaussian Naïve Bayes Classifier | Subject-Independent: {(AMIGOS: 2Class(Valence-56.4%, Arousal 57.7%)} | Spatial |
| Liu et al., 2021 [156] | SEED, SEED-IV, SEED-V, DREAMER, DEAP | Feature Processing: Differential Entropy (DE); Model: DCCA | Subject-Dependent:{(SEED: 94.6%),(SEED-IV: 87.5% ), (SEED-V: 85.3%), (DREAMER: Valence-90.6%, Arousal-89.0%, Dominance-90.7%), (DEAP-Valence: 85.6%, DEAP-Arousal:84.3%)} | Spatial |
| Yin et al., 2021 [194] | DEAP | Feature Processing: DE Features, Model: GCNN-LSTM hybrid model, named ECLGCNN. | 2 classes, subject-dependent modeling (Valence: 90.45%, Arousal: 90.60%), subject-independent modeling (Valence: 84.81%, Arousal: 85.27%) | Spatiotemporal |
| Hu et al., 2021 [193] | DEAP, AMIGOS | Feature Processing: None, use Scaling Layer to generate spectrogram-like features Model: ScalingNet | Subject Independent: {(DEAP: 2-Class (Valence-71.88%, Arousal-71.8%, Dominance 73.67%))} | Temporal |
| Song et al., 2021 [195] | SEED, SDEA, DREAMER, MPED | Feature Processing: DE and PSD for SEED, PSD and HHS for SDEA, PSD for DREAMER and MPED; features converted into EEG-based images. Model: GECNN, graph-embedded convolutional neural network. | Subject-Dependent: {(SEED: 92.93% ), (SDEA: 79.69), (DREAMER: Valence-95.73%, Arousal-92.79%)), (MPED: 40.98%)} Subject-Independent: {(SEED: 92.93), ( SDEA: 53.31%} | Spatiotemporal |
| Liu et al., 2022 [208] | SEED | Feature Processing: No handcrafted Features, raw signal. Model: 3DCANN, 3D-CNN layers for 5 × 1 s raw EEG signals, then attention block and classification. | Subject-Dependent: {(SEED: 97.35%)}, Subject-Independent: {(SEED: 96.37%)} | Spatiotemporal |
| Liu et al., 2023 [209] | SEED, SEED-IV, DEAP | Feature Processing: DE for SEED and SEED-IV; Model: EeT | Subject-Dependent: {(DEAP:2-Class (Valence-92.86%, Arousal-93.34%)), (SEED: 96.28% ), (SEED-IV: 83.27%)} | Spatiotemporal |
| Li et al., 2023 [27] | SEED, Dreamer | Feature Processing: DE feature; Model: STGATE, a transformer-encoder is applied for capturing time-frequency features and graph attention | Subject-Independent: {(SEED: 90.37%); (SEED-IV: 76.43%); Dreamer: Valence: 77.44%, Arousal 75.26%, Average: 76.35%)} | Spatiotemporal |
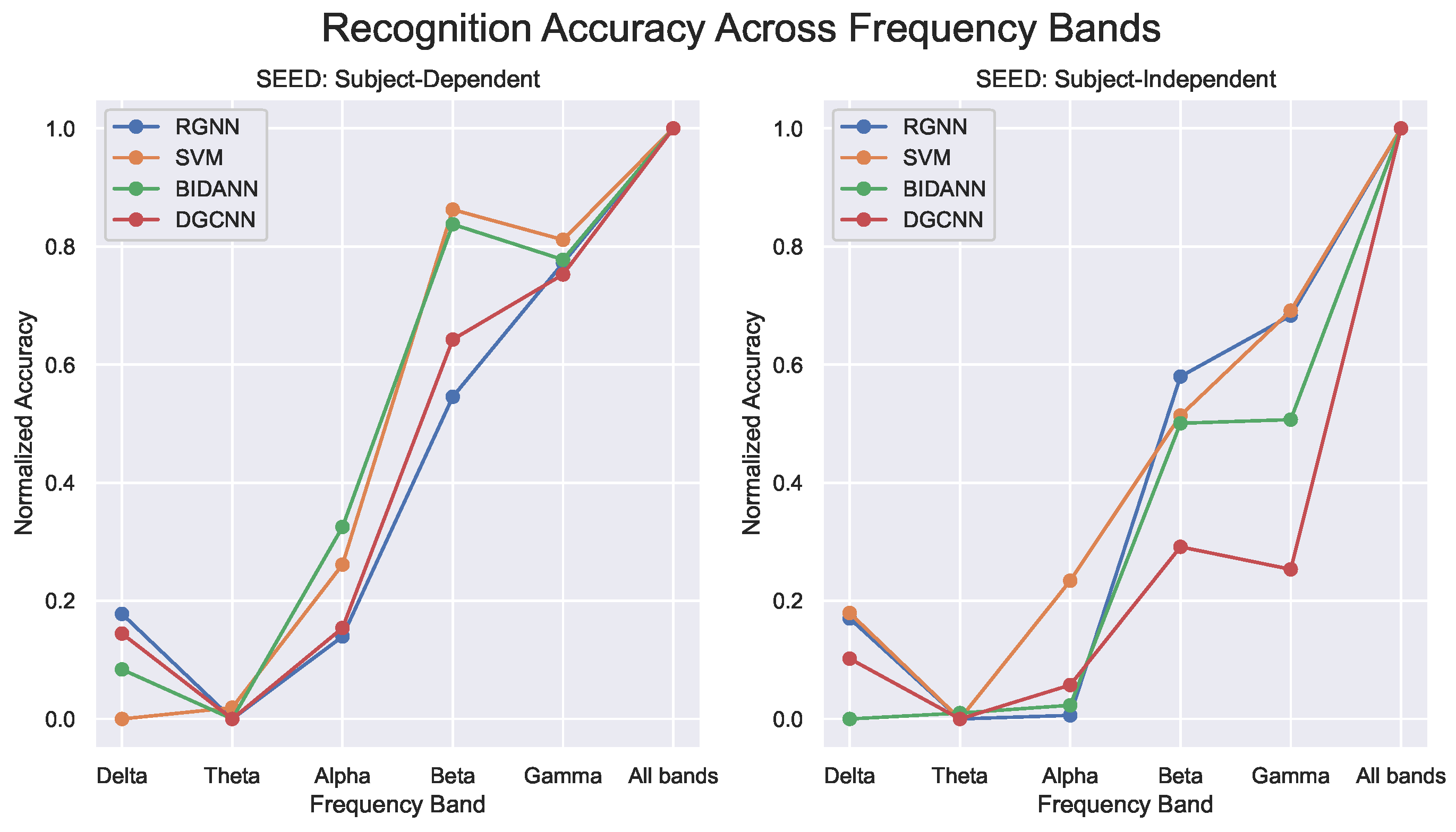
4.4. Evalutation of Recognition Models
5. Discussion
- To understand the “form” behind the “function”: given all the limitations in its basic components (i.e., neurons packed in limited physical space, the need for connectivity and computation efficiency, the presence of conduction delays over axonal branches, etc.), how the brain is eventually structured to overcome those limitations while maintaining an efficient communication mechanism.
- To understand that brain states are dynamic: functional membership of a neuron can change from one network pool to another under the influence of perturbation from an external stimulus and/or intrinsic population-wide neural activity that is, in turn, governed by different bands of oscillating rhythms.
- To know the advantages and limitations of emotion-related physiological markers: ERPs, alpha, beta, theta, delta, and gamma frequency bands and the information that can be interpreted or decoded from them.
- To avail oneself of the latest and widely adopted computational methods, algorithms, and models by the affective computing community for emotion recognition tasks.
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Barrett, L.F. Emotions are real. Emotion 2012, 12, 413–429. [Google Scholar] [CrossRef]
- Dalgleish, T. The emotional brain. Nat. Rev. Neurosci. 2004, 5, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.J. Emotion Regulation: Current Status and Future Prospects. Psychol. Inq. 2015, 26, 1–26. [Google Scholar] [CrossRef]
- Planalp, S. Varieties of Cues to Emotion in Naturally Occurring Situations. Cogn. Emot. 1996, 10, 137–154. [Google Scholar] [CrossRef]
- Scherer, K. Vocal communication of emotion: A review of research paradigms. Speech Commun. 2003, 40, 227–256. [Google Scholar] [CrossRef]
- World Health Organization. Mental Disorders. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/mental-disorders/?gad_source=1&gclid=Cj0KCQjwk6SwBhDPARIsAJ59GwfPLhgdxZysvhRHEbockNo7g8vHt3Y3TnHmhfEoJedNmyuLWbMre5AaAg-_EALw_wcB (accessed on 1 April 2024).
