Temporal Course of Interference Control from Early to Late Young Adulthood: An ERP Study


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Psychological Testing Battery
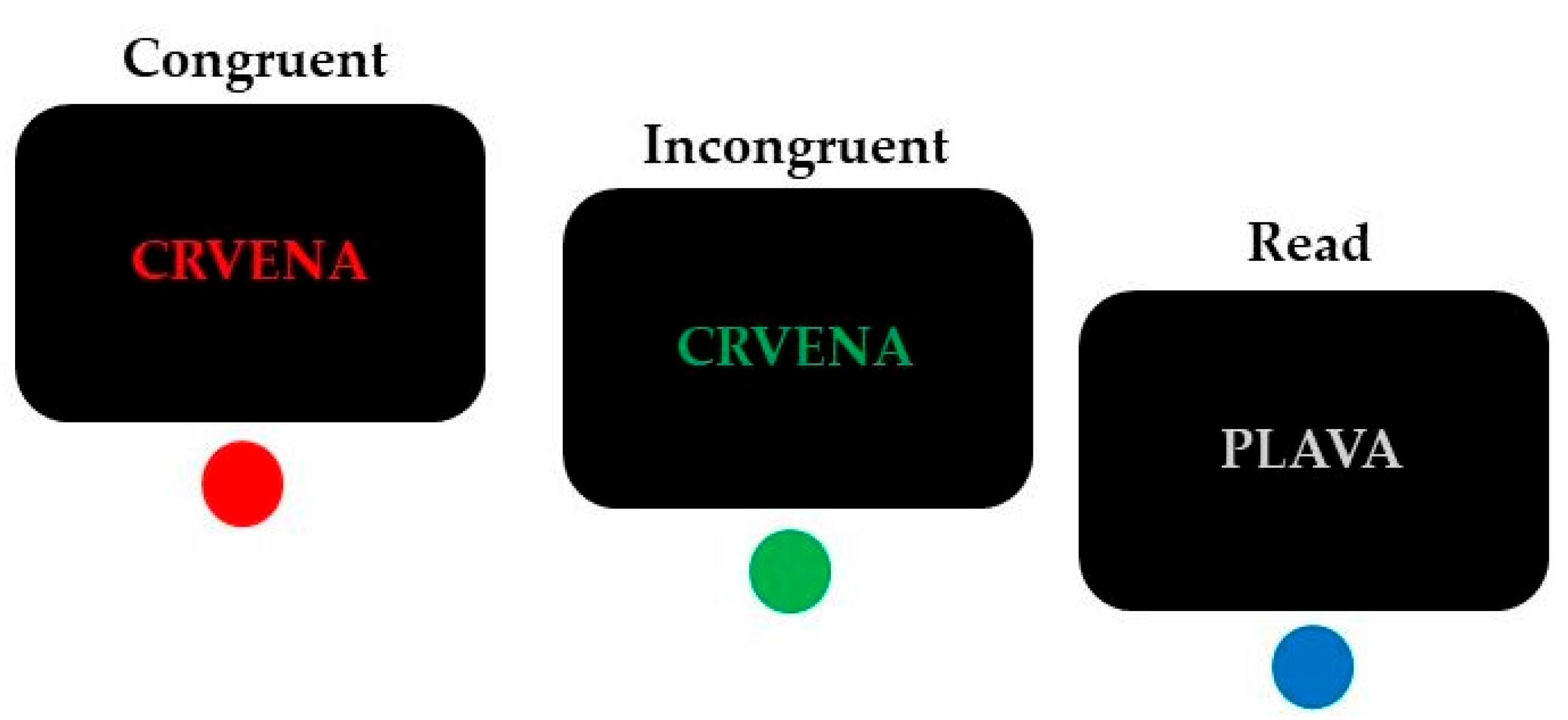
2.3. The Task
2.4. Procedure
2.5. Data Recording and Analysis
2.6. Statistical Analyses
3. Results
3.1. Psychological Testing Battery
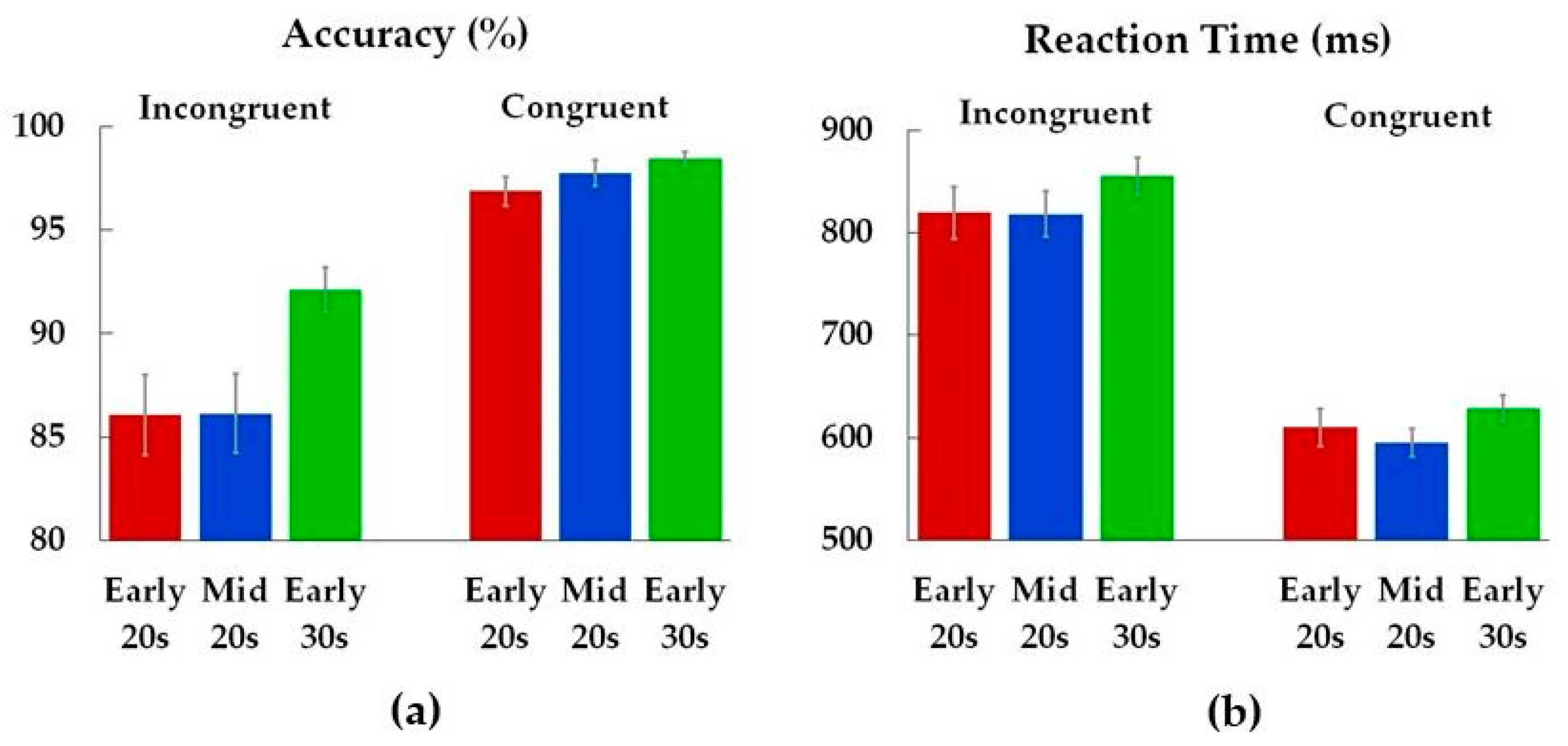
3.2. Performance
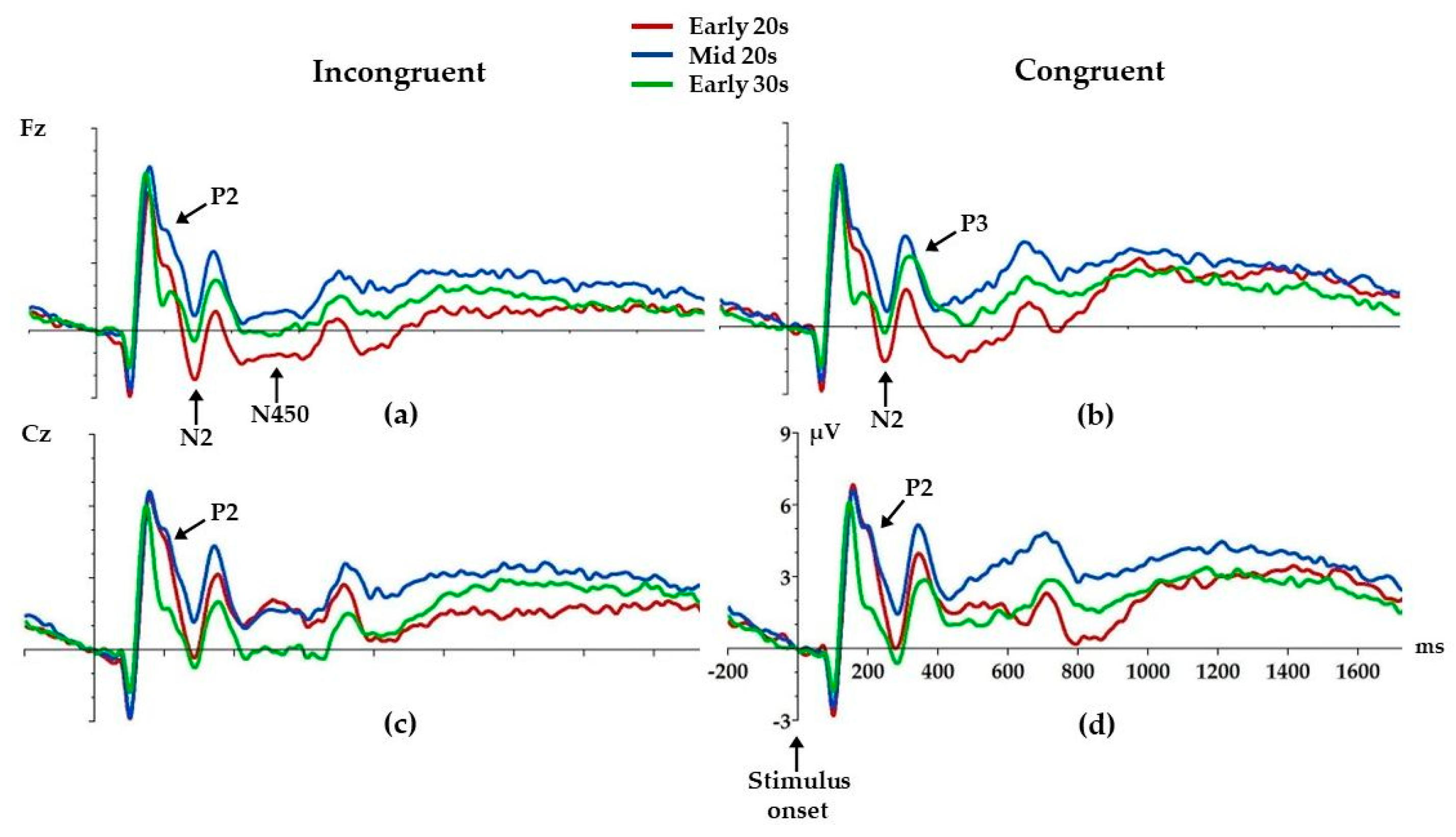
3.3. ERP Analysis
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- King, P.M.; Kitchener, K.S. Cognitive development in the emerging adult: The emergence of complex cognitive skills. In The Oxford Handbook of Emerging Adulthood; Arnett, J.J., Ed.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Ferguson, H.J.; Brunsdon, V.E.A.; Bradford, E.E.F. The developmental trajectories of executive function from adolescence to old age. Sci. Rep. 2021, 11, 1382. [Google Scholar] [CrossRef] [PubMed]
- Fjell, A.M.; Walhovd, K.B.; Brown, T.T.; Kuperman, J.M.; Chung, Y.; Hagler, D.J., Jr.; Venkatraman, V.; Roddey, J.C.; Erhart, M.; McCabe, C.; et al. Multimodal imaging of the self-regulating developing brain. Proc. Natl. Acad. Sci. USA 2012, 109, 19620–19625. [Google Scholar] [CrossRef]
- Buzzell, G.A.; Richards, J.E.; White, L.K.; Barker, T.V.; Pine, D.S.; Fox, N.A. Development of the error-monitoring system from ages 9-35: Unique insight provided by MRI-constrained source localization of EEG. Neuroimage 2017, 157, 13–26. [Google Scholar] [CrossRef]
