
Neuroimmunology of the Interleukins 13 and 4



Abstract
:
{kind=link}
{kind=link}
1. Introduction
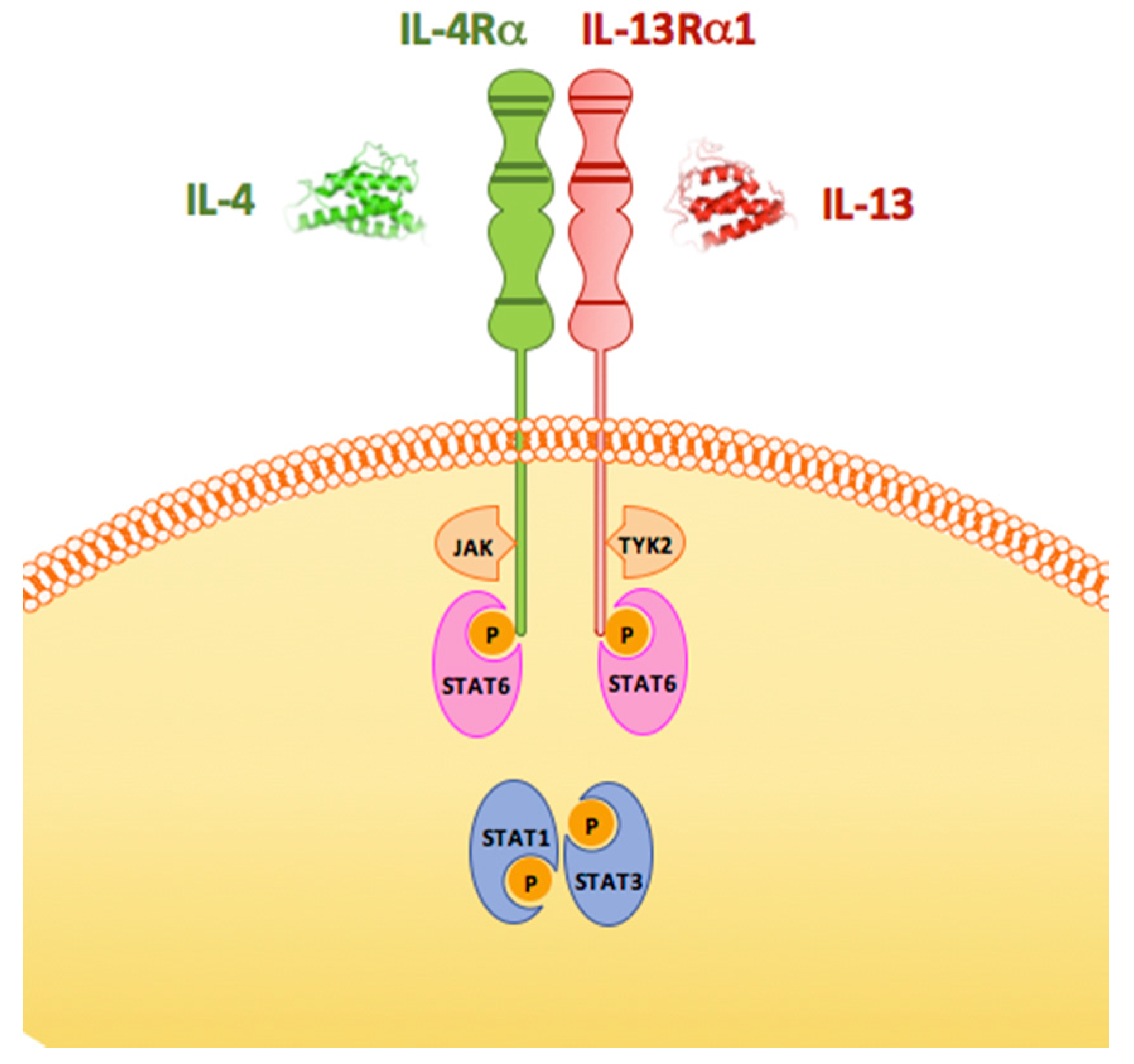
2. What We Know about IL-13 Comes from Studies of Its Biology in the Immune System
3. Expression of IL-13 and IL-4 in the CNS
4. What Is the Role of IL-13 and IL-4 in the CNS?
5. IL-13 and IL-4 in Multiple Sclerosis
6. Parkinson’s Disease
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morrison, B.E.; Marcondes, M.C.; Nomura, D.K.; Sanchez-Alavez, M.; Sanchez-Gonzalez, A.; Saar, I.; Kim, K.S.; Bartfai, T.; Maher, P.; Sugama, S.; et al. Cutting edge: IL-13Rα1 expression in dopaminergic neurons contributes to their oxidative stress-mediated loss following chronic peripheral treatment with lipopolysaccharide. J. Immunol. 2012, 189, 5498–5502. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, B.F.; Haas, H.; Falcone, F.H.; Albrecht, C.; Vollrath, I.B.; Noll, T.; Wolff, H.H.; Amon, U. Purified human peripheral blood basophils release interleukin-13 and preformed interleukin-4 following immunological activation. Eur. J. Immunol. 1996, 26, 2493–2498. [Google Scholar] [CrossRef] [PubMed]
- Haas, H.; Falcone, F.H.; Holland, M.J.; Schramm, G.; Haisch, K.; Gibbs, B.F.; Bufe, A.; Schlaak, M. Early interleukin-4: Its role in the switch towards a Th2 response and IgE-mediated allergy. Int. Arch. Allergy Immunol. 1999, 119, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Howard, M.; Paul, W.E. Interleukins for B lymphocytes. Lymphokine Res. 1982, 1, 1–4. [Google Scholar] [PubMed]
- Jaffe, J.S.; Raible, D.G.; Post, T.J.; Wang, Y.; Glaum, M.C.; Butterfield, J.H.; Schulman, E.S. Human lung mast cell activation leads to IL-13 mRNA expression and protein release. Am. J. Respir. Cell Mol. Biol. 1996, 15, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Battaile, J.T.; Patel, A.C.; You, Y.; Agapov, E.; Grayson, M.H.; Benoit, L.A.; Byers, D.E.; Alevy, Y.; Tucker, J.; et al. Persistent activation of an innate immune response translates respiratory viral infection into chronic lung disease. Nat. Med. 2008, 14, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Lee, K.H.; Lee, H.B.; Rhee, Y.K. Serum levels of interleukins (IL)-4, IL-5, IL-13, and interferon-gamma in acute asthma. J. Asthma 2001, 38, 665–671. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.N.; Culpepper, J.A.; de Waal Malefyt, R.; Briere, F.; Punnonen, J.; Aversa, G.; Sato, A.; Dang, W.; Cocks, G.B.; Menon, S.; et al. Interleukin 13, a T-cell-derived cytokine that regulates human monocyte and B-cell function. Proc. Natl. Acad. Sci. USA 1993, 90, 3735–3739. [Google Scholar] [CrossRef] [PubMed]
