
Complete Neuron Reconstruction Based on Branch Confidence

Abstract
:1. Introduction
2. Materials and Methods
2.1. Artifacts of Neuron Reconstruction
2.2. Common Features of Neuronal Branch
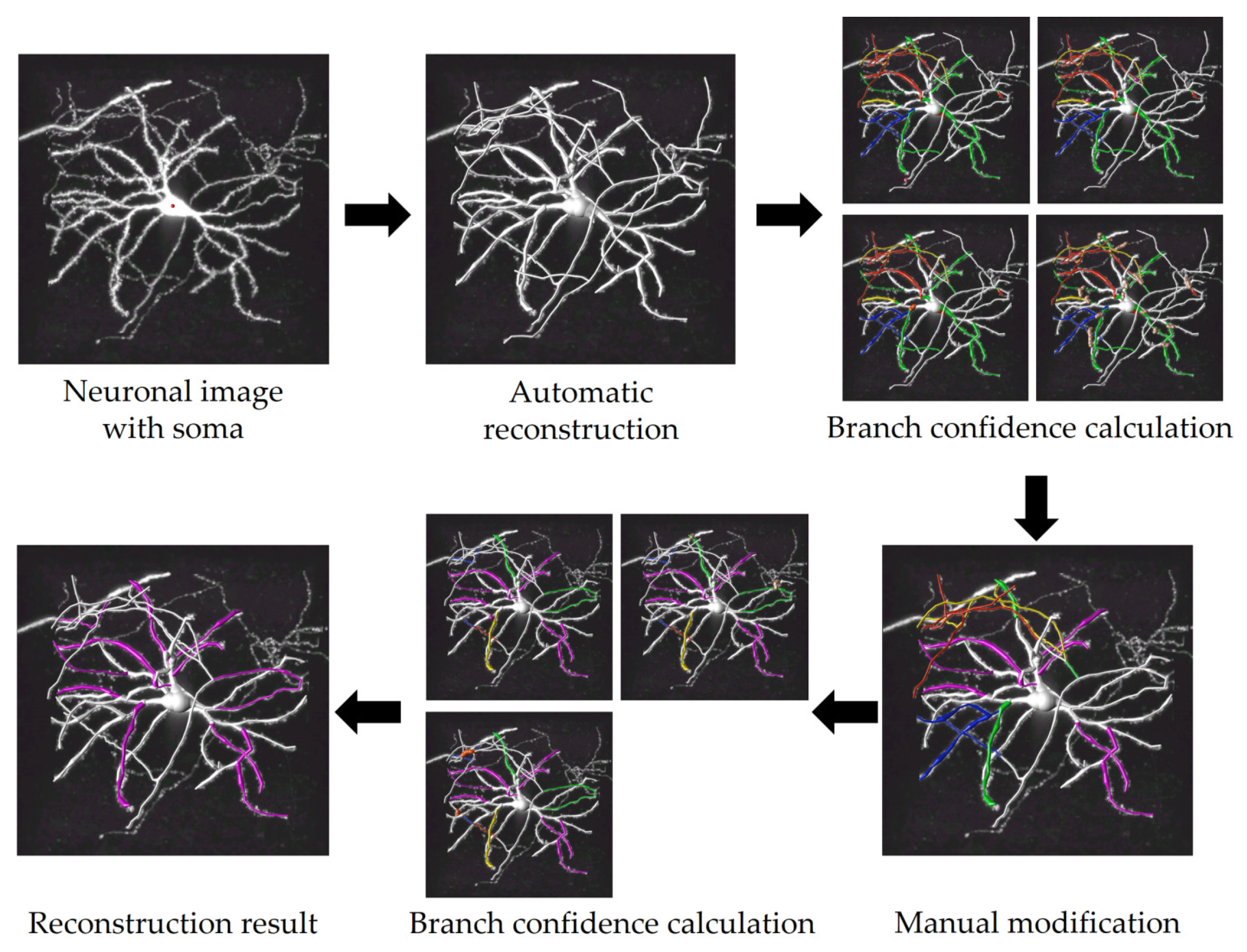
2.3. Model Designation
2.4. Data Preprocessing and Skeleton Branch Extraction
2.5. Branch Confidence Calculation
3. Results
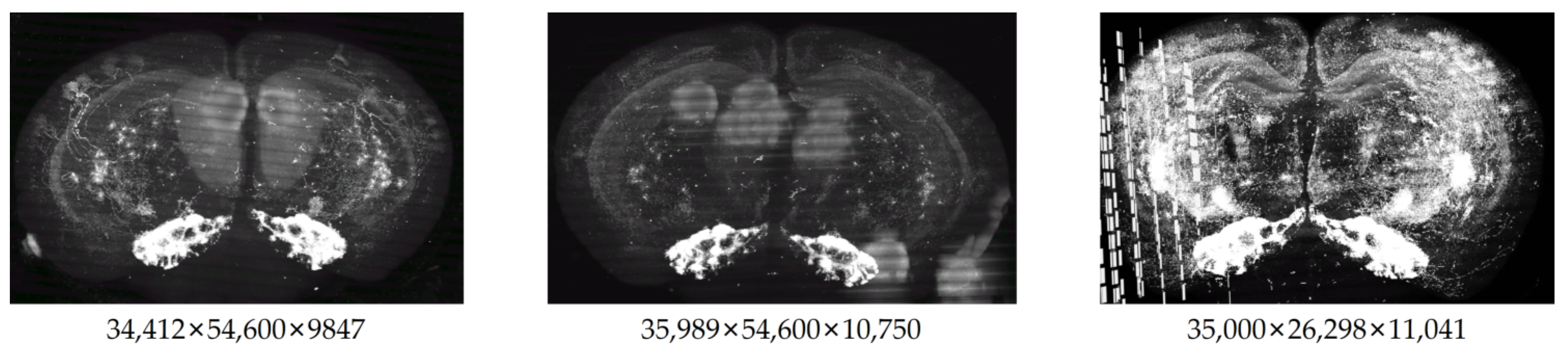
3.1. Datasets and Platform
3.2. Reconstruction Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Petrella, J.R.; Coleman, R.E.; Doraiswamy, P.M. Neuroimaging and early diagnosis of Alzheimer disease: A look to the future. Radiology 2003, 226, 315–336. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, A.; De Stefano, N. Clinical use of brain volumetry. J. Magn. Reson. Imaging 2013, 37, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.A.; Firestein, B.L. The dendritic tree and brain disorders. Mol. Cell. Neurosci. 2012, 50, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Xu, D.; Yuan, J.; Li, X.; Guo, C.; Peng, J.; Li, Y.; Schwarz, L.A.; Li, A.; Hu, B.; et al. High-throughput dual-colour precision imaging for brain-wide connectome with cytoarchitectonic landmarks at the cellular level. Nat. Commun. 2016, 7, 12142. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xiong, H.; Liu, Y.; Yang, T.; Li, A.; Huang, F.; Yin, F.; Su, L.; Liu, L.; Li, N.; et al. Chemical sectioning fluorescence tomography: High-throughput, high-contrast, multicolor, whole-brain imaging at subcellular resolution. Cell Rep. 2021, 34, 108709. [Google Scholar] [CrossRef] [PubMed]
- Dodt, H.U.; Leischner, U.; Schierloh, A.; Jährling, N.; Mauch, C.P.; Deininger, K.; Deussing, J.M.; Eder, M.; Zieglgänsberger, W.; Becker, K. Ultramicroscopy: Three-dimensional visualization of neuronal networks in the whole mouse brain. Nat. Methods 2007, 4, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, M.B.; Orger, M.B.; Robson, D.N.; Li, J.M.; Keller, P.J. Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nat. Methods 2013, 10, 413–420. [Google Scholar] [CrossRef]
- Glaser, J.R.; Glaser, E.M. Neuron imaging with Neurolucida—A PC-based system for image combining microscopy. Comput. Med. Imaging Graph. 1990, 14, 307–317. [Google Scholar] [CrossRef]