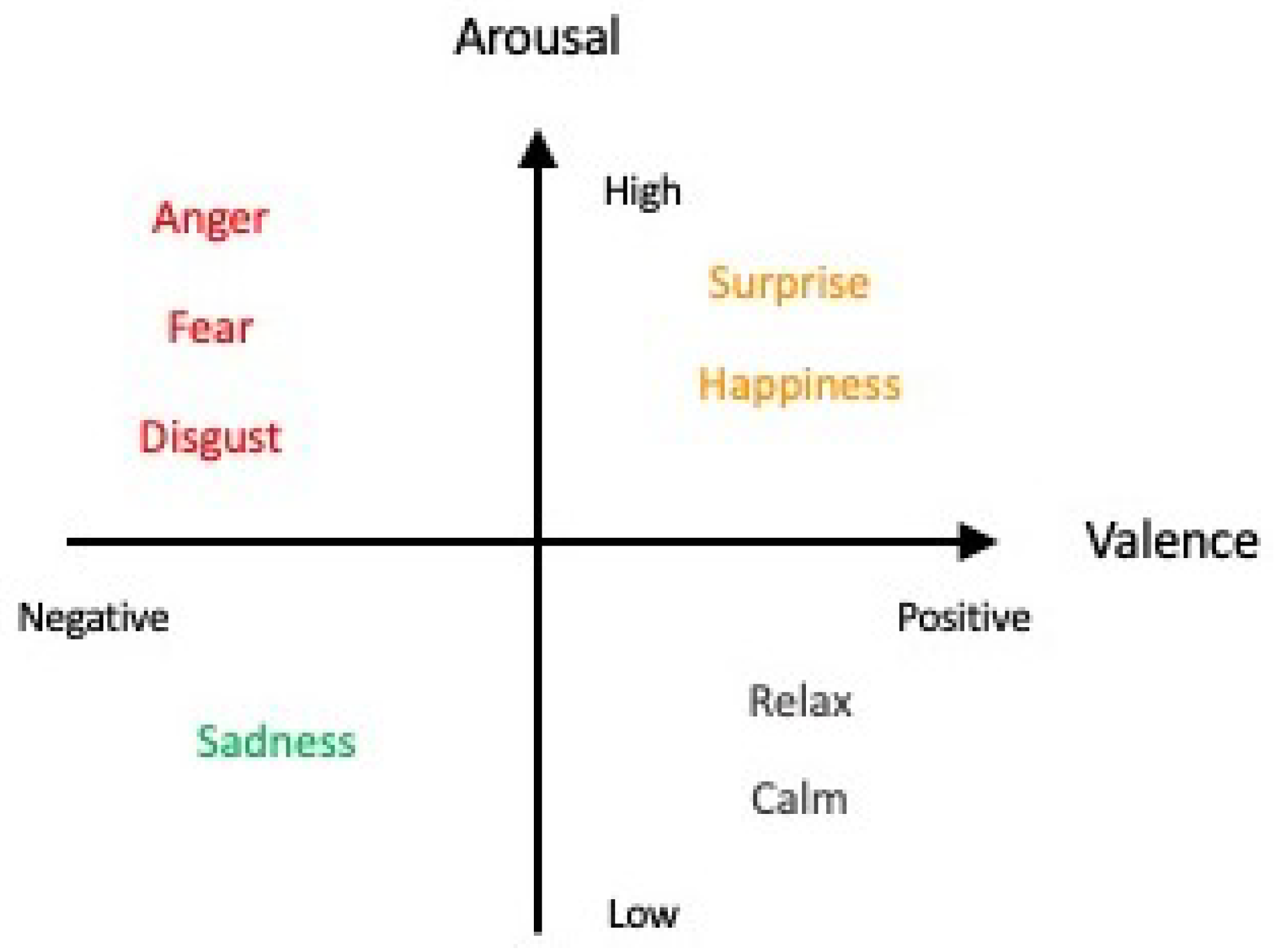
- Russell, J.A. A circumplex model of affect. J. Personal. Soc. Psychol. 1980, 39, 1161–1178. [Google Scholar] [CrossRef]
- Anderson, D.; Adolphs, R. A Framework for Studying Emotions across Species. Cell 2014, 157, 187–200. [Google Scholar] [CrossRef]
- Ekman, P. Are there basic emotions? Psychol. Rev. 1992, 99, 550–553. [Google Scholar] [CrossRef]
- Izard, C.E. Basic emotions, relations among emotions, and emotion-cognition relations. Psychol. Rev. 1992, 99, 561–565. [Google Scholar] [CrossRef]
- Izard, C.E. Basic Emotions, Natural Kinds, Emotion Schemas, and a New Paradigm. Perspect. Psychol. Sci. 2007, 2, 260–280. [Google Scholar] [CrossRef]
- Posner, J.; Russell, J.A.; Peterson, B.S. The circumplex model of affect: An integrative approach to affective neuroscience, cognitive development, and psychopathology. Dev. Psychopathol. 2005, 17, 715–734. [Google Scholar] [CrossRef]
- Levenson, R.W. Basic Emotion Questions. Emot. Rev. 2011, 3, 379–386. [Google Scholar] [CrossRef]
- Zhao, Y.; Xie, D.; Zhou, R.; Wang, N.; Yang, B. Evaluating users’ emotional experience in Mobile libraries: An emotional model based on the pleasure-arousal-dominance emotion model and the Five factor model. Front. Psychol. 2022, 13, 942198. [Google Scholar] [CrossRef] [PubMed]
- Mehrabian, A. Nonverbal Communication; Routledge: London, UK, 1972. [Google Scholar]
- Vytal, K.; Hamann, S. Neuroimaging Support for Discrete Neural Correlates of Basic Emotions: A Voxel-based Meta-analysis. J. Cogn. Neurosci. 2010, 22, 2864–2885. [Google Scholar] [CrossRef]
- Dzedzickis, A.; Kaklauskas, A.; Bucinskas, V. Human emotion recognition: Review of sensors and methods. Sensors 2020, 20, 592. [Google Scholar] [CrossRef]
- Horvers, A.; Tombeng, N.; Bosse, T.; Lazonder, A.W.; Molenaar, I. Detecting emotions through electrodermal activity in learning contexts: A systematic review. Sensors 2021, 21, 7869. [Google Scholar] [CrossRef] [PubMed]
- Veeranki, Y.R.; Kumar, H.; Ganapathy, N.; Natarajan, B.; Swaminathan, R. A Systematic Review of Sensing and Differentiating Dichotomous Emotional States Using Audio-Visual Stimuli. IEEE Access 2021, 9, 124434–124451. [Google Scholar] [CrossRef]
- Suhaimi, N.S.; Mountstephens, J.; Teo, J. EEG-based emotion recognition: A state-of-the-art review of current trends and opportunities. Comput. Intell. Neurosci. 2020, 2020, 8875426. [Google Scholar] [CrossRef]
- Posada-Quintero, H.F.; Chon, K.H. Phasic Component of Electrodermal Activity is more Correlated to Brain Activity than Tonic Component. In Proceedings of the 2019 IEEE EMBS International Conference on Biomedical & Health Informatics (BHI), Chicago, IL, USA, 19–22 May 2019; pp. 1–4. [Google Scholar] [CrossRef]
- Kroupi, E.; Vesin, J.M.; Ebrahimi, T. Phase-Amplitude Coupling between EEG and EDA While Experiencing Multimedia Content. In Proceedings of the 2013 Humaine Association Conference on Affective Computing and Intelligent Interaction, Geneva, Switzerland, 2–5 September 2013; pp. 865–870. [Google Scholar] [CrossRef]
- Li, X.; Song, D.; Zhang, P.; Yu, G.; Hou, Y.; Hu, B. Emotion recognition from multi-channel EEG data through Convolutional Recurrent Neural Network. In Proceedings of the 2016 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Shenzhen, China, 15–18 December 2016; pp. 352–359. [Google Scholar] [CrossRef]
- Atkinson, J.; Campos, D. Improving BCI-based emotion recognition by combining EEG feature selection and kernel classifiers. Expert Syst. Appl. 2016, 47, 35–41. [Google Scholar] [CrossRef]
- Shi, L.C.; Jiao, Y.Y.; Lu, B.L. Differential entropy feature for EEG-based vigilance estimation. In Proceedings of the 2013 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Osaka, Japan, 3–7 July 2013; pp. 6627–6630. [Google Scholar] [CrossRef]
- Lin, Y.P.; Wang, C.H.; Wu, T.L.; Jeng, S.K.; Chen, J.H. EEG-based emotion recognition in music listening: A comparison of schemes for multiclass support vector machine. In Proceedings of the 2009 IEEE International Conference on Acoustics, Speech and Signal Processing, Taipei, Taiwan, 19–24 April 2009; pp. 489–492. [Google Scholar] [CrossRef]
- Li, J.; Pan, W.; Huang, H.; Pan, J.; Wang, F. STGATE: Spatial-temporal graph attention network with a transformer encoder for EEG-based emotion recognition. Front. Hum. Neurosci. 2023, 17, 1169949. [Google Scholar] [CrossRef]
- Lew, W.C.L.; Wang, D.; Shylouskaya, K.; Zhang, Z.; Lim, J.H.; Ang, K.K.; Tan, A.H. EEG-based Emotion Recognition Using Spatial-Temporal Representation via Bi-GRU. In Proceedings of the 2020 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Montreal, QC, Canada, 20–24 July 2020; pp. 116–119. [Google Scholar] [CrossRef]
- Gross, J.J. Antecedent- and response-focused emotion regulation: Divergent consequences for experience, expression, and physiology. J. Personal. Soc. Psychol. 1998, 74, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Habel, U.; Klein, M.; Kellermann, T.; Shah, N.J.; Schneider, F. Same or different? Neural correlates of happy and sad mood in healthy males. NeuroImage 2005, 26, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Killgore, W.D.; Yurgelun-Todd, D.A. Activation of the amygdala and anterior cingulate during nonconscious processing of sad versus happy faces. NeuroImage 2004, 21, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Esslen, M.; Pascual-Marqui, R.; Hell, D.; Kochi, K.; Lehmann, D. Brain areas and time course of emotional processing. NeuroImage 2004, 21, 1189–1203. [Google Scholar] [CrossRef]
- Gorno-Tempini, M.L.; Pradelli, S.; Serafini, M.; Pagnoni, G.; Baraldi, P.; Porro, C.; Nicoletti, R.; Umità, C.; Nichelli, P. Explicit and Incidental Facial Expression Processing: An fMRI Study. NeuroImage 2001, 14, 465–473. [Google Scholar] [CrossRef]
- Kesler-West, M.L.; Andersen, A.H.; Smith, C.D.; Avison, M.J.; Davis, C.; Kryscio, R.J.; Blonder, L.X. Neural substrates of facial emotion processing using fMRI. Cogn. Brain Res. 2001, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- van de Riet, W.A.C.; Grèzes, J.; de Gelder, B. Specific and common brain regions involved in the perception of faces and bodies and the representation of their emotional expressions. Soc. Neurosci. 2009, 4, 101–120. [Google Scholar] [CrossRef]
- Fitzgerald, D.A.; Angstadt, M.; Jelsone, L.M.; Nathan, P.J.; Phan, K.L. Beyond threat: Amygdala reactivity across multiple expressions of facial affect. NeuroImage 2006, 30, 1441–1448. [Google Scholar] [CrossRef]
- Lange, K.; Williams, L.M.; Young, A.W.; Bullmore, E.T.; Brammer, M.J.; Williams, S.C.; Gray, J.A.; Phillips, M.L. Task instructions modulate neural responses to fearful facial expressions. Biol. Psychiatry 2003, 53, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Liddell, B.J.; Brown, K.J.; Kemp, A.H.; Barton, M.J.; Das, P.; Peduto, A.; Gordon, E.; Williams, L.M. A direct brainstem–amygdala–cortical ‘alarm’ system for subliminal signals of fear. NeuroImage 2005, 24, 235–243. [Google Scholar] [CrossRef]
- Phillips, M.L.; Williams, L.M.; Heining, M.; Herba, C.M.; Russell, T.; Andrew, C.; Bullmore, E.T.; Brammer, M.J.; Williams, S.C.; Morgan, M.; et al. Differential neural responses to overt and covert presentations of facial expressions of fear and disgust. NeuroImage 2004, 21, 1484–1496. [Google Scholar] [CrossRef]
- Thielscher, A.; Pessoa, L. Neural Correlates of Perceptual Choice and Decision Making during Fear–Disgust Discrimination. J. Neurosci. 2007, 27, 2908–2917. [Google Scholar] [CrossRef]
- Williams, L.M.; Das, P.; Liddell, B.; Olivieri, G.; Peduto, A.; Brammer, M.J.; Gordon, E. BOLD, sweat and fears: FMRI and skin conductance distinguish facial fear signals. NeuroReport 2005, 16, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Namburi, P.; Beyeler, A.; Yorozu, S.; Calhoon, G.G.; Halbert, S.A.; Wichmann, R.; Holden, S.S.; Mertens, K.L.; Anahtar, M.; Felix-Ortiz, A.C.; et al. A circuit mechanism for differentiating positive and negative associations. Nature 2015, 520, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Pignatelli, M.; Xu, S.; Itohara, S.; Tonegawa, S. Antagonistic negative and positive neurons of the basolateral amygdala. Nat. Neurosci. 2016, 19, 1636–1646. [Google Scholar] [CrossRef]
- Beyeler, A.; Namburi, P.; Glober, G.; Simonnet, C.; Calhoon, G.; Conyers, G.; Luck, R.; Wildes, C.; Tye, K. Divergent Routing of Positive and Negative Information from the Amygdala during Memory Retrieval. Neuron 2016, 90, 348–361. [Google Scholar] [CrossRef]
- Beyeler, A.; Chang, C.J.; Silvestre, M.; Lévêque, C.; Namburi, P.; Wildes, C.P.; Tye, K.M. Organization of Valence-Encoding and Projection-Defined Neurons in the Basolateral Amygdala. Cell Rep. 2018, 22, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Rolls, E.T. The orbitofrontal cortex. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1433–1444. [Google Scholar] [CrossRef]