- Minty, A.; Chalon, P.; Derocq, J.M.; Dumont, X.; Guillemot, J.C.; Kaghad, M.; Labit, C.; Leplatois, P.; Liauzun, P.; Miloux, B.; et al. Interleukin-13 is a new human lymphokine regulating inflammatory and immune responses. Nature 1993, 362, 248–250. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.G.; Dolganov, G.M.; Robbins, S.E.; Hinton, L.M.; Lovett, M. The selective isolation of novel cDNAs encoded by the regions surrounding the human interleukin 4 and 5 genes. Nucleic Acids Res. 1992, 20, 5173–5179. [Google Scholar] [CrossRef] [PubMed]
- Ochensberger, B.; Daepp, G.C.; Rihs, S.; Dahinden, C.A. Human blood basophils produce interleukin-13 in response to IgE-receptor-dependent and -independent activation. Blood 1996, 88, 3028–3037. [Google Scholar] [PubMed]
- Reglier, H.; Arce-Vicioso, M.; Fay, M.; Gougerot-Pocidalo, M.A.; Chollet-Martin, S. Lack of IL-10 and IL-13 production by human polymorphonuclear neutrophils. Cytokine 1998, 10, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Grendelmeier, P.; Altznauer, F.; Fischer, B.; Bizer, C.; Straumann, A.; Menz, G.; Blaser, K.; Wuthrich, B.; Simon, H.U. Eosinophils express functional IL-13 in eosinophilic inflammatory diseases. J. Immunol. 2002, 169, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, M.; Bontempelli, M.; Giacomelli, M.; Malerba, M.; Rossi, G.A.; Di Stefano, A.; Rossi, A.; Ricciardolo, F.L. High serum levels of tumour necrosis factor-α and interleukin-8 in severe asthma: Markers of systemic inflammation? Clin. Exp. Allergy 2006, 36, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Wills-Karp, M.; Luyimbazi, J.; Xu, X.; Schofield, B.; Neben, T.Y.; Karp, C.L.; Donaldson, D.D. Interleukin-13: Central mediator of allergic asthma. Science 1998, 282, 2258–2261. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Ho, C.Y.; Ko, F.W.; Chan, C.H.; Ho, A.S.; Hui, D.S.; Lam, C.W. Proinflammatory cytokines (IL-17, IL-6, IL-18 and IL-12) and Th cytokines (IFN-gamma, IL-4, IL-10 and IL-13) in patients with allergic asthma. Clin. Exp. Immunol. 2001, 125, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. IL-13 effector functions. Annu. Rev. Immunol. 2003, 21, 425–456. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.D.; Zurawski, S.M.; Mosmann, T.R.; Zurawski, G. A family of small inducible proteins secreted by leukocytes are members of a new superfamily that includes leukocyte and fibroblast-derived inflammatory agents, growth factors, and indicators of various activation processes. J. Immunol. 1989, 142, 679–687. [Google Scholar] [PubMed]
- Kuperman, D.A.; Schleimer, R.P. Interleukin-4, interleukin-13, signal transducer and activator of transcription factor 6, and allergic asthma. Curr. Mol. Med. 2008, 8, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Wills-Karp, M. Interleukin-13 in asthma pathogenesis. Immunol. Rev. 2004, 202, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Chiba, N.; Masuda, A.; Yoshikai, Y.; Matsuguchi, T. Ceramide inhibits LPS-induced production of IL-5, IL-10, and IL-13 from mast cells. J. Cell. Physiol. 2007, 213, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Gordon, J.R.; Wershil, B.K. Cytokine production by mast cells and basophils. Curr. Opin. Immunol. 1991, 3, 865–872. [Google Scholar] [CrossRef]
