Overt and Covert Effects of Mental Fatigue on Attention Networks: Evidence from Event-Related Potentials during the Attention Network Test


Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Methods
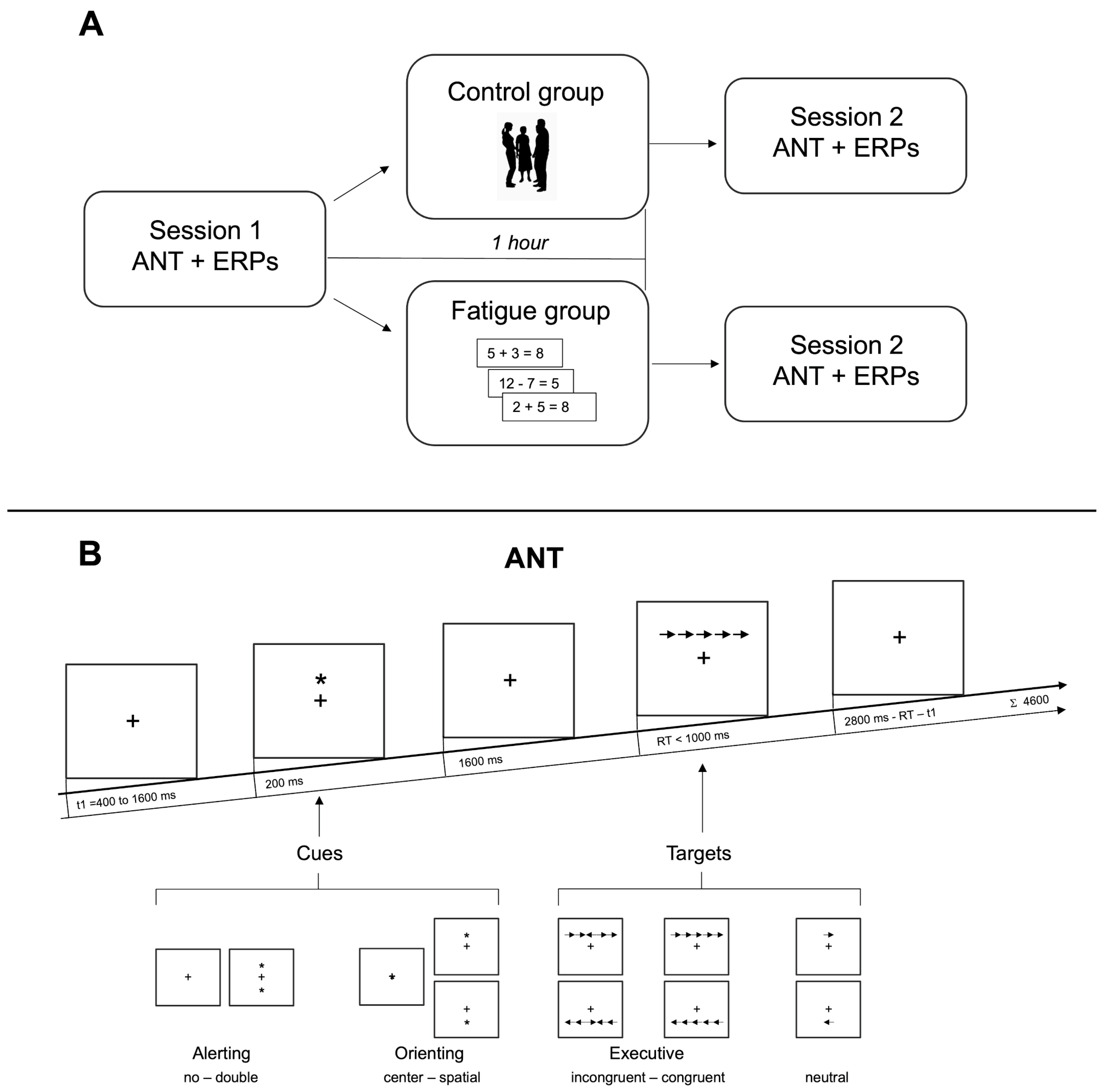
2.1.1. Subjects
2.1.2. Procedure
2.1.3. EEG Recording
2.1.4. Paradigm
2.1.5. ERP Analysis
- -
- Alerting: target N1nocue vs. target N1doublecue;
- -
- Orienting: target N1centralcue vs. target N1spatialcue;
- -
- Conflict: target N2incongruent vs. target N2congruent;
- -
- Alerting: target P3nocue vs. target P3doublecue;
- -
- Orienting: target P3centralcue vs. target P3spatialcue;
- -
- Conflict: target P3incongruent vs. target P3congruent;
2.2. Statistical Analyses
2.2.1. Overt Responses: ANT Analysis
2.2.2. Covert Responses: ERP Analyses
3. Results
3.1. Overt Responses
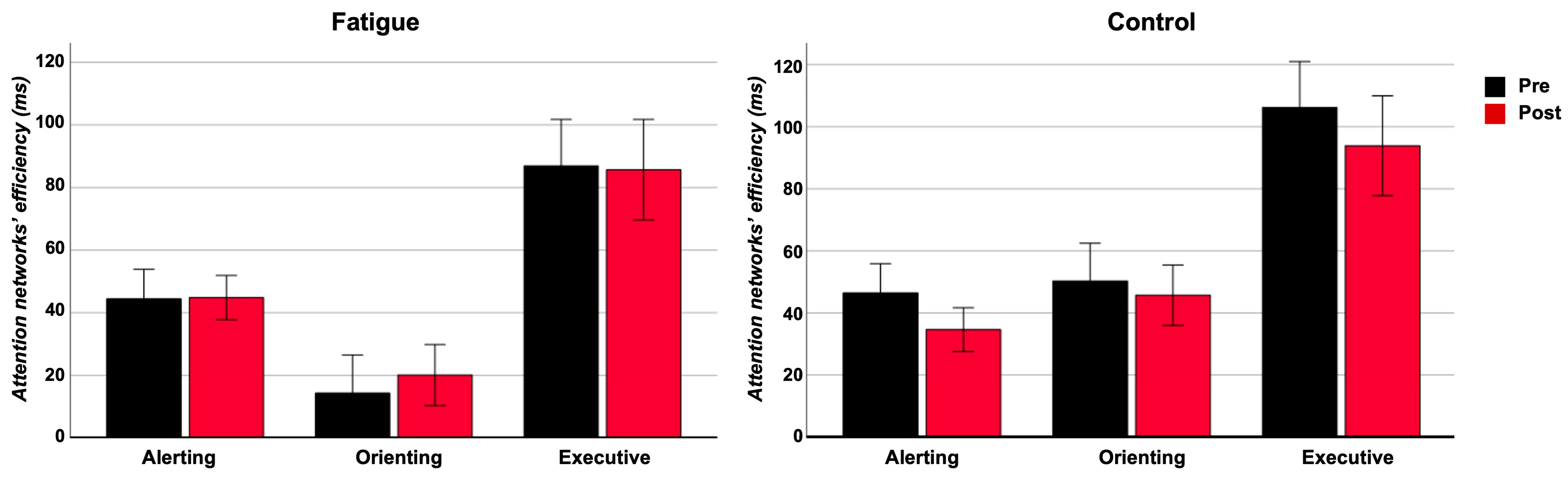
3.1.1. Network Efficiency
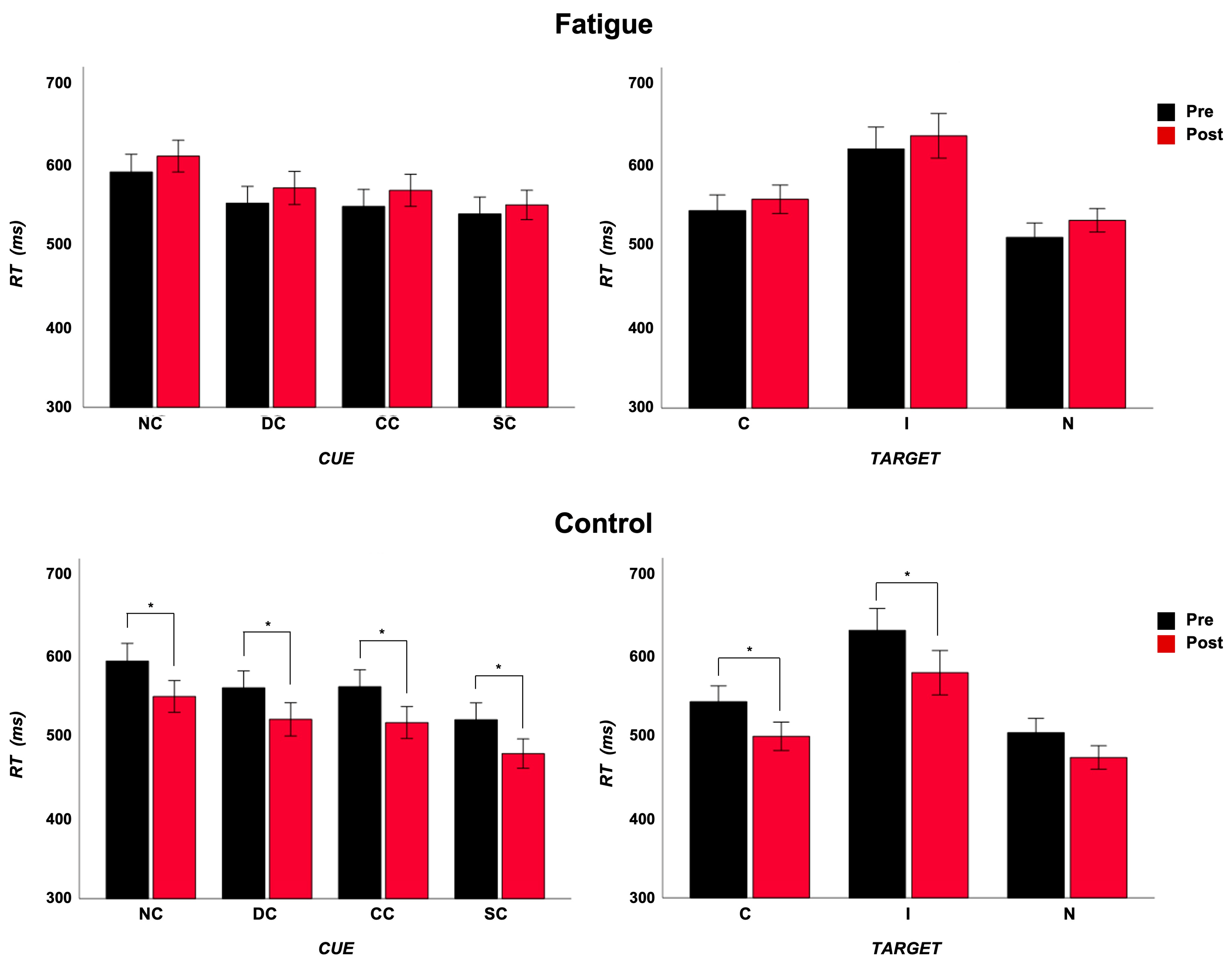
3.1.2. RT
3.1.3. Accuracy
3.2. Covert Responses: ERP Results
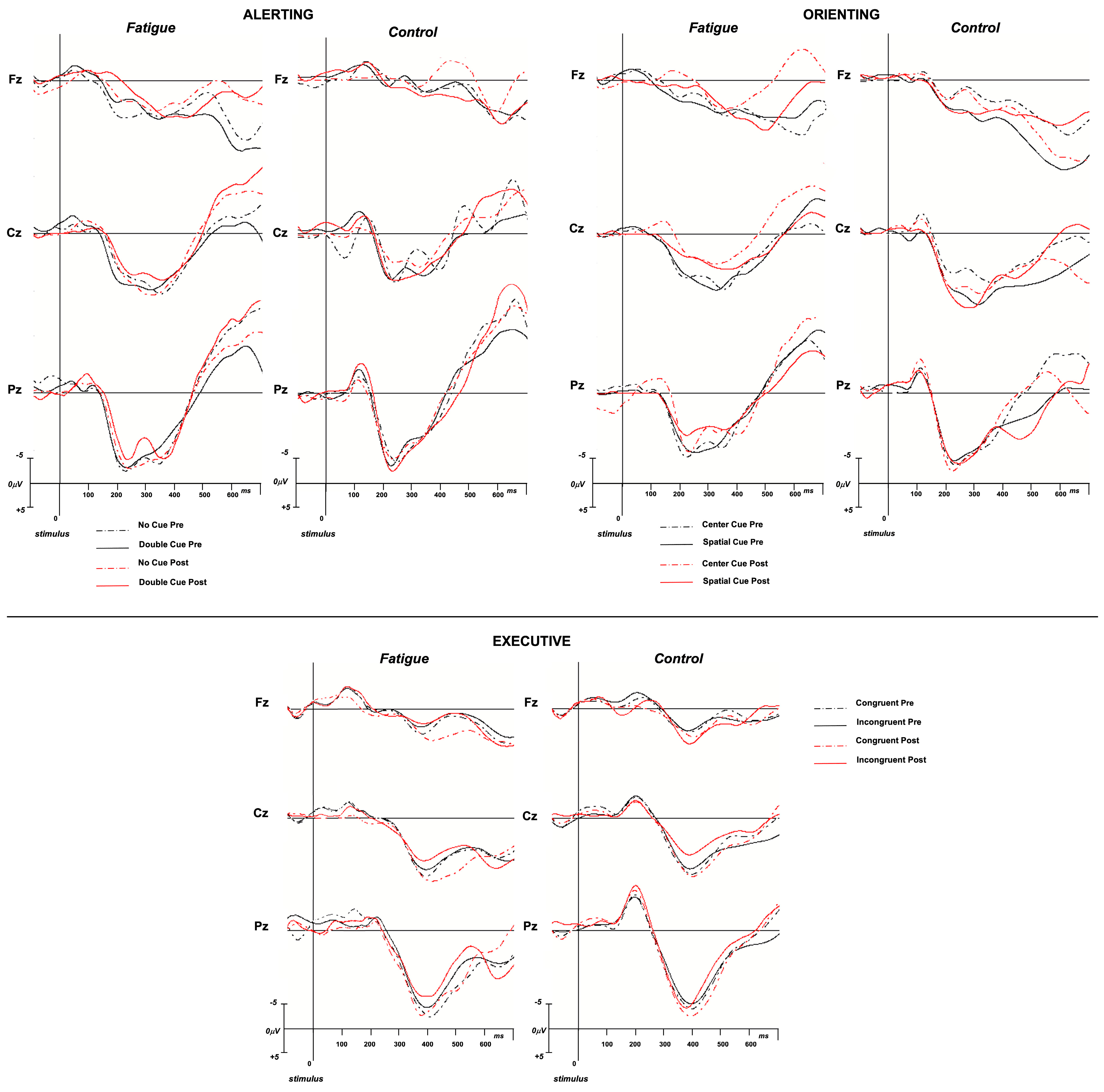
3.2.1. Alerting—N1
3.2.2. Alerting—P3
3.2.3. Orienting—N1
3.2.4. Orienting—P3
3.2.5. Conflict—N2
3.2.6. Conflict—P3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coull, J. Neural correlates of attention and arousal: Insights from electrophysiology, functional neuroimaging and psychopharmacology. Prog. Neurobiol. 1998, 55, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Block, S.R.; King, A.P.; Sripada, R.K.; Weissman, D.H.; Welsh, R.; Liberzon, I. Behavioral and neural correlates of disrupted orienting attention in posttraumatic stress disorder. Cogn. Affect. Behav. Neurosci. 2017, 17, 422–436. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.E.; Posner, M.I. The Attention System of the Human Brain: 20 Years After. Annu. Rev. Neurosci. 2012, 35, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Raz, A.; Buhle, J. Typologies of attentional networks. Nat. Rev. Neurosci. 