- Fuster, J.M. Memory in the Cerebral Cortex: An Empirical Approach to Neural Networks in the Human and Nonhuman Primate; MIT Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Goldman-Rakic, P. The prefrontal landscape: Implications of functional architecture for understanding human mentation and the central executive. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1445–1453. [Google Scholar] [CrossRef]
- Miller, E.K.; Erickson, C.A.; Desimone, R. Neural Mechanisms of Visual Working Memory in Prefrontal Cortex of the Macaque. J. Neurosci. 1996, 16, 5154–5167. [Google Scholar] [CrossRef]
- Passingham, R.E. The Frontal Lobes and Voluntary Action; Oxford University Press: London, UK, 1993. [Google Scholar]
- Grafman, J. Similarities and Distinctions among Current Models of Prefrontal Cortical Functions. Ann. N. Y. Acad. Sci. 1995, 769, 337–368. [Google Scholar] [CrossRef]
- Wise, S.P.; Murray, E.A.; Gerfen, C.R. The Frontal Cortex-Basal Ganglia System in Primates. Crit. Rev. Neurobiol. 1996, 10, 317–356. [Google Scholar] [CrossRef] [PubMed]
- Asaad, W.F.; Rainer, G.; Miller, E.K. Neural Activity in the Primate Prefrontal Cortex during Associative Learning. Neuron 1998, 21, 1399–1407. [Google Scholar] [CrossRef]
- Schultz, W.; Dickinson, A. Neuronal Coding of Prediction Errors. Annu. Rev. Neurosci. 2000, 23, 473–500. [Google Scholar] [CrossRef] [PubMed]
- Barbas, H.; Pandya, D.N. Patterns of Connections of the Prefrontal Cortex in the Rhesus Monkey Associated with Cortical Architecture. In Frontal Lobe Function and Dysfunction; Oxford Academic: New York, NY, USA, 1991; pp. 35–58. [Google Scholar] [CrossRef]
- Pandya, D.N.; Yeterian, E.H. Prefrontal cortex in relation to other cortical areas in rhesus monkey: Architecture and connections. Prog. Brain Res. 1991, 85, 63–94. [Google Scholar] [CrossRef]
- Petrides, M.; Pandya, D.N. Dorsolateral prefrontal cortex: Comparative cytoarchitectonic analysis in the human and the macaque brain and corticocortical connection patterns. Eur. J. Neurosci. 1999, 11, 1011–1036. [Google Scholar] [CrossRef]
- Bruce, C.; Desimone, R.; Gross, C.G. Visual properties of neurons in a polysensory area in superior temporal sulcus of the macaque. J. Neurophysiol. 1981, 46, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Pandya, D.N.; Barnes, C.L. Architecture and Connections of the Frontal Lobe. In The Frontal Lobes Revisited; Psychology Press: London, UK, 2019; pp. 41–72. [Google Scholar] [CrossRef]
- Bates, J.F.; Goldman-Rakic, P.S. Prefrontal connections of medial motor areas in the rhesus monkey. J. Comp. Neurol. 1993, 336, 211–228. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Pandya, D.N. Anatomic Organization of the Basilar Pontine Projections from Prefrontal Cortices in Rhesus Monkey. J. Neurosci. 1997, 17, 438–458. [Google Scholar] [CrossRef]
- Barbas, H.; de Olmos, J. Projections from the amygdala to basoventral and mediodorsal prefrontal regions in the rhesus monkey. J. Comp. Neurol. 1990, 300, 549–571. [Google Scholar] [CrossRef]
- Kitamura, T.; Ogawa, S.K.; Roy, D.S.; Okuyama, T.; Morrissey, M.D.; Smith, L.M.; Redondo, R.L.; Tonegawa, S. Engrams and circuits crucial for systems consolidation of a memory. Science 2017, 356, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Phan, K.; Wager, T.; Taylor, S.F.; Liberzon, I. Functional Neuroanatomy of Emotion: A Meta-Analysis of Emotion Activation Studies in PET and fMRI. NeuroImage 2002, 16, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.C.; Nimmo-Smith, I.; Lawrence, A.D. Functional neuroanatomy of emotions: A meta-analysis. Cogn. Affect. Behav. Neurosci. 2003, 3, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Kober, H.; Barrett, L.F.; Joseph, J.; Bliss-Moreau, E.; Lindquist, K.; Wager, T.D. Functional grouping and cortical–subcortical interactions in emotion: A meta-analysis of neuroimaging studies. NeuroImage 2008, 42, 998–1031. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, K.A.; Wager, T.D.; Kober, H.; Bliss-Moreau, E.; Barrett, L.F. The brain basis of emotion: A meta-analytic review. Behav. Brain Sci. 2012, 35, 121–143. [Google Scholar] [CrossRef] [PubMed]
- Kotiyar, B.I.; Khludova, G.G.; Myasnikov, A.A.; Gusev, P.A.; Lyal’ka, V.F. Temporal specificity in the action of stimuli during the formation of associative ultrastructural reorganizations in neurons of the cerebral cortex. Neurosci. Behav. Physiol. 1990, 20, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Wainio-Theberge, S.; Evers, K. Spatiotemporal Neuroscience—What is it and why we need it. Phys. Life Rev. 2020, 33, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Baese, L.; Watters, H.; Keilholz, S. Spatiotemporal patterns of spontaneous brain activity: A mini-review. Neurophotonics 2022, 9, 032209. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L. The Tall Office Building Artistically Considered. Lippincott Mag. 1986, 57, 403–409. [Google Scholar]
- Hartmann, P.; Ramseier, A.; Gudat, F.; Mihatsch, M.J.; Polasek, W.; Geisenhoff, C. Normal weight of the brain in adults in relation to age, sex, body height and weight. Pathologe 1994, 15, 165–170. [Google Scholar] [CrossRef]
- Edelman, G.M. Neural Darwinism: The Theory of Neuronal Group Selection; Basic Books: New York, NY, USA, 1987. [Google Scholar]
- Cherniak, C. Neural component placement. Trends Neurosci. 1995, 18, 522–527. [Google Scholar] [CrossRef]
- Sporns, O.; Tononi, G.; Edelman, G. Theoretical Neuroanatomy: Relating Anatomical and Functional Connectivity in Graphs and Cortical Connection Matrices. Cereb. Cortex 2000, 10, 127–141. [Google Scholar] [CrossRef]
- Buzsàki, G.; Eidelberg, E. Commissural projection to the dentate gyrus of the rat: Evidence for feed-forward inhibition. Brain Res. 1981, 230, 346–350. [Google Scholar] [CrossRef]
- Buzsáki, G.; Eidelberg, E. Direct afferent excitation and long-term potentiation of hippocampal interneurons. J. Neurophysiol. 1982, 48, 597–607. [Google Scholar] [CrossRef]
- Buzsáki, G. Feed-forward inhibition in the hippocampal formation. Prog. Neurobiol. 1984, 22, 131–153. [Google Scholar] [CrossRef]
- Pouille, F.; Scanziani, M. Enforcement of Temporal Fidelity in Pyramidal Cells by Somatic Feed-Forward Inhibition. Science 2001, 293, 1159–1163. [Google Scholar] [CrossRef]
- Swadlow, H.A.; Gusev, A.G. Receptive-field construction in cortical inhibitory interneurons. Nat. Neurosci. 2002, 5, 403–404. [Google Scholar] [CrossRef]
- Steriade, M.; McCormick, D.A.; Sejnowski, T.J. Thalamocortical Oscillations in the Sleeping and Aroused Brain. Science 1993, 262, 679–685. [Google Scholar] [CrossRef]
- Gulyás, A.I.; Miles, R.; Sik, A.; Tóth, K.; Tamamaki, N.; Freund, T.F. Hippocampal pyramidal cells excite inhibitory neurons through a single release site. Nature 1993, 366, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Gulyás, A.I.; Mlles, R.; Hájos, N.; Freund, T.F. Precision and Variability in Postsynaptic Target Selection of Inhibitory Cells in the Hippocampal CA3 Region. Eur. J. Neurosci. 1993, 5, 1729–1751. [Google Scholar] [CrossRef] [PubMed]
- Csicsvari, J.; Hirase, H.; Czurko, A.; Buzsáki, G. Reliability and State Dependence of Pyramidal Cell–Interneuron Synapses in the Hippocampus. Neuron 1998, 21, 179–189. [Google Scholar] [CrossRef]
- Nusser, Z.; Mody, I. Selective Modulation of Tonic and Phasic Inhibitions in Dentate Gyrus Granule Cells. J. Neurophysiol. 2002, 87, 2624–2628. [Google Scholar] [CrossRef]
- Barthó, P.; Hirase, H.; Monconduit, L.; Zugaro, M.; Harris, K.D.; Buzsáki, G. Characterization of Neocortical Principal Cells and Interneurons by Network Interactions and Extracellular Features. J. Neurophysiol. 2004, 92, 600–608. [Google Scholar] [CrossRef]
- Silberberg, G.; Wu, C.; Markram, H. Synaptic dynamics control the timing of neuronal excitation in the activated neocortical microcircuit. J. Physiol. 2004, 556, 19–27. [Google Scholar] [CrossRef]
- Klausberger, T.; Magill, P.J.; Márton, L.F.; Roberts, J.D.B.; Cobden, P.M.; Buzsáki, G.; Somogyi, P. Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo. Nature 2003, 421, 844–848. [Google Scholar] [CrossRef]
- Marder, E.; Calabrese, R.L. Principles of rhythmic motor pattern generation. Physiol. Rev. 1996, 76, 687–717. [Google Scholar] [CrossRef]
- Traub, R.D.; Whittington, M.A.; Stanford, I.M.; Jefferys, J.G.R. A mechanism for generation of long-range synchronous fast oscillations in the cortex. Nature 1996, 383, 621–624. [Google Scholar] [CrossRef]
- Buzsáki, G.; Chrobak, J.J. Temporal structure in spatially organized neuronal ensembles: A role for interneuronal networks. Curr. Opin. Neurobiol. 1995, 5, 504–510. [Google Scholar] [CrossRef]