- Masuda, A.; Yoshikai, Y.; Aiba, K.; Matsuguchi, T. Th2 cytokine production from mast cells is directly induced by lipopolysaccharide and distinctly regulated by c-Jun N-terminal kinase and p38 pathways. J. Immunol. 2002, 169, 3801–3810. [Google Scholar] [CrossRef] [PubMed]
- Supajatura, V.; Ushio, H.; Nakao, A.; Okumura, K.; Ra, C.; Ogawa, H. Protective roles of mast cells against enterobacterial infection are mediated by Toll-like receptor 4. J. Immunol. 2001, 167, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Aman, M.J.; Tayebi, N.; Obiri, N.I.; Puri, R.K.; Modi, W.S.; Leonard, W.J. cDNA cloning and characterization of the human interleukin 13 receptor α chain. J. Biol. Chem. 1996, 271, 29265–29270. [Google Scholar] [PubMed]
- Callard, R.E.; Matthews, D.J.; Hibbert, L. IL-4 and IL-13 receptors: Are they one and the same? Immunol. Today 1996, 17, 108–110. [Google Scholar] [CrossRef]
- Hilton, D.J.; Zhang, J.G.; Metcalf, D.; Alexander, W.S.; Nicola, N.A.; Willson, T.A. Cloning and characterization of a binding subunit of the interleukin 13 receptor that is also a component of the interleukin 4 receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Harris, M.B.; Rothman, P. IL-4/IL-13 signaling beyond JAK/STAT. J. Allergy Clin. Immunol. 2000, 105 Pt 1, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Leonard, W.J.; O’Shea, J.J. Jaks and STATs: Biological implications. Annu. Rev. Immunol. 1998, 16, 293–322. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.X.; Migone, T.S.; Tsang, M.; Friedmann, M.; Weatherbee, J.A.; Zhou, L.; Yamauchi, A.; Bloom, E.T.; Mietz, J.; John, S.; et al. The role of shared receptor motifs and common Stat proteins in the generation of cytokine pleiotropy and redundancy by IL-2, IL-4, IL-7, IL-13, and IL-15. Immunity 1995, 2, 331–339. [Google Scholar] [CrossRef]
- Nelms, K.; Keegan, A.D.; Zamorano, J.; Ryan, J.J.; Paul, W.E. The IL-4 receptor: Signaling mechanisms and biologic functions. Annu. Rev. Immunol. 1999, 17, 701–738. [Google Scholar] [CrossRef] [PubMed]
- Orchansky, P.L.; Ayres, S.D.; Hilton, D.J.; Schrader, J.W. An interleukin (IL)-13 receptor lacking the cytoplasmic domain fails to transduce IL-13-induced signals and inhibits responses to IL-4. J. Biol. Chem. 1997, 272, 22940–22947. [Google Scholar] [CrossRef] [PubMed]
- Caput, D.; Laurent, P.; Kaghad, M.; Lelias, J.M.; Lefort, S.; Vita, N.; Ferrara, P. Cloning and characterization of a specific interleukin (IL)-13 binding protein structurally related to the IL-5 receptor α chain. J. Biol. Chem. 1996, 271, 16921–16926. [Google Scholar] [PubMed]