- Mills, K.L.; Goddings, A.L.; Clasen, L.S.; Giedd, J.N.; Blakemore, S.J. The Developmental Mismatch in Structural Brain Maturation during Adolescence. Dev. Neurosci. 2014, 36, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Gogtay, N.; Giedd, J.N.; Lusk, L.; Hayashi, K.M.; Greenstein, D.; Vaituzis, A.C.; Nugent, T.F., 3rd; Herman, D.H.; Clasen, L.S.; Toga, A.W.; et al. Dynamic mapping of human cortical development during childhood through early adulthood. Proc. Nacl. Acad. Sci. USA 2004, 101, 8174–8179. [Google Scholar] [CrossRef] [PubMed]
- Knežević, M.; Veroude, K.; Jolles, J.; Krabbendam, L. Neural Correlates of Performance Monitoring During the Transition to Young Adulthood. Mind Brain Educ. 2016, 10, 81–90. [Google Scholar] [CrossRef]
- Knežević, M.; Marinković, K. Neurodynamic correlates of response inhibition from emerging to mid adulthood. Cogn. Dev. 2017, 43, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Knežević, M. Error processing in young adulthood: Age-related differences in electrophysiology and behavioral performance. Neuropsychology 2024, 38, 259–267. [Google Scholar] [CrossRef]
- Knežević, M. Differences in the Efficiency of Cognitive Control across Young Adulthood: An ERP Perspective. Brain Sci. 2024, 14, 347. [Google Scholar] [CrossRef]
- Rubia, K.; Smith, A.B.; Woolley, J.; Nosarti, C.; Heyman, I.; Taylor, E.; Brammer, M. Progressive increase of frontostriatal brain activation from childhood to adulthood during event-related tasks of cognitive control. Hum. Brain Mapp. 2006, 27, 973–993. [Google Scholar] [CrossRef]