2006, 7, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; McCandliss, B.D.; Fossella, J.; Flombaum, J.I.; Posner, M.I. The activation of attentional networks. NeuroImage 2005, 26, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; McCandliss, B.D.; Sommer, T.; Raz, A.; Posner, M.I. Testing the Efficiency and Independence of Attentional Networks. J. Cogn. Neurosci. 2002, 14, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Raz, A. Anatomy of attentional networks. Anat. Rec. B New Anat. 2004, 281, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.C.; Barry, R.J.; Connolly, J.F.; Fischer, C.; Michie, P.T.; Näätänen, R.; Polich, J.; Reinvang, I.; Van Petten, C. Event-related potentials in clinical research: Guidelines for eliciting, recording, and quantifying mismatch negativity, P300, and N400. Clin. Neurophysiol. 2009, 120, 1883–1908. [Google Scholar] [CrossRef]
- Neuhaus, A.H.; Urbanek, C.; Opgen-Rhein, C.; Hahn, E.; Ta, T.M.T.; Koehler, S.; Gross, M.; Dettling, M. Event-related potentials associated with Attention Network Test. Int. J. Psychophysiol. 2010, 76, 72–79. [Google Scholar] [CrossRef]
- Luck, S.J. An Introduction to the Event-Related Potential Technique, 2nd ed.; MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Verleger, R. On the utility of P3 latency as an index of mental chronometry. Psychophysiology 1997, 34, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Folstein, J.R.; Van Petten, C. Influence of cognitive control and mismatch on the N2 component of the ERP: A review. Psychophysiology 2008, 45, 152–170. [Google Scholar] [CrossRef] [PubMed]