- Donaldson, D.D.; Whitters, M.J.; Fitz, L.J.; Neben, T.Y.; Finnerty, H.; Henderson, S.L.; O’Hara, R.M., Jr.; Beier, D.R.; Turner, K.J.; Wood, C.R.; et al. The murine IL-13 receptor α 2: Molecular cloning, characterization, and comparison with murine IL-13 receptor α 1. J. Immunol. 1998, 161, 2317–2324. [Google Scholar] [PubMed]
- Feng, N.; Lugli, S.M.; Schnyder, B.; Gauchat, J.F.; Graber, P.; Schlagenhauf, E.; Schnarr, B.; Wiederkehr-Adam, M.; Duschl, A.; Heim, M.H.; et al. The interleukin-4/interleukin-13 receptor of human synovial fibroblasts: Overexpression of the nonsignaling interleukin-13 receptor α2. Lab. Investig. 1998, 78, 591–602. [Google Scholar] [PubMed]
- Liu, H.; Jacobs, B.S.; Liu, J.; Prayson, R.A.; Estes, M.L.; Barnett, G.H.; Barna, B.P. Interleukin-13 sensitivity and receptor phenotypes of human glial cell lines: Non-neoplastic glia and low-grade astrocytoma differ from malignant glioma. Cancer Immunol. Immunother. 2000, 49, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Debinski, W.; Gibo, D.M.; Hulet, S.W.; Connor, J.R.; Gillespie, G.Y. Receptor for interleukin 13 is a marker and therapeutic target for human high-grade gliomas. Clin. Cancer Res. 1999, 5, 985–990. [Google Scholar] [PubMed]
- Sengupta, S.; Thaci, B.; Crawford, A.C.; Sampath, P. Interleukin-13 receptor α 2-targeted glioblastoma immunotherapy. Biomed. Res. Int. 2014, 2014, 952128. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.J.; Wu, Q.; Kammer, W.; Friedrich, K.; Smith, J.M.; Kerr, I.M.; Stark, G.R.; Williams, B.R. Receptor-associated constitutive protein tyrosine phosphatase activity controls the kinase function of JAK1. Proc. Natl. Acad. Sci. USA 1997, 94, 8563–8568. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.N.; Noble, M.E.; Owen, D.J. Active and inactive protein kinases: Structural basis for regulation. Cell 1996, 85, 149–158. [Google Scholar] [CrossRef]
- Hou, J.; Schindler, U.; Henzel, W.J.; Ho, T.C.; Brasseur, M.; McKnight, S.L. An interleukin-4-induced transcription factor: IL-4 Stat. Science 1994, 265, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Mikita, T.; Campbell, D.; Wu, P.; Williamson, K.; Schindler, U. Requirements for interleukin-4-induced gene expression and functional characterization of Stat6. Mol. Cell. Biol. 1996, 16, 5811–5820. [Google Scholar] [CrossRef] [PubMed]
- Schindler, U.; Wu, P.; Rothe, M.; Brasseur, M.; McKnight, S.L. Components of a Stat recognition code: Evidence for two layers of molecular selectivity. Immunity 1995, 2, 689–697. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Shukla, M.; Yakubenko, V.P.; Mulya, A.; Kundu, S.; Cathcart, M.K. IL-4 and IL-13 employ discrete signaling pathways for target gene expression in alternatively activated monocytes/macrophages. Free Radic. Biol. Med. 2013, 54, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.E., Jr.; Kerr, I.M.; Stark, G.R. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.; Fu, P.; Levine, A.D. REDOX regulation of IL-13 signaling in intestinal epithelial cells: Usage of alternate pathways mediates distinct gene expression patterns. Cell. Signal. 