- D’amour, J.; Froemke, R. Inhibitory and Excitatory Spike-Timing-Dependent Plasticity in the Auditory Cortex. Neuron 2015, 86, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Hafting, T.; Fyhn, M.; Bonnevie, T.; Moser, M.B.; Moser, E.I. Hippocampus-independent phase precession in entorhinal grid cells. Nature 2008, 453, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- McClelland, J.L.; McNaughton, B.L.; O’Reilly, R.C. Why there are complementary learning systems in the hippocampus and neocortex: Insights from the successes and failures of connectionist models of learning and memory. Psychol. Rev. 1995, 102, 419–457. [Google Scholar] [CrossRef] [PubMed]
- Contreras, D.; Llinás, R. Voltage-Sensitive Dye Imaging of Neocortical Spatiotemporal Dynamics to Afferent Activation Frequency. J. Neurosci. 2001, 21, 9403–9413. [Google Scholar] [CrossRef] [PubMed]
- von Stein, A. Synchronization Between Temporal and Parietal Cortex During Multimodal Object Processing in Man. Cereb. Cortex 1999, 9, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Sarnthein, J.; Petsche, H.; Rappelsberger, P.; Shaw, G.L.; von Stein, A. Synchronization between prefrontal and posterior association cortex during human working memory. Proc. Natl. Acad. Sci. USA 1998, 95, 7092–7096. [Google Scholar] [CrossRef] [PubMed]
- Alekseichuk, I.; Turi, Z.; Amador de Lara, G.; Antal, A.; Paulus, W. Spatial Working Memory in Humans Depends on Theta and High Gamma Synchronization in the Prefrontal Cortex. Curr. Biol. 2016, 26, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, S.; Buzsáki, G. A 4 Hz Oscillation Adaptively Synchronizes Prefrontal, VTA, and Hippocampal Activities. Neuron 2011, 72, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Dzhelyova, M.; Jacques, C.; Rossion, B. At a Single Glance: Fast Periodic Visual Stimulation Uncovers the Spatio-Temporal Dynamics of Brief Facial Expression Changes in the Human Brain. Cereb. Cortex 2016, 27, 4106–4123. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Han, L.; Pan, Z.; Luo, Y.; Wang, P. Influence of attention on bimodal integration during emotional change decoding: ERP evidence. Int. J. Psychophysiol. 2016, 106, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Proverbio, A.M.; Santoni, S.; Adorni, R. ERP Markers of Valence Coding in Emotional Speech Processing. iScience 2020, 23, 100933. [Google Scholar] [CrossRef]
- Bondy, E.; Stewart, J.G.; Hajcak, G.; Weinberg, A.; Tarlow, N.; Mittal, V.A.; Auerbach, R.P. Emotion processing in female youth: Testing the stability of the late positive potential. Psychophysiology 2017, 55, e12977. [Google Scholar] [CrossRef]
- Schirmer, A.; Chen, C.B.; Ching, A.; Tan, L.; Hong, R.Y. Vocal emotions influence verbal memory: Neural correlates and interindividual differences. Cogn. Affect. Behav. Neurosci. 2012, 13, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Schrammen, E.; Grimshaw, G.M.; Berlijn, A.M.; Ocklenburg, S.; Peterburs, J. Response inhibition to emotional faces is modulated by functional hemispheric asymmetries linked to handedness. Brain Cogn. 2020, 145, 105629. [Google Scholar] [CrossRef] [PubMed]
- Eimer, M.; Holmes, A.; McGlone, F.P. The role of spatial attention in the processing of facial expression: An ERP study of rapid brain responses to six basic emotions. Cogn. Affect. Behav. Neurosci. 2003, 3, 97–110. [Google Scholar] [CrossRef]
- Ashley, V.; Vuilleumier, P.; Swick, D. Time course and specificity of event-related potentials to emotional expressions. NeuroReport 2004, 15, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Martini, N.; Menicucci, D.; Sebastiani, L.; Bedini, R.; Pingitore, A.; Vanello, N.; Milanesi, M.; Landini, L.; Gemignani, A. The dynamics of EEG gamma responses to unpleasant visual stimuli: From local activity to functional connectivity. NeuroImage 2012, 60, 922–932. [Google Scholar] [CrossRef]
- Krolak-Salmon, P.; Hénaff, M.A.; Vighetto, A.; Bertrand, O.; Mauguière, F. Early Amygdala Reaction to Fear Spreading in Occipital, Temporal, and Frontal Cortex. Neuron 2004, 42, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.; Bar, M. See it with feeling: Affective predictions during object perception. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Pourtois, G.; Spinelli, L.; Seeck, M.; Vuilleumier, P. Temporal precedence of emotion over attention modulations in the lateral amygdala: Intracranial ERP evidence from a patient with temporal lobe epilepsy. Cogn. Affect. Behav. Neurosci. 2010, 10, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, N.; Kawasaki, H.; Oya, H.; Howard, M.A.; Adolphs, R. Decoding Face Information in Time, Frequency and Space from Direct Intracranial Recordings of the Human Brain. PLoS ONE 2008, 3, e3892. [Google Scholar] [CrossRef]
- Jung, J.; Bayle, D.; Jerbi, K.; Vidal, J.R.; Hénaff, M.A.; Ossandon, T.; Bertrand, O.; Mauguière, F.; Lachaux, J.P. Intracerebral gamma modulations reveal interaction between emotional processing and action outcome evaluation in the human orbitofrontal cortex. Int. J. Psychophysiol. 2011, 79, 64–72. [Google Scholar] [CrossRef]
- Protachevicz, P.R.; Borges, F.S.; Iarosz, K.C.; Baptista, M.S.; Lameu, E.L.; Hansen, M.; Caldas, I.L.; Szezech, J.D.; Batista, A.M.; Kurths, J. Influence of delayed conductance on neuronal synchronization. Front. Physiol. 2020, 11, 1053. [Google Scholar] [CrossRef]
- Atasoy, S.; Deco, G.; Kringelbach, M.L. Harmonic waves as the fundamental principle underlying temporo-spatial dynamicsof brain and mind: Comment on “Is temporo-spatial dynamics the “common currency” of brainand mind? In Quest of “Spatiotemporal Neuroscience”” Georg Northoff et al. Phys. Life Rev. 2020, 33, 67–69. [Google Scholar] [CrossRef]
- Buzsáki, G. Rhythms of the Brain; Oxford University Press: Oxford, UK, 2006. [Google Scholar] [CrossRef]
- Güntekin, B.; Basar, E. Emotional face expressions are differentiated with brain oscillations. Int. J. Psychophysiol. 2007, 64, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Popov, T.; Miller, G.A.; Rockstroh, B.; Weisz, N. Modulation of α Power and Functional Connectivity during Facial Affect Recognition. J. Neurosci. 2013, 33, 6018–6026. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Stevenson, R.F.; Mander, B.A.; Mnatsakanyan, L.; Hsu, F.P.; Vadera, S.; Knight, R.T.; Yassa, M.A.; Lin, J.J. Multiplexing of Theta and Alpha Rhythms in the Amygdala-Hippocampal Circuit Supports Pattern Separation of Emotional Information. Neuron 2019, 102, 887–898.e5. [Google Scholar] [CrossRef]
- Tang, Y.; Li, Y.; Wang, J.; Tong, S.; Li, H.; Yan, J. Induced gamma activity in EEG represents cognitive control during detecting emotional expressions. In Proceedings of the 2011 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Boston, MA, USA, 30 August–3 September 2011. [Google Scholar] [CrossRef]
- Schubring, D.; Schupp, H.T. Emotion and Brain Oscillations: High Arousal is Associated with Decreases in Alpha- and Lower Beta-Band Power. Cereb. Cortex 2020, 31, 1597–1608. [Google Scholar] [CrossRef]
- Berger, A.; Pixa, N.; Doppelmayr, M. P95 Frequency-specific after-effects of transcranial alternating current stimulation (tACS) on motor learning. Clin. Neurophysiol. 2017, 128, e376. [Google Scholar] [CrossRef]
- Clayton, M.S.; Yeung, N.; Cohen Kadosh, R. Electrical stimulation of alpha oscillations stabilizes performance on visual attention tasks. J. Exp. Psychol. Gen. 2019, 148, 203–220. [Google Scholar] [CrossRef]
- van Schouwenburg, M.R.; Zanto, T.P.; Gazzaley, A. Spatial Attention and the Effects of Frontoparietal Alpha Band Stimulation. Front. Hum. Neurosci. 2017, 10, 658. [Google Scholar] [CrossRef]
- Schack, B.; Klimesch, W.; Sauseng, P. Phase synchronization between theta and upper alpha oscillations in a working memory task. Int. J. Psychophysiol. 2005, 57, 105–114. [Google Scholar] [CrossRef]
- Haenschel, C.; Baldeweg, T.; Croft, R.J.; Whittington, M.; Gruzelier, J. Gamma and beta frequency oscillations in response to novel auditory stimuli: A comparison of human electroencephalogram (EEG) data with in vitro models. Proc. Natl. Acad. Sci. USA 2000, 97, 7645–7650. [Google Scholar] [CrossRef] [PubMed]
- Kisley, M.A.; Cornwell, Z.M. Gamma and beta neural activity evoked during a sensory gating paradigm: Effects of auditory, somatosensory and cross-modal stimulation. Clin. Neurophysiol. 2006, 117, 2549–2563. [Google Scholar] [CrossRef]
- Engel, A.K.; Fries, P. Beta-band oscillations—Signalling the status quo? Curr. Opin. Neurobiol. 2010, 20, 156–165. [Google Scholar] [CrossRef]