- Boksem, M.A.; Tops, M. Mental fatigue: Costs and benefits. Brain Res. Rev. 2008, 59, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Lorist, M.M. Impact of top-down control during mental fatigue. Brain Res. 2008, 1232, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Massar, S.A.; Wester, A.E.; Volkerts, E.R.; Kenemans, J.L. Manipulation specific effects of mental fatigue: Evidence from novelty processing and simulated driving. Psychophysiology 2010, 47, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Boksem, M.A.; Meijman, T.F.; Lorist, M.M. Effects of mental fatigue on attention: An ERP study. Cogn. Brain Res. 2005, 25, 107–116. [Google Scholar] [CrossRef] [PubMed]
- van der Linden, D.; Frese, M.; Meijman, T.F. Mental fatigue and the control of cognitive processes: Effects on perseveration and planning. Acta Psychol. 2003, 113, 45–65. [Google Scholar] [CrossRef]
- Boksem, M.A.; Meijman, T.F.; Lorist, M.M. Mental fatigue, motivation and action monitoring. Biol. Psychol. 2005, 72, 123–132. [Google Scholar] [CrossRef]
- Kato, Y.; Endo, H.; Kizuka, T. Mental fatigue and impaired response processes: Event-related brain potentials in a Go/NoGo task. Int. J. Psychophysiol. 2009, 72, 204–211. [Google Scholar] [CrossRef]
- Yang, B.; Xiao, W.; Liu, X.; Wu, S.; Miao, D. Mental fatigue impairs pre-attentive processing: A MMN study. Neurosci. Lett. 2013, 532, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.; Behan, P.O. Fatigue in neurological disorders. Lancet 2004, 363, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Togo, F.; Lange, G.; Natelson, B.H.; Quigley, K.S. Attention network test: Assessment of cognitive function in chronic fatigue syndrome. J. Neuropsychol. 2015, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Moss-Morris, R.; Risdale, A.; Lynch, J.; Jeevaratnam, P.; Bradley, B.P.; Mogg, K. Attention processes in chronic fatigue syndrome: Attentional bias for health-related threat and the role of attentional control. Behav. Res. Ther. 2014, 52, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Keynejad, R.C.; Keynejad, R.C.; Fenby, E.; Fenby, E.; Pick, S.; Pick, S.; Moss-Morris, R.; Moss-Morris, R.; Hirsch, C.; Hirsch, C.; et