2010, 22, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Bluthe, R.M.; Bristow, A.; Lestage, J.; Imbs, C.; Dantzer, R. Central injection of interleukin-13 potentiates LPS-induced sickness behavior in rats. Neuroreport 2001, 12, 3979–3983. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.H.; Lee, D.Y.; Park, K.W.; Kim, S.U.; Yang, M.S.; Joe, E.H.; Jin, B.K. Microglia expressing interleukin-13 undergo cell death and contribute to neuronal survival in vivo. Glia 2004, 46, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-S.; Ji, K.-A.; Jeon, S.-B.; Jin, B.-K.; Kim, S.U.; Jou, N.; Joe, E. Interleukin-13 enhances cyclooxygenase-2 expression in activated rat brain microglia: Implications for death of activated microglia. J. Immunol. 2006, 177, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.S.; Park, E.J.; Sohn, S.; Kwon, H.J.; Shin, W.H.; Pyo, H.K.; Jin, B.; Choi, K.S.; Jou, I.; Joe, E.H. Interleukin-13 and -4 induce death of activated microglia. Glia 2002, 38, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, Y.; Shimoda, T.; Uno, K.; Tateishi, N.; Furuya, S.; Yagi, K.; Suzuki, K.; Fujita, S. The effects of MPTP on the activation of microglia/astrocytes and cytokine/chemokine levels in different mice strains. J. Neuroimmunol. 2008, 204, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.T.; Lee, C.H.; Yoo, K.Y.; Choi, J.H.; Li, H.; Park, O.K.; Yan, B.; Hwang, I.K.; Kwon, Y.G.; Kim, Y.M.; et al. Maintenance of anti-inflammatory cytokines and reduction of glial activation in the ischemic hippocampal CA1 region preconditioned with lipopolysaccharide. J. Neurol. Sci. 2010, 296, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, H.; Sun, G.; Zhang, J.; Edwards, N.J.; Aronowski, J. Neuronal interleukin-4 as a modulator of microglial pathways and ischemic brain damage. J. Neurosci. 2015, 35, 11281–11291. [Google Scholar] [CrossRef] [PubMed]
- Latta, C.H.; Sudduth, T.L.; Weekman, E.M.; Brothers, H.M.; Abner, E.L.; Popa, G.J.; Mendenhall, M.D.; Gonzalez-Oregon, F.; Braun, K.; Wilcock, D.M. Determining the role of IL-4 induced neuroinflammation in microglial activity and amyloid-β using BV2 microglial cells and APP/PS1 transgenic mice. J. Neuroinflamm. 2015, 12, 41. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Xu, L.; Wei, L.; White, R.E.; Ouyang, Y.B.; Giffard, R.G. IL-4 is required for sex differences in vulnerability to focal ischemia in mice. Stroke 2015, 46, 2271–2276. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Park, H.J.; Starkweather, A.; An, K.; Shim, I. Decreased interleukin-4 release from the neurons of the Locus Coeruleus in response to immobilization stress. Med. Inflamm. 2016, 2016, 3501905. [Google Scholar] [CrossRef] [PubMed]
- Conti, B.; The Scripps Research Institute, CA, USA. Unpublished work. 2016.