- Theodoraki, T.E.; McGeown, S.P.; Rhodes, S.M.; MacPherson, S.E. Developmental changes in executive functions during adolescence: A study of inhibition, shifting, and working memory. Br. J. Dev. Psychol. 2020, 38, 74–89. [Google Scholar] [CrossRef] [PubMed]
- Korzeniowski, C.; Ison, M.S.; Difabio de Anglat, H. A Summary of the Developmental Trajectory of Executive Functions from Birth to Adulthood. In Psychiatry and Neuroscience Update; Gargiulo, P.Á., Mesones Arroyo, H.L., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Scarpina, F.; Tagini, S. The Stroop Color and Word Test. Front. Psychol. 2017, 8, 557. [Google Scholar] [CrossRef]
- Friedman, N.P.; Miyake, A. Unity and diversity of executive functions: Individual differences as a window on cognitive structure. Cortex 2017, 86, 186–204. [Google Scholar] [CrossRef] [PubMed]
- Floden, D.; Vallesi, A.; Stuss, D.T. Task Context and Frontal Lobe Activation in the Stroop Task. J. Cogn. Neurosci. 2011, 23, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Lei, X.; Ding, C.; Li, H.; Chen, A. The neural mechanisms of semantic and response conflicts: An fMRI study of practice-related effects in the Stroop task. Neuroimage 2013, 66, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.L.; Thompson, P.M. Typical and atypical brain development: A review of neuroimaging studies. Dialogues Clin. Neurosci. 2013, 15, 359–384. [Google Scholar] [CrossRef] [PubMed]
- Luna, B.; Velanova, K.; Geier, C.F. Methodological approaches in developmental neuroimaging studies. Hum. Brain Mapp. 2010, 31, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Luck, S. An Introduction to the Event-Related Potential Technique; The MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Wild-Wall, N.; Falkenstein, M.; Gajewski, P.D. Neural correlates of changes in a visual search task due to cognitive training in seniors. Neural Plast. 