al. Attentional Processing and Interpretative Bias in Functional Neurological Disorder. Psychosom. Med. 2020, 82, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Monsell, S. Task switching. Trends Cogn. Sci. 2003, 7, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.; Behan, P.O. Fatigue and basal ganglia. J. Neurol. Sci. 2000, 179, 34–42. [Google Scholar] [CrossRef]
- Pauletti, C.; Mannarelli, D.; Locuratolo, N.; Pollini, L.; Currà, A.; Marinelli, L.; Rinalduzzi, S.; Fattapposta, F. Attention in Parkinson’s disease with fatigue: Evidence from the attention network test. J. Neural Transm. 2017, 124, 335–345. [Google Scholar] [CrossRef]
- Möckel, T.; Beste, C.; Wascher, E. The Effects of Time on Task in Response Selection—An ERP Study of Mental Fatigue. Sci. Rep. 2015, 5, 10113. [Google Scholar] [CrossRef]
- McNair, D.; Lorr, M.; Droppleman, L. Manual for the Profile of Mood States; Educational and Industrial Testing Service: San Diego, CA, USA, 1971. [Google Scholar]
- Picton, T.; Bentin, S.; Berg, P.; Donchin, E.; Hillyard, S.; Johnson, R.; Miller, G.; Ritter, W.; Ruchkin, D.; Rugg, M.D.; et al. Guidelines for using human event-related potentials to study cognition: Recording standards and publication criteria. Psychophysiology 2000, 37, 127–152. [Google Scholar] [CrossRef]
- Kratz, O.; Studer, P.; Malcherek, S.; Erbe, K.; Moll, G.H.; Heinrich, H. Attentional processes in children with ADHD: An event-related potential study using the attention network test. Int. J. Psychophysiol. 2011, 81, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Zholdassova, M.; Kustubayeva, A.; Matthews, G. The ANT Executive Control Index: No Evidence for Temporal Decrement. Hum. Factors J. Hum. Factors Ergon. Soc. 2021, 63, 254–273. [Google Scholar] [CrossRef] [PubMed]