- Nam, J.H.; Park, K.W.; Park, E.S.; Lee, Y.B.; Lee, H.G.; Baik, H.H.; Kim, Y.S.; Maeng, S.; Park, J.; Jin, B.K. Interleukin-13/-4-induced oxidative stress contributes to death of hippocampal neurons in aβ1-42-treated hippocampus in vivo. Antioxid. Redox Signal. 2012, 16, 1369–1383. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Baik, H.H.; Jin, B.K. Interleukin-4-induced oxidative stress via microglial NADPH oxidase contributes to the death of hippocampal neurons in vivo. Curr. Aging Sci. 2008, 1, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Baik, H.H.; Jin, B.K. IL-13-induced oxidative stress via microglial NADPH oxidase contributes to death of hippocampal neurons in vivo. J. Immunol. 2009, 183, 4666–4674. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.C.; Burudi, E.M.; Alirezaei, M.; Flynn, C.C.; Watry, D.D.; Lanigan, C.M.; Fox, H.S. IFN-gamma-induced IDO and WRS expression in microglia is differentially regulated by IL-4. Glia 2007, 55, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.M.; Lyons, A.; O’Connell, F.; Deighan, B.F.; Barry, C.E.; Anyakoha, N.G.; Nicolaou, A.; Lynch, M.A. A pivotal role for interleukin-4 in atorvastatin-associated neuroprotection in rat brain. J. Biol. Chem. 2008, 283, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Deboy, C.A.; Xin, J.; Byram, S.C.; Serpe, C.J.; Sanders, V.M.; Jones, K.J. Immune-mediated neuroprotection of axotomized mouse facial motoneurons is dependent on the IL-4/STAT6 signaling pathway in CD4(+) T cells. Exp. Neurol. 2006, 201, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Mancino, R.; Bergami, A.; Mori, F.; Castelli, M.; De Chiara, V.; Studer, V.; Mataluni, G.; Sancesario, G.; Parisi, V.; et al. Potential role of IL-13 in neuroprotection and cortical excitability regulation in multiple sclerosis. Mult. Scler. J. 2011, 17, 1301–1312. [Google Scholar] [CrossRef] [PubMed]
- Won, S.Y.; Kim, S.R.; Maeng, S.; Jin, B.K. Interleukin-13/Interleukin-4-induced oxidative stress contributes to death of prothrombinkringle-2 (pKr-2)-activated microglia. J. Neuroimmunol. 2013, 265, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Bluthe, R.M.; Lestage, J.; Rees, G.; Bristow, A.; Dantzer, R. Dual effect of central injection of recombinant rat interleukin-4 on lipopolysaccharide-induced sickness behavior in rats. Neuropsychopharmacology 2002, 26, 86–93. [Google Scholar] [CrossRef]
- Linker, R.A.; Sendtner, M.; Gold, R. Mechanisms of axonal degeneration in EAE—Lessons from CNTF and MHC I knockout mice. J. Neurol. Sci. 2005, 233, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Zeis, T.; Graumann, U.; Reynolds, R.; Schaeren-Wiemers, N. Normal-appearing white matter in multiple sclerosis is in a subtle balance between inflammation and neuroprotection. Brain 2008, 131, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Seyfizadeh, N.; Kazemi, T.; Farhoudi, M.; Reza Aliparasti, M.; Sadeghi-Bazargani, H.; Almasi, S.; Babaloo, Z. Association of IL-13 single nucleotide polymorphisms in Iranian patients to multiple sclerosis. Am. J. Clin. Exp. Immunol. 2014, 3, 124–129. [Google Scholar] [PubMed]
- Rozsa, K.S.; Rubakhin, S.S.; Szucs, A.; Hughes, T.K.; Stefano, G.B. Opposite effects of interleukin-2 and interleukin-4 on GABA-induced inward currents of dialysed lymnaea neurons. Gen. Pharmacol. 1997, 29, 73–77. [Google Scholar] [CrossRef]