2012, 2012, 529057. [Google Scholar] [CrossRef] [PubMed]
- Tafuro, A.; Ambrosini, E.; Puccioni, O.; Vallesi, A. Brain oscillations in cognitive control: A cross-sectional study with a spatial stroop task. Neuropsychologia 2019, 133, 107190. [Google Scholar] [CrossRef]
- Liotti, M.; Woldorff, M.G.; Perez, R.; Mayberg, H.S. An ERP study of the temporal course of the Stroop color-word interference effect. Neuropsychologia 2000, 38, 701–711. [Google Scholar] [CrossRef]
- Markela-Lerenc, J.; Ille, N.; Kaiser, S.; Fiedler, P.; Mundt, C.; Weisbrod, M. Prefrontal-cingulate activation during executive control: Which comes first? Brain Res. Cogn. Brain Res. 2004, 18, 278–287. [Google Scholar] [CrossRef]
- Sahinoglu, B.; Dogan, G. Event-Related Potentials and the Stroop Effect. Eurasian J. Med. 2016, 48, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Spronk, M.; Jonkman, L.M. Electrophysiological evidence for different effects of working memory load on interference control in adolescents than adults. Int. J. Psychophysiol. 2012, 83, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Szucs, D.; Soltesz, F. Functional definition of the N450 event-related brain potential marker of conflict processing: A numerical Stroop study. BMC Neurosci. 2012, 13, 35. [Google Scholar] [CrossRef]