- Moghani, M.K.; Zeidabadi, R.; Kaseb, M.R.S.; Borujeni, I.B. Mental Fatigue Reduces the Benefits of Self-Controlled Feedback on Learning a Force Production Task. Percept. Mot. Ski. 2021, 128, 2398–2414. [Google Scholar] [CrossRef] [PubMed]
- Vogel, E.K.; Luck, S.J. The Visual N1 Component as an Index of a Discrimination Process. Psychophysiology 2000, 37, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Groom, M.J.; Cragg, L. Differential modulation of the N2 and P3 event-related potentials by response conflict and inhibition. Brain Cogn. 2015, 97, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hinss, M.F.; Brock, A.M.; Roy, R.N. The double task-switching protocol: An investigation into the effects of similarity and conflict on cognitive flexibility in the context of mental fatigue. PLoS ONE 2023, 18, e0279021. [Google Scholar] [CrossRef]
- Yu, S.; Mückschel, M.; Beste, C. Event-related synchronization/desynchronization and functional neuroanatomical regions associated with fatigue effects on cognitive flexibility. J. Neurophysiol. 2021, 126, 383–397. [Google Scholar] [CrossRef]
- Jennings, J.M.; Dagenbach, D.; Engle, C.M.; Funke, L.J. Age-Related Changes and the Attention Network Task: An Examination of Alerting, Orienting, and Executive Function. Aging, Neuropsychol. Cogn. 2007, 14, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.S.; Biel, A.L.; Wegier, P.; Lapp, L.K.; Dyson, B.J.; Spaniol, J. Age differences in the Attention Network Test: Evidence from behavior and event-related potentials. Brain Cogn. 2016, 102, 65–79. [Google Scholar] [CrossRef]
- Dash, T.; Berroir, P.; Joanette, Y.; Ansaldo, A.I. Alerting, Orienting, and Executive Control: The Effect of Bilingualism and Age on the Subcomponents of Attention. Front. Neurol. 2019, 10, 1122. [Google Scholar] [CrossRef]