- Güntekin, B.; Basar, E. Event-related beta oscillations are affected by emotional eliciting stimuli. Neurosci. Lett. 2010, 483, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, C.C.; Daut, R.; Brower, M.; Bragg, A. Electroencephalographic α-band and β-band correlates of perspective-taking and personal distress. NeuroReport 2011, 22, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Miskovic, V.; Schmidt, L.A. Cross-regional cortical synchronization during affective image viewing. Brain Res. 2010, 1362, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Aktürk, T.; de Graaf, T.A.; Abra, Y.; Şahoğlu Göktaş, S.; Özkan, D.; Kula, A.; Güntekin, B. Event-related EEG oscillatory responses elicited by dynamic facial expression. BioMed. Eng. OnLine 2021, 20, 1217178. [Google Scholar] [CrossRef] [PubMed]
- Zion-Golumbic, E.; Kutas, M.; Bentin, S. Neural Dynamics Associated with Semantic and Episodic Memory for Faces: Evidence from Multiple Frequency Bands. J. Cogn. Neurosci. 2010, 22, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Basar, E.; Özgören, M.; Öniz, A.; Schmiedt, C.; Başar-Eroğlu, C. Brain oscillations differentiate the picture of one’s own grandmother. Int. J. Psychophysiol. 2007, 64, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kendrick, K.M.; Zhou, H.; Zhan, Y.; Feng, J. A Computational Study on Altered Theta-Gamma Coupling during Learning and Phase Coding. PLoS ONE 2012, 7, e36472. [Google Scholar] [CrossRef]
- Knyazev, G.; Slobodskoj-Plusnin, J.; Bocharov, A. Event-related delta and theta synchronization during explicit and implicit emotion processing. Neuroscience 2009, 164, 1588–1600. [Google Scholar] [CrossRef] [PubMed]
- González-Roldan, A.M.; Martínez-Jauand, M.; Muñoz-García, M.A.; Sitges, C.; Cifre, I.; Montoya, P. Temporal dissociation in the brain processing of pain and anger faces with different intensities of emotional expression. Pain 2011, 152, 853–859. [Google Scholar] [CrossRef]
- Jaušovec, N.; Jaušovec, K.; Pahor, A. The influence of theta transcranial alternating current stimulation (tACS) on working memory storage and processing functions. Acta Psychol. 2014, 146, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Köster, M.; Finger, H.; Graetz, S.; Kater, M.; Gruber, T. Theta-gamma coupling binds visual perceptual features in an associative memory task. Sci. Rep. 2018, 8, 17688. [Google Scholar] [CrossRef] [PubMed]
- Klados, M.A.; Frantzidis, C.; Vivas, A.B.; Papadelis, C.; Lithari, C.; Pappas, C.; Bamidis, P.D. A Framework Combining Delta Event-Related Oscillations (EROs) and Synchronisation Effects (ERD/ERS) to Study Emotional Processing. Comput. Intell. Neurosci. 2009, 2009, 549419. [Google Scholar] [CrossRef] [PubMed]
- Balconi, M.; Lucchiari, C. Consciousness and arousal effects on emotional face processing as revealed by brain oscillations. A gamma band analysis. Int. J. Psychophysiol. 2008, 67, 41–46. [Google Scholar] [CrossRef]
- Balconi, M.; Pozzoli, U. Arousal effect on emotional face comprehension. Physiol. Behav. 2009, 97, 455–462. [Google Scholar] [CrossRef]
- Öhman, A.; Flykt, A.; Esteves, F. Emotion drives attention: Detecting the snake in the grass. J. Exp. Psychol. Gen. 2001, 130, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Holroyd, T.; Jones, M.; Hendler, T.; Blair, J. Neural dynamics for facial threat processing as revealed by gamma band synchronization using MEG. NeuroImage 2007, 34, 839–847. [Google Scholar] [CrossRef]
- Sato, W.; Kochiyama, T.; Uono, S.; Matsuda, K.; Usui, K.; Inoue, Y.; Toichi, M. Rapid amygdala gamma oscillations in response to fearful facial expressions. Neuropsychologia 2011, 49, 612–617. [Google Scholar] [CrossRef]
- Pike, F.G.; Goddard, R.S.; Suckling, J.M.; Ganter, P.; Kasthuri, N.; Paulsen, O. Distinct frequency preferences of different types of rat hippocampal neurones in response to oscillatory input currents. J. Physiol. 2000, 529, 205–213. [Google Scholar] [CrossRef]
- Markram, H.; Toledo-Rodriguez, M.; Wang, Y.; Gupta, A.; Silberberg, G.; Wu, C. Interneurons of the neocortical inhibitory system. Nat. Rev. Neurosci. 2004, 5, 793–807. [Google Scholar] [CrossRef]
- Bressler, S.L.; Kelso, J. Cortical coordination dynamics and cognition. Trends Cogn. Sci. 2001, 5, 26–36. [Google Scholar] [CrossRef]
- Koelstra, S.; Muhl, C.; Soleymani, M.; Lee, J.S.; Yazdani, A.; Ebrahimi, T.; Pun, T.; Nijholt, A.; Patras, I. DEAP: A Database for Emotion Analysis; Using Physiological Signals. IEEE Trans. Affect. Comput. 2012, 3, 18–31. [Google Scholar] [CrossRef]
- Soleymani, M.; Lichtenauer, J.; Pun, T.; Pantic, M. A Multimodal Database for Affect Recognition and Implicit Tagging. IEEE Trans. Affect. Comput. 2012, 3, 42–55. [Google Scholar] [CrossRef]
- Zheng, W.L.; Lu, B.L. Investigating Critical Frequency Bands and Channels for EEG-Based Emotion Recognition with Deep Neural Networks. IEEE Trans. Auton. Ment. Dev. 2015, 7, 162–175. [Google Scholar] [CrossRef]
- Katsigiannis, S.; Ramzan, N. DREAMER: A Database for Emotion Recognition Through EEG and ECG Signals from Wireless Low-cost Off-the-Shelf Devices. IEEE J. Biomed. Health Inform. 2018, 22, 98–107. [Google Scholar] [CrossRef]
- Zheng, W.L.; Liu, W.; Lu, Y.; Lu, B.L.; Cichocki, A. EmotionMeter: A Multimodal Framework for Recognizing Human Emotions. IEEE Trans. Cybern. 2019, 49, 1110–1122. [Google Scholar] [CrossRef]
- Song, T.; Zheng, W.; Lu, C.; Zong, Y.; Zhang, X.; Cui, Z. MPED: A Multi-Modal Physiological Emotion Database for Discrete Emotion Recognition. IEEE Access 2019, 7, 12177–12191. [Google Scholar] [CrossRef]
- Miranda-Correa, J.A.; Abadi, M.K.; Sebe, N.; Patras, I. AMIGOS: A Dataset for Affect, Personality and Mood Research on Individuals and Groups. IEEE Trans. Affect. Comput. 2021, 12, 479–493. [Google Scholar] [CrossRef]
- Liu, W.; Qiu, J.L.; Zheng, W.L.; Lu, B.L. Comparing Recognition Performance and Robustness of Multimodal Deep Learning Models for Multimodal Emotion Recognition. IEEE Trans. Cogn. Dev. Syst. 2022, 14, 715–729. [Google Scholar] [CrossRef]
- Li, X.; Zbhang, Y.; Tiwari, P.; Song, D.; Hu, B.; Yang, M.; Zhao, Z.; Kumar, N.; Marttinen, P. EEG Based Emotion Recognition: A Tutorial and Review. ACM Comput. Surv. 2022, 55, 79. [Google Scholar] [CrossRef]
- Lew, W.C.L.; Wang, D.; Ang, K.K.; Lim, J.H.; Quek, C.; Tan, A.H. EEG-Video Emotion-Based Summarization: Learning With EEG Auxiliary Signals. IEEE Trans. Affect. Comput. 2022, 13, 1827–1839. [Google Scholar] [CrossRef]
- Hjorth, B. EEG analysis based on time domain properties. Electroencephalogr. Clin. Neurophysiol. 1970, 29, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Petrantonakis, P.C.; Hadjileontiadis, L.J. Emotion Recognition from EEG Using Higher Order Crossings. IEEE Trans. Inf. Technol. Biomed. 2010, 14, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Petrantonakis, P.C.; Hadjileontiadis, L.J. Emotion Recognition from Brain Signals Using Hybrid Adaptive Filtering and Higher Order Crossings Analysis. IEEE Trans. Affect. Comput. 2010, 1, 81–97. [Google Scholar] [CrossRef]
- Veeranki, Y.R.; Ganapathy, N.; Swaminathan, R. Non-Parametric Classifiers Based Emotion Classification Using Electrodermal Activity and Modified Hjorth Features. In Public Health and Informatics; IOS Press: Amsterdam, The Netherlands, 2021. [Google Scholar] [CrossRef]
- Olofsson, J.K.; Nordin, S.; Sequeira, H.; Polich, J. Affective picture processing: An integrative review of ERP findings. Biol. Psychol. 2008, 77, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Bernat, E.; Bunce, S.; Shevrin, H. Event-related brain potentials differentiate positive and negative mood adjectives during both supraliminal and subliminal visual processing. Int. J. Psychophysiol. 2001, 42, 11–34. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.X. Analyzing Neural Time Series Data; The MIT Press: Cambridge, MA, USA, 2014. [Google Scholar] [CrossRef]
- Sorkhabi, M.M. Emotion detection from EEG signals with continuous wavelet analyzing. Am. J. Comput. Res. Repos 2014, 2, 66–70. [Google Scholar]
- Mohammadi, Z.; Frounchi, J.; Amiri, M. Wavelet-based emotion recognition system using EEG signal. Neural Comput. Appl. 2016, 28, 1985–1990. [Google Scholar] [CrossRef]
- Houssein, E.H.; Hamad, A.; Hassanien, A.E.; Fahmy, A.A. Epileptic detection based on whale optimization enhanced support vector machine. J. Inf. Optim. Sci. 2019, 40, 699–723. [Google Scholar] [CrossRef]
- Huang, J.R.; Fan, S.Z.; Abbod, M.; Jen, K.K.; Wu, J.F.; Shieh, J.S. Application of Multivariate Empirical Mode Decomposition and Sample Entropy in EEG Signals via Artificial Neural Networks for Interpreting Depth of Anesthesia. Entropy 2013, 15, 3325–3339. [Google Scholar] [CrossRef]
- Rehman, N.u.; Aftab, H. Multivariate Variational Mode Decomposition. IEEE Trans. Signal Process. 2019, 67, 6039–6052. [Google Scholar] [CrossRef]
- Padhmashree, V.; Bhattacharyya, A. Human emotion recognition based on time–frequency analysis of multivariate EEG signal. Knowl.-Based Syst. 2022, 238, 107867. [Google Scholar] [CrossRef]