- Kessler, R.C.; Amminger, G.P.; Aguilar-Gaxiola, S.; Alonso, J.; Lee, S.; Ustün, T.B. Age of onset of mental disorders: A review of recent literature. Curr. Opin. Psychiatry 2007, 20, 359–364. [Google Scholar] [CrossRef]
- Cuijpers, P.; Smit, F.; Aalten, P.; Batelaan, N.; Klein, A.; Salemink, E.; Spinhoven, P.; Struijs, S.; Vonk, P.; Wiers, R.W.; et al. The Associations of Common Psychological Problems With Mental Disorders Among College Students. Front. Psychiatry. 2021, 12, 573637. [Google Scholar] [CrossRef] [PubMed]
- Gogtay, N.; Vyas, N.S.; Testa, R.; Wood, S.J.; Pantelis, C. Age of onset of schizophrenia: Perspectives from structural neuroimaging studies. Schizophr. Bull. 2011, 37, 504–513. [Google Scholar] [CrossRef]
- Lozano, V.; Soriano, M.F.; Aznarte, J.I.; Gómez-Ariza, C.J.; Bajo, M.T. Interference control commonalities in patients with schizophrenia, bipolar disorder, and borderline personality disorder. J. Clin. Exp. Neuropsychol. 2016, 38, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Spinella, M. Normative data and a short form of the Barratt Impulsiveness Scale. Neurosci. J. 2007, 117, 359–368. [Google Scholar] [CrossRef]
- Eysenck, H.J.; Eysenck, S.B.G. Eysenckov Upitnik Licnosti—EPQ; Naklada Slap: Jastrebarsko, Croatia, 1994. [Google Scholar]
- Sučević, D.; Momirović, A.; Fruk, G.; Auguštin, B. Kognitivni Neverbalni Test—KNT; Naklada Slap: Jastrebarsko, Croatia, 2004. [Google Scholar]
- Knežević, M.; Tarabić, B.N.; Tomac, P.; Vincek, A.; Ivanda, L. Role of age, gender and education in information processing speed. Psych. Top. 2015, 24, 173–185. Available online: https://hrcak.srce.hr/142127 (accessed on 4 April 2024).