- Peng, H.; Bria, A.; Zhou, Z.; Iannello, G.; Long, F. Extensible visualization and analysis for multidimensional images using Vaa3D. Nat. Protoc. 2014, 9, 193–208. [Google Scholar] [CrossRef]
- Winnubst, J.; Bas, E.; Ferreira, T.A.; Wu, Z.; Economo, M.N.; Edson, P.; Arthur, B.J.; Bruns, C.; Rokicki, K.; Schauder, D.; et al. Reconstruction of 1,000 projection neurons reveals new cell types and organization of long-range connectivity in the mouse brain. Cell 2019, 179, 268–281. [Google Scholar] [CrossRef]
- Economo, M.N.; Clack, N.G.; Lavis, L.D.; Gerfen, C.R.; Svoboda, K.; Myers, E.W.; Chandrashekar, J. A platform for brain-wide imaging and reconstruction of individual neurons. eLife 2016, 5, e10566. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Meijering, E.; Ascoli, G.A. From diadem to bigneuron. Neuroinformatics 2015, 13, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Peng, H. APP2: Automatic tracing of 3D neuron morphology based on hierarchical pruning of a gray-weighted image distance-tree. Bioinformatics 2013, 29, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Ming, X.; Li, A.; Wu, J.; Yan, C.; Ding, W.; Gong, H.; Zeng, S.; Liu, Q. Rapid reconstruction of 3D neuronal morphology from light microscopy images with augmented rayburst sampling. PLoS ONE 2013, 8, e84557. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.W.; Lee, Y.C.; Pradana, H.; Zhou, Z.; Peng, H. Ensemble neuron tracer for 3D neuron reconstruction. Neuroinformatics 2017, 15, 185–198. [Google Scholar] [CrossRef]
- Li, Q.; Shen, L. 3D neuron reconstruction in tangled neuronal image with deep networks. IEEE Trans. Med. Imaging 2019, 39, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, X.; Xiong, Z.; Liu, D.; Zeng, J.; Xie, C.; Zhang, Y.; Zha, Z.J.; Bi, G.; Wu, F. Neuronal population reconstruction from ultra-scale optical microscopy images via progressive learning. IEEE Trans. Med. Imaging 2020, 39, 4034–4046. [Google Scholar] [CrossRef]
- Chen, W.; Liu, M.; Du, H.; Radojević, M.; Wang, Y.; Meijering, E. Deep-learning-based automated neuron reconstruction from 3D microscopy images using synthetic training images. IEEE Trans. Med. Imaging 2021, 41, 1031–1042. [Google Scholar] [CrossRef]