- Sanna, A.; Fois, M.L.; Arru, G.; Huang, Y.M.; Link, H.; Pugliatti, M.; Rosati, G.; Sotgiu, S. Glatiramer acetate reduces lymphocyte proliferation and enhances IL-5 and IL-13 production through modulation of monocyte-derived dendritic cells in multiple sclerosis. Clin. Exp. Immunol. 2006, 143, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Cash, E.; Minty, A.; Ferrara, P.; Caput, D.; Fradelizi, D.; Rott, O. Macrophage-inactivating IL-13 suppresses experimental autoimmune encephalomyelitis in rats. J. Immunol. 1994, 153, 4258–4267. [Google Scholar] [PubMed]
- Offner, H.; Subramanian, S.; Wang, C.; Afentoulis, M.; Vandenbark, A.A.; Huan, J.; Burrows, G.G. Treatment of passive experimental autoimmune encephalomyelitis in SJL mice with a recombinant TCR ligand induces IL-13 and prevents axonal injury. J. Immunol. 2005, 175, 4103–4111. [Google Scholar] [CrossRef] [PubMed]
- Young, D.A.; Lowe, L.D.; Booth, S.S.; Whitters, M.J.; Nicholson, L.; Kuchroo, V.K.; Collins, M. IL-4, IL-10, IL-13, and TGF-β from an altered peptide ligand-specific Th2 cell clone down-regulate adoptive transfer of experimental autoimmune encephalomyelitis. J. Immunol. 2000, 164, 3563–3572. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Repáraz, J.; Rynda, A.; Ascón, M.A.; Yang, X.; Kochetkova, I.; Riccardi, C.; Callis, G.; Trunkle, T.; Pascual, D.W. IL-13 production by regulatory T cells protects against experimental autoimmune encephalomyelitis independently of autoantigen. J. Immunol. 2008, 181, 954–968. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Sinha, S.; Kaler, L.J.; Proctor, T.M.; Teuscher, C.; Vandenbark, A.A.; Offner, H. IL-13-mediated gender difference in susceptibility to autoimmune encephalomyelitis. J. Immunol. (Baltim. Md. 1950) 2008, 180, 2679–2685. [Google Scholar] [CrossRef]
- Verdu, E.F.; Deng, Y.; Bercik, P.; Collins, S.M. Modulatory effects of estrogen in two murine models of experimental colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, K.; Suenobu, M.; Yoshida, A.; Koga, K.; Hyodo, A.; Ohtsuka, H.; Kuniyasu, A.; Tamamaki, N.; Sugimoto, Y.; Nakayama, H. Intracerebral microinjection of interleukin-4/interleukin-13 reduces β-amyloid accumulation in the ipsilateral side and improves cognitive deficits in young amyloid precursor protein 23 mice. Neuroscience 2012, 207, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Man (OMIM) database. Available online: http://www.omim.org (accessed on 9 June 2016).
- Frank-Cannon, T.C.; Tran, T.; Ruhn, K.A.; Martinez, T.N.; Hong, J.; Marvin, M.; Hartley, M.; Trevino, I.; O’Brien, D.E.; Casey, B.; et al. Parkin deficiency increases vulnerability to inflammation-related nigral degeneration. J. Neurosci. 2008, 28, 10825–10834. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mori, S.; Maher, P.; Conti, B. Neuroimmunology of the Interleukins 13 and 4. Brain Sci. 2016, 6, 18. https://doi.org/10.3390/brainsci6020018
Mori S, Maher P, Conti B. Neuroimmunology of the Interleukins 13 and 4. Brain Sciences. 2016; 6(2):18. https://doi.org/10.3390/brainsci6020018
Chicago/Turabian StyleMori, Simone, Pamela Maher, and Bruno Conti. 2016. "Neuroimmunology of the Interleukins 13 and 4" Brain Sciences 6, no. 2: 18. https://doi.org/10.3390/brainsci6020018
APA StyleMori, S., Maher, P., & Conti, B. (2016). Neuroimmunology of the Interleukins 13 and 4. Brain Sciences, 6(2), 18. https://doi.org/10.3390/brainsci6020018