- Zali-Vargahan, B.; Charmin, A.; Kalbkhani, H.; Barghandan, S. Deep time-frequency features and semi-supervised dimension reduction for subject-independent emotion recognition from multi-channel EEG signals. Biomed. Signal Process. Control 2023, 85, 104806. [Google Scholar] [CrossRef]
- Veeranki, Y.R.; McNaboe, R.; Posada-Quintero, H.F. EEG-Based Seizure Detection Using Variable-Frequency Complex Demodulation and Convolutional Neural Networks. Signals 2023, 4, 816–835. [Google Scholar] [CrossRef]
- Zorick, T.; Landers, J.; Leuchter, A.; Mandelkern, M.A. EEG multifractal analysis correlates with cognitive testing scores and clinical staging in mild cognitive impairment. J. Clin. Neurosci. 2020, 76, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, B.; Martinez-Rodrigo, A.; Alcaraz, R.; Fernandez-Caballero, A. A Review on Nonlinear Methods Using Electroencephalographic Recordings for Emotion Recognition. IEEE Trans. Affect. Comput. 2021, 12, 801–820. [Google Scholar] [CrossRef]
- Veeranki, Y.R.; Diaz, L.R.M.; Swaminathan, R.; Posada-Quintero, H.F. Nonlinear Signal Processing Methods for Automatic Emotion Recognition Using Electrodermal Activity. IEEE Sens. J. 2024, 24, 8079–8093. [Google Scholar] [CrossRef]
- Zhang, Y.; Duan, L.; Duan, M. A new feature extraction approach using improved symbolic aggregate approximation for machinery intelligent diagnosis. Measurement 2019, 133, 468–478. [Google Scholar] [CrossRef]
- Nardelli, M.; Greco, A.; Sebastiani, L.; Scilingo, E.P. ComEDA: A new tool for stress assessment based on electrodermal activity. Comput. Biol. Med. 2022, 150, 106144. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, M.; Zhang, J.; Chen, Z.; Huang, W.; Shen, G.; Zhang, M. Network Theory Based EHG Signal Analysis and its Application in Preterm Prediction. IEEE J. Biomed. Health Inform. 2022, 26, 2876–2887. [Google Scholar] [CrossRef] [PubMed]
- Lashkari, S.; Sheikhani, A.; Hashemi Golpayegani, M.R.; Moghimi, A.; Kobravi, H. Detection and Prediction of Absence Seizures Based on Nonlinear Analysis of the EEG in Wag/Rij Animal Model. Int. Clin. Neurosci. J. 2018, 5, 21–27. [Google Scholar] [CrossRef]
- Bruder, G.E.; Stewart, J.W.; McGrath, P.J. Right brain, left brain in depressive disorders: Clinical and theoretical implications of behavioral, electrophysiological and neuroimaging findings. Neurosci. Biobehav. Rev. 2017, 78, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Wager, T.D.; Phan, K.; Liberzon, I.; Taylor, S.F. Valence, gender, and lateralization of functional brain anatomy in emotion: A meta-analysis of findings from neuroimaging. NeuroImage 2003, 19, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Mouri, F.I.; Valderrama, C.E.; Camorlinga, S.G. Identifying relevant asymmetry features of EEG for emotion processing. Front. Psychol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.N.; Zhu, J.Y.; Lu, B.L. Differential entropy feature for EEG-based emotion classification. In Proceedings of the 2013 6th International IEEE/EMBS Conference on Neural Engineering (NER), San Diego, CA, USA, 6–8 November 2013; pp. 81–84. [Google Scholar] [CrossRef]
- Šverko, Z.; Vrankić, M.; Vlahinić, S.; Rogelj, P. Complex Pearson Correlation Coefficient for EEG Connectivity Analysis. Sensors 2022, 22, 1477. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.K.; Barrett, A.B.; Barnett, L. Granger Causality Analysis in Neuroscience and Neuroimaging. J. Neurosci. 2015, 35, 3293–3297. [Google Scholar] [CrossRef] [PubMed]
- Pullon, R.M.; Yan, L.; Sleigh, J.W.; Warnaby, C.E. Granger Causality of the Electroencephalogram Reveals Abrupt Global Loss of Cortical Information Flow during Propofol-induced Loss of Responsiveness. Anesthesiology 2020, 133, 774–786. [Google Scholar] [CrossRef]
- Zhong, P.; Wang, D.; Miao, C. EEG-Based Emotion Recognition Using Regularized Graph Neural Networks. IEEE Trans. Affect. Comput. 2022, 13, 1290–1301. [Google Scholar] [CrossRef]
- Song, T.; Zheng, W.; Song, P.; Cui, Z. EEG Emotion Recognition Using Dynamical Graph Convolutional Neural Networks. IEEE Trans. Affect. Comput. 2020, 11, 532–541. [Google Scholar] [CrossRef]
- Wang, M.; El-Fiqi, H.; Hu, J.; Abbass, H.A. Convolutional Neural Networks Using Dynamic Functional Connectivity for EEG-Based Person Identification in Diverse Human States. IEEE Trans. Inf. Forensics Secur. 2019, 14, 3259–3272. [Google Scholar] [CrossRef]
- Wang, X.H.; Zhang, T.; Xu, X.M.; Chen, L.; Xing, X.F.; Chen, C.L.P. EEG Emotion Recognition Using Dynamical Graph Convolutional Neural Networks and Broad Learning System. In Proceedings of the 2018 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Madrid, Spain, 3–6 December 2018; pp. 1240–1244. [Google Scholar] [CrossRef]
- Thammasan, N.; Fukui, K.i.; Numao, M. Application of deep belief networks in eeg-based dynamic music-emotion recognition. In Proceedings of the 2016 International Joint Conference on Neural Networks (IJCNN), Vancouver, BC, Canada, 24–29 July 2016; pp. 881–888. [Google Scholar] [CrossRef]
- Hu, J.; Wang, C.; Jia, Q.; Bu, Q.; Sutcliffe, R.; Feng, J. ScalingNet: Extracting features from raw EEG data for emotion recognition. Neurocomputing 2021, 463, 177–184. [Google Scholar] [CrossRef]
- Yin, Y.; Zheng, X.; Hu, B.; Zhang, Y.; Cui, X. EEG emotion recognition using fusion model of graph convolutional neural networks and LSTM. Appl. Soft Comput. 2021, 100, 106954. [Google Scholar] [CrossRef]
- Song, T.; Zheng, W.; Liu, S.; Zong, Y.; Cui, Z.; Li, Y. Graph-Embedded Convolutional Neural Network for Image-Based EEG Emotion Recognition. IEEE Trans. Emerg. Top. Comput. 2022, 10, 1399–1413. [Google Scholar] [CrossRef]
- Ackermann, P.; Kohlschein, C.; Bitsch, J.Á.; Wehrle, K.; Jeschke, S. EEG-based automatic emotion recognition: Feature extraction, selection and classification methods. In Proceedings of the 2016 IEEE 18th International Conference on e-Health Networking, Applications and Services (Healthcom), Munich, Germany, 14–16 September 2016; pp. 1–6. [Google Scholar] [CrossRef]
- Yin, Z.; Zhao, M.; Wang, Y.; Yang, J.; Zhang, J. Recognition of emotions using multimodal physiological signals and an ensemble deep learning model. Comput. Methods Programs Biomed. 2017, 140, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jin, Y.M.; Zheng, W.L.; Lu, B.L. Cross-Subject Emotion Recognition Using Deep Adaptation Networks. In Neural Information Processing; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 403–413. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, Q.; Qiu, M.; Wang, Y.; Chen, X. Emotion Recognition from Multi-Channel EEG through Parallel Convolutional Recurrent Neural Network. In Proceedings of the 2018 International Joint Conference on Neural Networks (IJCNN), Rio de Janeiro, Brazil, 8–13 July 2018; pp. 1–7. [Google Scholar] [CrossRef]
- Zhang, T.; Zheng, W.; Cui, Z.; Zong, Y.; Li, Y. Spatial–Temporal Recurrent Neural Network for Emotion Recognition. IEEE Trans. Cybern. 2018, 49, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, W.; Cui, Z.; Zhang, T.; Zong, Y. A Novel Neural Network Model Based on Cerebral Hemispheric Asymmetry for EEG Emotion Recognition. In Proceedings of the 27th International Joint Conference on Artificial Intelligence, IJCAI’18, Stockholm, Sweden, 13–19 July 2018; pp. 1561–1567. [Google Scholar]
- Li, Y.; Zheng, W.; Wang, L.; Zong, Y.; Cui, Z. From Regional to Global Brain: A Novel Hierarchical Spatial-Temporal Neural Network Model for EEG Emotion Recognition. IEEE Trans. Affect. Comput. 2019, 13, 568–578. [Google Scholar] [CrossRef]
- Duan, T.; Chauhan, M.; Shaikh, M.A.; Chu, J.; Srihari, S. Ultra Efficient Transfer Learning with Meta Update for Cross Subject EEG Classification. arXiv 2020, arXiv:2003.06113. [Google Scholar]
- Duan, T.; Shaikh, M.A.; Chauhan, M.; Chu, J.; Srihari, R.K.; Pathak, A.; Srihari, S.N. Meta Learn on Constrained Transfer Learning for Low Resource Cross Subject EEG Classification. IEEE Access 2020, 8, 224791–224802. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Zheng, W.; Zong, Y.; Qi, L.; Cui, Z.; Zhang, T.; Song, T. A Novel Bi-Hemispheric Discrepancy Model for EEG Emotion Recognition. IEEE Trans. Cogn. Dev. Syst. 2021, 13, 354–367. [Google Scholar] [CrossRef]
- Ding, K.M.; Kimura, T.; Fukui, K.i.; Numao, M. EEG emotion Enhancement using Task-specific Domain Adversarial Neural Network. In Proceedings of the 2021 International Joint Conference on Neural Networks (IJCNN), Shenzhen, China, 18–22 July 2021; pp. 1–8. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Ruan, Q.; Wang, S.; Wang, C. Cross-subject EEG emotion classification based on few-label adversarial domain adaption. Expert Syst. Appl. 2021, 185, 115581. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Zhao, L.; Li, B.; Hu, W.; Yu, J.; Zhang, Y.D. 3DCANN: A Spatio-Temporal Convolution Attention Neural Network for EEG Emotion Recognition. IEEE J. Biomed. Health Inform. 2022, 26, 5321–5331. [Google Scholar] [CrossRef]