- Marinkovic, K.; Rickenbacher, E.; Azma, S.; Artsy, E. Acute alcohol intoxication impairs top-down regulation of Stroop incongruity as revealed by blood oxygen level-dependent functional magnetic resonance imaging. Hum. Brain Mapp. 2012, 33, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Repovs, G. The mode of response and the Stroop effect: A reaction time analysis. Horiz. Psychol. 2004, 13, 105–114. Available online: http://psiholoska-obzorja.si/arhiv_clanki/2004_2/repovs2.pdf (accessed on 4 April 2024).
- Kappenman, E.S.; Gamble, M.L.; Luck, S. ERP Boot Camp: Data Analysis Tutorials; University of California: Davis, CA, USA, 2011. [Google Scholar]
- Zurron, M.; Pouso, M.; Lindín, M.; Galdo, S.; Díaz, F. Event-Related Potentials with the Stroop colour-word task: Timing of semantic conflict. Int. J. Psychophysiol. 2009, 72, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Jongen, E.M.M.; Jonkman, L.M. The developmental pattern of stimulus and response interference in a color-object Stroop task: An ERP study. BMC Neurosci. 2008, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.J.; Tavabi, K.; Caffarra, S.; Yeatman, J.D. Automaticity in the reading circuitry. Brain Lang. 2021, 214, 104906. [Google Scholar] [CrossRef] [PubMed]
- Wright, B.C. What Stroop tasks can tell us about selective attention from childhood to adulthood. Br. J. Psychol. 2017, 108, 583–607. [Google Scholar] [CrossRef] [PubMed]
- Macleod, C.M. Half a Century of Research on the Stroop Effect—An Integrative Review. Psychol. Bull. 1991, 109, 163–203. [Google Scholar] [CrossRef] [PubMed]
- Luna, B.; Padmanabhan, A.; O’Hearn, K. What has fMRI told us about the Development of Cognitive Control through Adolescence? Brain Cogn. 2010, 72, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.; Luna, B. Adolescence as a neurobiological critical period for the development of higher-order cognition. Neurosci. Biobehav. Rev. 2018, 94, 179–195. [Google Scholar] [CrossRef]
- Gajewski, P.D.; Falkenstein, M.; Thönes, S.; Wascher, E. Stroop task performance across the lifespan: High cognitive reserve in older age is associated with enhanced proactive and reactive interference control. Neuroimage 2020, 207, 116430. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Goicoa, M.; Galdo-Álvarez, S.; Díaz, F.; Zurrón, M. Effect of Normal Aging and of Mild Cognitive Impairment on Event-Related Potentials to a Stroop Color-Word Task. J. Alzheimer’s Dis. 2016, 52, 1487–1501. [Google Scholar] [CrossRef] [PubMed]
- West, R.; Langley, M.M.; Bailey, K. Signaling a switch: Neural correlates of task switching guided by task cues and transition cues. Psychophysiology 2011, 48, 612–623. [Google Scholar] [CrossRef]