- Radojević, M.; Meijering, E. Automated neuron tracing using probability hypothesis density filtering. Bioinformatics 2017, 33, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Athey, T.L.; Tward, D.J.; Mueller, U.; Vogelstein, J.T.; Miller, M.I. Hidden Markov modeling for maximum probability neuron reconstruction. Commun. Biol. 2022, 5, 388. [Google Scholar] [CrossRef]
- Li, S.; Quan, T.; Zhou, H.; Yin, F.; Li, A.; Fu, L.; Luo, Q.; Gong, H.; Zeng, S. Identifying weak signals in inhomogeneous neuronal images for large-scale tracing of sparsely distributed neurites. Neuroinformatics 2019, 17, 497–514. [Google Scholar] [CrossRef]
- Liu, Y.; Zhong, Y.; Zhao, X.; Liu, L.; Ding, L.; Peng, H. Tracing weak neuron fibers. Bioinformatics 2023, 39, btac816. [Google Scholar] [CrossRef] [PubMed]
- Palágyi, K.; Kuba, A. A parallel 3D 12-subiteration thinning algorithm. Graph. Model. Image Process. 1999, 61, 199–221. [Google Scholar] [CrossRef]
- Radojević, M.; Meijering, E. Automated neuron reconstruction from 3D fluorescence microscopy images using sequential Monte Carlo estimation. Neuroinformatics 2019, 17, 423–442. [Google Scholar] [CrossRef]
- Peng, H.; Hawrylycz, M.; Roskams, J.; Hill, S.; Spruston, N.; Meijering, E.; Ascoli, G.A. BigNeuron: Large-scale 3D neuron reconstruction from optical microscopy images. Neuron 2015, 87, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Ruan, Z.; Long, F.; Simpson, J.H.; Myers, E.W. V3D enables real-time 3D visualization and quantitative analysis of large-scale biological image data sets. Nat. Biotechnol. 2010, 28, 348–353. [Google Scholar] [CrossRef]
- Quan, T.; Zhou, H.; Li, J.; Li, S.; Li, A.; Li, Y.; Lv, X.; Luo, Q.; Gong, H.; Zeng, S. NeuroGPS-Tree: Automatic reconstruction of large-scale neuronal populations with dense neurites. Nat. Methods 2016, 13, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Ruan, Z.; Atasoy, D.; Sternson, S. Automatic reconstruction of 3D neuron structures using a graph-augmented deformable model. Bioinformatics 2010, 26, i38–i46. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, D.; Song, Y.; Peng, H.; Cai, W. Automated 3-D neuron tracing with precise branch erasing and confidence controlled back tracking. IEEE Trans. Med. Imaging 2018, 37, 2441–2452. [Google Scholar] [CrossRef]