- Liu, J.; Wu, H.; Zhang, L.; Zhao, Y. Spatial-Temporal Transformers for EEG Emotion Recognition. In Proceedings of the 6th International Conference on Advances in Artificial Intelligence, ICAAI ’22, Birmingham, UK, 21–23 October 2023; pp. 116–120. [Google Scholar] [CrossRef]
- Vogel, E.K.; Machizawa, M.G. Neural activity predicts individual differences in visual working memory capacity. Nature 2004, 428, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Dolcos, F.; Katsumi, Y.; Moore, M.; Berggren, N.; de Gelder, B.; Derakshan, N.; Hamm, A.O.; Koster, E.H.; Ladouceur, C.D.; Okon-Singer, H.; et al. Neural correlates of emotion-attention interactions: From perception, learning, and memory to social cognition, individual differences, and training interventions. Neurosci. Biobehav. Rev. 2020, 108, 559–601. [Google Scholar] [CrossRef]
- Hamann, S.; Canli, T. Individual differences in emotion processing. Curr. Opin. Neurobiol. 2004, 14, 233–238. [Google Scholar] [CrossRef]
- Lahnakoski, J.M.; Glerean, E.; Jääskeläinen, I.P.; Hyönä, J.; Hari, R.; Sams, M.; Nummenmaa, L. Synchronous brain activity across individuals underlies shared psychological perspectives. NeuroImage 2014, 100, 316–324. [Google Scholar] [CrossRef]
- Mohammad, Y.; Nishida, T. Using physiological signals to detect natural interactive behavior. Appl. Intell. 2010, 33, 79–92. [Google Scholar] [CrossRef]
- Lan, Z.; Sourina, O.; Wang, L.; Scherer, R.; Müller-Putz, G.R. Domain Adaptation Techniques for EEG-Based Emotion Recognition: A Comparative Study on Two Public Datasets. IEEE Trans. Cogn. Dev. Syst. 2019, 11, 85–94. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, S.Y.; Zheng, W.L.; Lu, B.L. WGAN Domain Adaptation for EEG-Based Emotion Recognition. In Neural Information Processing; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 275–286. [Google Scholar] [CrossRef]
- Ganin, Y.; Ustinova, E.; Ajakan, H.; Germain, P.; Larochelle, H.; Laviolette, F.; Marchand, M.; Lempitsky, V. Domain-Adversarial Training of Neural Networks. J. Mach. Learn. Res. 2016, 17, 1–35. [Google Scholar]
- Jiménez-Guarneros, M.; Gómez-Gil, P. Standardization-refinement domain adaptation method for cross-subject EEG-based classification in imagined speech recognition. Pattern Recognit. Lett. 2021, 141, 54–60. [Google Scholar] [CrossRef]
- Wan, Z.; Yang, R.; Huang, M.; Zeng, N.; Liu, X. A review on transfer learning in EEG signal analysis. Neurocomputing 2021, 421, 1–14. [Google Scholar] [CrossRef]
- Soleymani, M.; Pantic, M.; Pun, T. Multimodal Emotion Recognition in Response to Videos. IEEE Trans. Affect. Comput. 2012, 3, 211–223. [Google Scholar] [CrossRef]
- Neumeister, P.; Feldker, K.; Heitmann, C.Y.; Helmich, R.; Gathmann, B.; Becker, M.P.; Straube, T. Interpersonal violence in posttraumatic women: Brain networks triggered by trauma-related pictures. Soc. Cogn. Affect. Neurosci. 2016, 12, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Brohawn, K.H.; Offringa, R.; Pfaff, D.L.; Hughes, K.C.; Shin, L.M. The Neural Correlates of Emotional Memory in Posttraumatic Stress Disorder. Biol. Psychiatry 2010, 68, 1023–1030. [Google Scholar] [CrossRef]
- Hayes, J.P.; Hayes, S.M.; Mikedis, A.M. Quantitative meta-analysis of neural activity in posttraumatic stress disorder. Biol. Mood Anxiety Disord. 2012, 2, 9. [Google Scholar] [CrossRef]
- Stevens, J.S.; Jovanovic, T.; Fani, N.; Ely, T.D.; Glover, E.M.; Bradley, B.; Ressler, K.J. Disrupted amygdala-prefrontal functional connectivity in civilian women with posttraumatic stress disorder. J. Psychiatr. Res. 2013, 47, 1469–1478. [Google Scholar] [CrossRef]
- Gilboa, A.; Shalev, A.Y.; Laor, L.; Lester, H.; Louzoun, Y.; Chisin, R.; Bonne, O. Functional connectivity of the prefrontal cortex and the amygdala in posttraumatic stress disorder. Biol. Psychiatry 2004, 55, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Fonzo, G.A.; Simmons, A.N.; Thorp, S.R.; Norman, S.B.; Paulus, M.P.; Stein, M.B. Exaggerated and Disconnected Insular-Amygdalar Blood Oxygenation Level-Dependent Response to Threat-Related Emotional Faces in Women with Intimate-Partner Violence Posttraumatic Stress Disorder. Biol. Psychiatry 2010, 68, 433–441. [Google Scholar] [CrossRef]
- Liu, T.; Ke, J.; Qi, R.; Zhang, L.; Zhang, Z.; Xu, Q.; Zhong, Y.; Lu, G.; Chen, F. Altered functional connectivity of the amygdala and its subregions in typhoon-related post-traumatic stress disorder. Brain Behav. 2020, 11, e01952. [Google Scholar] [CrossRef]
- Milad, M.R.; Wright, C.I.; Orr, S.P.; Pitman, R.K.; Quirk, G.J.; Rauch, S.L. Recall of Fear Extinction in Humans Activates the Ventromedial Prefrontal Cortex and Hippocampus in Concert. Biol. Psychiatry 2007, 62, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Kalisch, R.; Korenfeld, E.; Stephan, K.E.; Weiskopf, N.; Seymour, B.; Dolan, R.J. Context-Dependent Human Extinction Memory Is Mediated by a Ventromedial Prefrontal and Hippocampal Network. J. Neurosci. 2006, 26, 9503–9511. [Google Scholar] [CrossRef] [PubMed]
- Lonsdorf, T.B.; Haaker, J.; Kalisch, R. Long-term expression of human contextual fear and extinction memories involves amygdala, hippocampus and ventromedial prefrontal cortex: A reinstatement study in two independent samples. Soc. Cogn. Affect. Neurosci. 2014, 9, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ramirez, S.; Pang, P.T.; Puryear, C.B.; Govindarajan, A.; Deisseroth, K.; Tonegawa, S. Optogenetic stimulation of a hippocampal engram activates fear memory recall. Nature 2012, 484, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, K.A.; Maren, S. Hippocampal Inactivation Disrupts Contextual Retrieval of Fear Memory after Extinction. J. Neurosci. 2001, 21, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Osipova, D.; Takashima, A.; Oostenveld, R.; Fernández, G.; Maris, E.; Jensen, O. Theta and Gamma Oscillations Predict Encoding and Retrieval of Declarative Memory. J. Neurosci. 2006, 26, 7523–7531. [Google Scholar] [CrossRef]
- Sederberg, P.B.; Kahana, M.J.; Howard, M.W.; Donner, E.J.; Madsen, J.R. Theta and Gamma Oscillations during Encoding Predict Subsequent Recall. J. Neurosci. 2003, 23, 10809–10814. [Google Scholar] [CrossRef]



| Emotion | Study | Activated Brain Region(s) | Stimulus Modality | Imaging Modality |
|---|---|---|---|---|
| Happiness | Habel et al., 2005 [30] | LEFT dorsolateral prefrontal cortex, LEFT anterior cingulate gyrus, RIGHT cingulate gyrus, LEFT posterior cingulate gyrus, LEFT middle temporal gyrus, LEFT parietal cortex, LEFT angular gyrus, RIGHT paracentral lobule, LEFT parahippocampal gyrus, LEFT amygdala | Visual (static images) | fMRI |
| Killgore and Yurgelun- Todd, 2004 [31] | LEFT/RIGHT amygdala, LEFT/RIGHT anterior cingulate | Visual (static images) | fMRI | |
| Esslen et al., 2004 [32] | LEFT/RIGHT frontal cortex, LEFT/RIGHT ventromedial frontal, LEFT/RIGHT temporal cortex, RIGHT parietal area, anterior cingulate cortex, RIGHT frontal areas | Visual (static images) | EEG (LORETA) | |
| Gorno-Tempini et al., 2001 [33] | RIGHT precentral sulcus, RIGHT middle and inferior frontal gyri, RIGHT posterior fusiform gyrus, RIGHT anterior insula, occipito-temporal cortex, prefrontal cortex, amygdala, basal ganglia, LEFT/RIGHT orbitofrontal cortex | Visual (static images) | fMRI | |
| Kesler-West et al., 2001 [34] | LEFT/RIGHT fusiform gyri, LEFT/RIGHT amygdala, LEFT/RIGHT entorhinal cortices, RIGHT superior temporal sulcus, RIGHT inferior occipital gyrus, LEFT/RIGHT inferior frontal gyri, RIGHT angular gyrus, LEFT/RIGHT lingual gyri, medial frontal/cingulate sulcus | Visual (static images) | fMRI | |
| van de Riet et al., 2009 [35] | calcarine sulcus, anterior cingulate gyrus, cerebellum, RIGHT fusiform gyrus, parietal lobe, primary motor cortex, premotor cortex, primary somatosensory cortex, olfactory sulcus, insula | Visual (static images of faces and bodies) | fMRI | |
| Fitzgerald et al., 2006 [36] | LEFT amygdala, LEFT lingual gyrus | Visual (static images of faces) | fMRI | |
| Sadness | Habel et al., 2005 [30] | LEFT dorsolateral prefrontal cortex, LEFT orbitofrontal cortex, LEFT superior frontal gyrus, LEFT middle temporal gyrus, LEFT superior temporal gyrus, LEFT precuneus, LEFT parahippocampal gyrus, LEFT amygdala-hippocampal area, LEFT putamen, RIGHT fasciculus occipito-frontalis, LEFT insula | Visual (static images of faces) | fMRI |
| Killgore and Yurgelun-Todd, 2004 [31] | LEFT/RIGHT amygdala, LEFT/RIGHT anterior cingulate | Visual (static images of faces) | fMRI | |