- Zanto, T.P.; Rubens, M.T.; Thangavel, A.; Gazzaley, A. Causal role of the prefrontal cortex in top-down modulation of visual processing and working memory. Nat. Neurosci. 2011, 14, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Crowley, K.E.; Colrain, I.M. A review of the evidence for P2 being an independent component process: Age, sleep and modality. Clin. Neurophysiol. 2004, 115, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, Z.; Clark, K.; Merrikhi, Y.; Mueller, A.; Pettine, W.; Isabel Vanegas, M.; Moore, T.; Noudoost, B. Prefrontal Contributions to Attention and Working Memory. Curr. Top. Behav. Neurosci. 2019, 41, 129–153. [Google Scholar] [CrossRef] [PubMed]
- Iannaccone, R.; Hauser, T.U.; Staempfli, P.; Walitza, S.; Brandeis, D.; Brem, S. Conflict monitoring and error processing: New insights from simultaneous EEG-fMRI. Neuroimage 2015, 105, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Clawson, A.; Clayson, P.E.; Keith, C.M.; Catron, C.; Larson, M.J. Conflict and performance monitoring throughout the lifespan: An event-related potential (ERP) and temporospatial component analysis. Biol. Psychol. 2017, 124, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Larson, M.J.; Clayson, P.E. The relationship between cognitive performance and electrophysiological indices of performance monitoring. Cogn. Affect. Behav. Neurosci. 2011, 11, 159–171. [Google Scholar] [CrossRef]
- Gajewski, P.D.; Falkenstein, M. Diversity of the P3 in the task-switching paradigm. Brain Res. 2011, 1411, 87–97. [Google Scholar] [CrossRef]
- Ladouceur, C.D.; Dahl, R.E.; Carter, C.S. Development of action monitoring through adolescence into adulthood: ERP and source localization. Dev. Sci. 2007, 10, 874–891. [Google Scholar] [CrossRef] [PubMed]
- Vara, A.S.; Pang, E.W.; Vidal, J.; Anagnostou, E.; Taylor, M.J. Neural mechanisms of inhibitory control continue to mature in adolescence. Dev. Cogn. Neurosci. 2014, 10, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ojha, A.; Parr, A.C.; Foran, W.; Calabro, F.J.; Luna, B. Puberty contributes to adolescent development of fronto-striatal functional connectivity supporting inhibitory control. Dev. Cogn. Neurosci. 2022, 58, 101183. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Hernández, S.P.; Rahman, M.S.; Voigt, K.; Malvaso, A. Inhibitory Control Development: A Network Neuroscience Perspective. Front. Psychol. 2022, 13, 651547. [Google Scholar] [CrossRef] [PubMed]
- Petanjek, Z.; Judaš, M.; Šimic, G.; Rasin, M.R.; Uylings, H.B.; Rakic, P.; Kostovic, I. Extraordinary neoteny of synaptic spines in the human prefrontal cortex. Proc. Natl. Acad. Sci. USA 2011, 108, 13281–13286. [Google Scholar] [CrossRef] [PubMed]
- Huttenlocher, P.R.; Dabholkar, A.S. Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol. 1997, 20, 167–178. [Google Scholar] [CrossRef]
- Logue, S.F.; Gould, T.J. The neural and genetic basis of executive function: Attention, cognitive flexibility, and response inhibition. Pharmacol. Biochem. Behav. 2014, 123, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Markon, K.E.; Clark, L. Toward a theory of distinct types of “impulsive” behaviors: A meta-analysis of self-report and behavioral measures. Psychol. Bull. 2014, 140, 374–408. [Google Scholar] [CrossRef]