- Liu, M.; Luo, H.; Tan, Y.; Wang, X.; Chen, W. Improved V-Net Based Image Segmentation for 3D Neuron Reconstruction. In Proceedings of the 2018 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Madrid, Spain, 3–6 December 2018; IEEE: Piscataway, NJ, USA, 2018; pp. 443–448. [Google Scholar]
- Yang, B.; Liu, M.; Wang, Y.; Zhang, K.; Meijering, E. Structure-guided segmentation for 3D neuron reconstruction. IEEE Trans. Med. Imaging 2021, 41, 903–914. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Method | Time (s) | Precision | Recall | F1-Score | ESA | DSA | PDS |
|---|---|---|---|---|---|---|---|---|
| Whole-Brain | Manual | 1021.656 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 |
| Automatic | 8.058 | 0.708 | 0.919 | 0.764 | 4.175 | 12.253 | 0.187 | |
| Automatic+Manual | 1028.262 | 0.929 | 0.937 | 0.928 | 0.819 | 5.553 | 0.054 | |
| MouseLight | 1348.945 | 0.802 | 0.773 | 0.765 | 1.915 | 6.308 | 0.184 | |
| Ours | 704.635 | 0.931 | 0.930 | 0.930 | 0.753 | 4.948 | 0.051 | |
| BigNeuron | Manual | 743.366 | 0.967 | 0.907 | 0.936 | 0.934 | 4.411 | 0.049 |
| Automatic | 3.560 | 0.968 | 0.346 | 0.499 | 4.918 | 8.290 | 0.313 | |
| Automatic+Manual | 669.420 | 0.950 | 0.850 | 0.896 | 1.066 | 4.104 | 0.080 | |
| MouseLight | 332.586 | 0.834 | 0.561 | 0.641 | 2.637 | 5.687 | 0.268 | |
| Ours | 620.583 | 0.969 | 0.883 | 0.924 | 0.957 | 4.074 | 0.057 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Wang, Y. Complete Neuron Reconstruction Based on Branch Confidence. Brain Sci. 2024, 14, 396. https://doi.org/10.3390/brainsci14040396
Zeng Y, Wang Y. Complete Neuron Reconstruction Based on Branch Confidence. Brain Sciences. 2024; 14(4):396. https://doi.org/10.3390/brainsci14040396
Chicago/Turabian StyleZeng, Ying, and Yimin Wang. 2024. "Complete Neuron Reconstruction Based on Branch Confidence" Brain Sciences 14, no. 4: 396. https://doi.org/10.3390/brainsci14040396
APA StyleZeng, Y., & Wang, Y. (2024). Complete Neuron Reconstruction Based on Branch Confidence. Brain Sciences, 14(4), 396. https://doi.org/10.3390/brainsci14040396