- Lorist, M.M.; Boksem, M.A.; Ridderinkhof, K.R. Impaired cognitive control and reduced cingulate activity during mental fatigue. Cogn. Brain Res. 2005, 24, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Faber, L.G.; Maurits, N.M.; Lorist, M.M. Mental Fatigue Affects Visual Selective Attention. PLoS ONE 2012, 7, e48073. [Google Scholar] [CrossRef] [PubMed]
- Lorist, M.M.; Klein, M.; Nieuwenhuis, S.; De Jong, R.; Mulder, G.; Meijman, T.F. Mental Fatigue and Task Control: Planning and Preparation. Psychophysiology 2000, 37, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Ren, J.; Wang, B.; Zhu, Q. Effects of Relaxing Music on Mental Fatigue Induced by a Continuous Performance Task: Behavioral and ERPs Evidence. PLoS ONE 2015, 10, e0136446. [Google Scholar] [CrossRef] [PubMed]
- Dotare, M.; Bader, M.; Mesrobian, S.K.; Asai, Y.; Villa, A.E.P.; Lintas, A. Attention Networks in ADHD Adults after Working Memory Training with a Dual n-Back Task. Brain Sci. 2020, 10, 715. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aversion Scale | POMS—Fatigue | |||||
|---|---|---|---|---|---|---|
| Pre | Post | p * | Pre | Post | p * | |
| NF | 0.8 ± 2.3 | 0.6 ± 1.7 | 0.64 | 6.5 ± 3.5 | 7.1 ± 3.5 | 0.48 |
| F | 0.4 ± 0.7 | 5.4 ± 2.2 | <0.001 | 9.0 ± 6.3 | 10.9 ± 6.7 | 0.02 |
| FLANKER | CUE | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| NC | DC | CC | SC | ||||||
| Mean RT ms (SD) | PRE | POST | PRE | POST | PRE | POST | PRE | POST | |
| NF | Congruent | 570 (63) | 516 (51) | 532 (49) | 492 (59) | 532 (57) | 489 (58) | 493 (68) | 444 (65) |
| Incongruent | 653 (92) | 602 (92) | 640 (110) | 578 (83) | 634 (94) | 565 (70) | 574 (100) | 515 (92) | |
| Neutral | 531 (38) | 493 (61) | 492 (51) | 456 (47) | 485 (43) | 455 (48) | 466 (57) | 430 (53) | |
| F | Congruent | 563 (94) | 572 (75) | 526 (93) | 546 (85) | 523 (98) | 546 (88) | 518 (98) | 513 (58) |
| Incongruent | 655 (138) | 662 (124) | 609 (120) | 636 (129) | 592 (123) | 625 (133) | 593 (118) | 588 (104) | |
| Neutral | 516 (106) | 560 (78) | 487 (92) | 500 (56) | 488 (79) | 506 (61) | 500 (84) | 504 (60) | |