| Esslen et al., 2004 [32] | LEFT postcentral area, RIGHT prefrontal cortex, LEFT/RIGHT ventromedial frontal, LEFT/RIGHT orbitofrontal, RIGHT prefrontal cortex, LEFT/RIGHT temporal cortex, RIGHT occipital cortex, LEFT superior frontal cortex, RIGHT frontal cortex, LEFT/RIGHT posterior cingulate cortex, LEFT/RIGHT occipital lobe | Visual (static images of faces) | EEG (LORETA) | |
| Kesler-West et al., 2001 [34] | LEFT/RIGHT fusiform gyri, LEFT/RIGHT amygdala, LEFT/RIGHT entorhinal cortices, RIGHT superior temporal sulcus, RIGHT inferior occipital gyrus, LEFT/RIGHT inferior frontal gyri, RIGHT angular gyrus, LEFT/RIGHT lingual gyri | Visual (static images of faces) | fMRI | |
| Fitzgerald et al., 2006 [36] | LEFT amygdala, RIGHT middle temporal gyrus, LEFT middle frontal gyrus, RIGHT inferior frontal gyrus | Visual (static images of faces) | fMRI | |
| Fear | Esslen et al., 2004 [32] | LEFT temporal area, RIGHT frontal pole, RIGHT temporal cortex | Visual (static images of faces) | EEG (LORETA) |
| Kesler-West et al., 2001 [34] | LEFT/RIGHT fusiform gyri, LEFT/RIGHT amygdala, LEFT/RIGHT entorhinal cortices, RIGHT superior temporal sulcus, RIGHT inferior occipital gyrus, LEFT/RIGHT inferior frontal gyri, RIGHT angular gyrus, LEFT/RIGHT lingual gyri | Visual (static images of faces) | fMRI | |
| van de Riet et al., 2009 [35] | calcarine sulcus, anterior cingulate gyrus, cerebellum, LEFT/RIGHT amygdala, LEFT/RIGHT fusiform gyri, RIGHT inferior occipital gyrus, RIGHT superior temporal sulcus, superior colliculus | Visual (static images of faces and bodies) | fMRI | |
| Fitzgerald et al., 2006 [36] | LEFT amygdala, LEFT inferior frontal gyrus | Visual (static images of faces) | fMRI | |
| Lange et al., 2003 [37] | LEFT amygdala, LEFT hippocampus, LEFT putamen, LEFT ventrofrontal gyrus, LEFT lingual gyrus, LEFT/RIGHT cerebellum | Visual (static images of faces) | fMRI | |
| Liddell et al., 2005 [38] | LEFT/RIGHT Amygdala, LEFT pulvinar, LEFT superior colliculus, LEFT locus coeruleus, LEFT/RIGHT anterior cingulate, LEFT inferior frontal gyrus, RIGHT middle frontal gyrus, LEFT cingulate gyrus, LEFT medial frontal, LEFT superior/middle frontal gyri, LEFT caudate, LEFT insula, LEFT postcentral gyrus, LEFT superior temporal gyrus, LEFT middle temporal gyrus, LEFT inferior temporal gyrus | Visual (static images of faces) | fMRI | |
| Phillips et al., 2004 [39] | LEFT/RIGHT posterior cingulate gyrus, LEFT putamen, RIGHT inferior parietal lobule, RIGHT amygdala/hippocampus, RIGHT anterior cingulate gyrus, RIGHT lingual gyrus, RIGHT medial frontal gyrus, RIGHT precuneus, RIGHT cerebellum, LEFT inferior frontal gyrus, LEFT/RIGHT superior temporal gyrus, RIGHT middle temporal gyrus | Visual (static images of faces) | fMRI | |
| Thielscher & Pessoa, 2007 [40] | LEFT lingual gyrus, RIGHT precuneus, LEFT fusiform gyrus, RIGHT entorhinal cortex, LEFT/RIGHT anterior insula/inferior frontal gyrus, RIGHT inferior frontal sulcus, RIGHT orbital gyrus, RIGHT thalamus, LEFT caudate | Visual (static images of faces) | fMRI | |
| Williams et al., 2005 [41] | LEFT amygdala, LEFT hippocampus, LEFT/RIGHT medial prefrontal cortex, RIGHT anterior cingulate, RIGHT lateral prefrontal cortex, RIGHT occipital gyrus, RIGHT fusiform gyrus, LEFT superior temporal gyrus, LEFT/RIGHT postcentral gyrus, LEFT putamen, LEFT thalamus | Visual (static images of faces) | fMRI | |
| Anger | Esslen et al., 2004 [32] | RIGHT frontal lobe, MEDIAL superior frontal gyrus, RIGHT prefrontal cortex | Visual (static images of faces) | EEG (LORETA) |
| Kesler-West et al., 2001 [34] | LEFT/RIGHT fusiform gyri, LEFT/RIGHT amygdala, LEFT/RIGHT entorhinal cortices, RIGHT superior temporal sulcus, RIGHT inferior occipital gyrus, LEFT/RIGHT inferior frontal gyri, RIGHT angular gyrus, LEFT/RIGHT lingual gyri, LEFT precentral gyrus, medial superior frontal gyrus, RIGHT lateral occipital gyrus | Visual (static images of faces) | fMRI | |
| Fitzgerald et al., 2006 [36] | LEFT amygdala, LEFT inferior temporal gyrus, LEFT fusiform gyrus, LEFT middle frontal gyrus | Visual (static images of faces) | fMRI | |
| Williams et al., 2005 [41] | RIGHT amygdala, LEFT/RIGHT medial prefrontal cortex, RIGHT anterior cingulate, LEFT lateral prefrontal cortex, LEFT occipital gyrus, LEFT fusiform gyrus, RIGHT superior temporal gyrus, RIGHT putamen, RIGHT thalamus | Visual (static images of faces) | fMRI | |
| Disgust | Esslen et al., 2004 [32] | RIGHT frontal cortex, LEFT/RIGHT frontal lobes, anterior cingulate cortex, premotor cortex | Visual (static images of faces) | EEG (LORETA) |
| Gorno-Tempini et al., 2001 [33] | RIGHT precentral sulcus, RIGHT middle and inferior frontal gyri, RIGHT posterior fusiform gyrus, RIGHT anterior insula, occipito-temporal cortex, prefrontal cortex, LEFT amygdala, basal ganglia, RIGHT striatum, RIGHT head of caudate nucleus, RIGHT thalamus | Visual (static images of faces) | fMRI | |
| Fitzgerald et al., 2006 [36] | LEFT amygdala, LEFT inferior occipital gyrus, LEFT middle occipital gyrus, LEFT medial frontal gyrus, LEFT lingual gyrus, LEFT inferior frontal gyrus, LEFT medial frontal gyrus, LEFT inferior frontal gyrus | Visual (static images of faces) | fMRI | |
| Phillips et al., 2004 [39] | LEFT middle temporal gyrus, LEFT/RIGHT posterior cingulate gyrus, RIGHT precuneus, LEFT/RIGHT cerebellum, LEFT postcentral gyrus, LEFT inferior parietal lobule, RIGHT insula, RIGHT anterior cingulate gyrus, RIGHT lingual gyrus, RIGHT superior temporal gyrus, LEFT middle occipital gyrus | Visual (static images of faces) | fMRI | |
| Thielscher & Pessoa, 2007 [40] | RIGHT middle occipital gyrus, LEFT intraparietal sulcus, RIGHT middle temporal gyrus | Visual (static images of faces) | fMRI | |
| Williams et al., 2005 [41] | LEFT/RIGHT insula, LEFT/RIGHT amygdala-hippocampal complex, RIGHT hippocampal gyrus, RIGHT medial prefrontal cortex, LEFT/RIGHT anterior cingulate, LEFT/RIGHT lateral prefrontal cortex, RIGHT occipital gyrus, LEFT/RIGHT superior temporal gyrus | Visual (static images of faces) | fMRI |
| Year | Dataset | Stimuli | Emotion Model | Modalities Type | EEG Stats (Channels, Sampling Frequency, etc.) | Participants | EEG Device |
|---|---|---|---|---|---|---|---|
| 2012 | DEAP [149] | 40 Video clips— Music Video | Valence, Arousal, Dominance, Liking, Familiarity | EEG, EOG, EMG, Galvanic Skin Response, Respiration, Temperature, Plethysmograph. | 32 EEG channels, 12 peripheral channels, Rae Signals—512 Hz, Preprocessed data: 128 Hz) | 32 (16 male, 16 female), Race: Mixed | Biosemi ActiveTwo |
| 2012 | MAHNOB HCI [150] | 20 Video Clips | Dimensional: Arousal, Valence, Control, Predictability. Emotion | EEG, EXG, GSR, Respiration, Temperature, Eye Gaze | 32 Channels | 27 (11male, 16 female), Race: Mixed | Biosemi ActiveTwo |
| 2015 | SEED [151] | 15 Video Clips | Positive, Negative, Neutral | EEG, EMG, EOG | 62 Channels | 15 (7 male, 8 female), Race: Chinese | ESI NeuroScan |
| 2018 | Dreamer [152] | 18 Video Clips | Arousal, Valence, Dominance | EEG, ECG | 14 Channels | 25 (14 male, 11 female), Race: Mixed | Emotiv EPOC |
| 2019 | SEED-IV [153] | 72 Video Clips | Happy, Sad, Fear and Neutral | EEG, Physiological Signals | 62 Channels | 15 (7 male, 8 female), Race: Chinese | ESI Neuroscan |
| 2019 | MPED [154] | 28 Video Clips | Joy, Funny, Anger, Fear, Disgust, sad, neutral | EEG, ECG, GSR, Respiration | 62 Channels | 23 (10 male, 13 female), Race: Chinese | ESI NeuroScan |
| 2021 | AMIGOS [155] | 16 Videos | Big-Five Personality traits, Emotions; Valence Arousal, Dominance, Liking, Familiarity | EEG, ECG, GSR, Audio Visual, Depth | 14 Channels, 128 Hz | 40 (27 male, 13 female) for short videos; 37 for Long Videos, Race: Mixed | Emotiv EPOC NEuroheadset |
| 2022 | SEED-V [156] | 15 Videos | Happy, Sad, Fear, Neutral, Disgust | EEG, Physiological Signals | 62 Channels | 20 (10 male, 10 female), Race: Chinese | ESI NeuroScan |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, R.Y.; Lew, W.-C.L.; Ang, K.K. Review of EEG Affective Recognition with a Neuroscience Perspective. Brain Sci. 2024, 14, 364. https://doi.org/10.3390/brainsci14040364
Lim RY, Lew W-CL, Ang KK. Review of EEG Affective Recognition with a Neuroscience Perspective. Brain Sciences. 2024; 14(4):364. https://doi.org/10.3390/brainsci14040364
Chicago/Turabian StyleLim, Rosary Yuting, Wai-Cheong Lincoln Lew, and Kai Keng Ang. 2024. "Review of EEG Affective Recognition with a Neuroscience Perspective" Brain Sciences 14, no. 4: 364. https://doi.org/10.3390/brainsci14040364
APA StyleLim, R. Y., Lew, W.-C. L., & Ang, K. K. (2024). Review of EEG Affective Recognition with a Neuroscience Perspective. Brain Sciences, 14(4), 364. https://doi.org/10.3390/brainsci14040364