- Fishman, I.; Ng, R. Error-related brain activity in extraverts: Evidence for altered response monitoring in social context. Biol. Psychol. 2013, 3, 225–230. [Google Scholar] [CrossRef]
- Arnett, J.J. Emerging Adulthood: The Winding Road from Late Teens through the Twenties; Oxford University Press: Oxford, UK, 2015. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Early 20s M (±SD) | Mid 20s M (±SD) | Early 30s M (±SD) | F(2,128) | p | ŋp2 |
|---|---|---|---|---|---|---|
| Non-verbal IQ | 33.3 (±4.45) | 32.1 (±5.24) | 31.6 (±6.00) | 1.2 | 0.30 | 0.02 |
| Info. processing | 45.0 (±5.24) | 45.4 (±5.33) | 43.8 (±4.99) | 1.1 | 0.35 | 0.02 |
| Impulsivity | 29.6 (±4.52) | 29.9 (±5.32) | 30.1 (±4.88) | 0.1 | 0.87 | 0.00 |
| Psychoticism | 3.8 (±2.34) | 4.0 (±2.10) | 4.3 (±2.09) | 0.7 | 0.52 | 0.01 |
| Extraversion | 14.5 (±4.54) | 15.0 (±4.43) | 14.4 (±4.07) | 0.2 | 0.83 | 0.00 |
| Neuroticism | 9.1 (±4.68) | 8.2 (±5.53) | 6.9 (±4.55) | 2.1 | 0.12 | 0.03 |
| Variable | F(2,128) | p | ŋp2 |
|---|---|---|---|
| Accuracy | |||
| Incongruent | 4.1 | 0.02 * | 0.06 |
| Congruent | 1.8 | 0.17 | 0.03 |
| Reaction Time | |||
| Incongruent | 0.9 | 0.42 | 0.01 |
| Congruent | 1.2 | 0.30 | 0.02 |
| Component | Condition | F(2,128) | p | ŋp2 |
|---|---|---|---|---|
| Fz electrode | ||||
| P2 | Incongruent | 4.1 | 0.02 * | 0.06 |
| Congruent | 2.9 | 0.06 | 0.04 | |
| N2 | Incongruent | 4.9 | 0.01 * | 0.07 |
| Congruent | 3.8 | 0.02 * | 0.06 | |
| P3 | Incongruent | 2.8 | 0.06 | 0.04 |
| Congruent | 4.9 | 0.01 * | 0.07 | |
| N450 | Incongruent | 6.1 | <0.01 ** | 0.09 |
| Congruent | 2.4 | 0.10 | 0.04 | |
| Cz electrode | ||||
| P2 | Incongruent | 5.4 | <0.01 ** | 0.08 |
| Congruent | 5.9 | <0.01 ** | 0.08 | |
| N2 | Incongruent | 2.7 | 0.07 | 0.04 |
| Congruent | 2.6 | 0.07 | 0.04 | |
| P3 | Incongruent | 1.1 | 0.33 | 0.02 |
| Congruent | 0.7 | 0.50 | 0.01 | |
| N450 | Incongruent | 2.9 | 0.06 | 0.04 |
| Congruent | 2.0 | 0.14 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knežević, M. Temporal Course of Interference Control from Early to Late Young Adulthood: An ERP Study. Brain Sci. 2024, 14, 536. https://doi.org/10.3390/brainsci14060536
Knežević M. Temporal Course of Interference Control from Early to Late Young Adulthood: An ERP Study. Brain Sciences. 2024; 14(6):536. https://doi.org/10.3390/brainsci14060536
Chicago/Turabian StyleKnežević, Martina. 2024. "Temporal Course of Interference Control from Early to Late Young Adulthood: An ERP Study" Brain Sciences 14, no. 6: 536. https://doi.org/10.3390/brainsci14060536
APA StyleKnežević, M. (2024). Temporal Course of Interference Control from Early to Late Young Adulthood: An ERP Study. Brain Sciences, 14(6), 536. https://doi.org/10.3390/brainsci14060536