| % Accuracy (SD) | |||||||||
| NF | Congruent | 1.00 (0) | 1.00 (0) | 1.00 (0) | 1.00 (0) | 1.00 (0) | 1.00 (0) | 1.00 (0.02) | 1.00 (0) |
| Incongruent | 0.98 (0.06) | 0.98 (0.04) | 0.98 (0.03) | 0.99 (0.05) | 0.99 (0.03) | 0.99 (0.03) | 1.00 (0.02) | 1.00 (0.02) | |
| Neutral | 0.99 (0.02) | 1.00 (0) | 1.00 (0) | 1.00 (0) | 0.99 (0.04) | 1.00 (0.02) | 0.99 (0.02) | 1.00 (0) | |
| F | Congruent | 1.00 (0.02) | 0.99 (0.02) | 1.00 (0) | 1.00 (0) | 1.00 (0) | 1.00 (0) | 1.00 (0.02) | 0.93 (0.26) |
| Incongruent | 0.99 (0.03) | 0.96 (0.06) | 0.97 (0.04) | 0.97 (0.05) | 0.99 (0.03) | 0.98 (0.05) | 0.99 (0.03) | 0.97 (0.04) | |
| Neutral | 1.00 (0) | 0.99 (0.02) | 1.00 (0) | 1.00 (0.02) | 1.00 (0.02) | 1.00 (0) | 1.00 (0.02) | 0.99 (0.03) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pauletti, C.; Mannarelli, D.; Fattapposta, F. Overt and Covert Effects of Mental Fatigue on Attention Networks: Evidence from Event-Related Potentials during the Attention Network Test. Brain Sci. 2024, 14, 803. https://doi.org/10.3390/brainsci14080803
Pauletti C, Mannarelli D, Fattapposta F. Overt and Covert Effects of Mental Fatigue on Attention Networks: Evidence from Event-Related Potentials during the Attention Network Test. Brain Sciences. 2024; 14(8):803. https://doi.org/10.3390/brainsci14080803
Chicago/Turabian StylePauletti, Caterina, Daniela Mannarelli, and Francesco Fattapposta. 2024. "Overt and Covert Effects of Mental Fatigue on Attention Networks: Evidence from Event-Related Potentials during the Attention Network Test" Brain Sciences 14, no. 8: 803. https://doi.org/10.3390/brainsci14080803
APA StylePauletti, C., Mannarelli, D., & Fattapposta, F. (2024). Overt and Covert Effects of Mental Fatigue on Attention Networks: Evidence from Event-Related Potentials during the Attention Network Test. Brain Sciences, 14(8), 803. https://doi.org/10.3390/brainsci14080803