Imagery-Mediated Verbal Learning Depends on Vividness–Familiarity Interactions: The Possible Role of Dualistic Resting State Network Activity Interference


Abstract
:1. Introduction
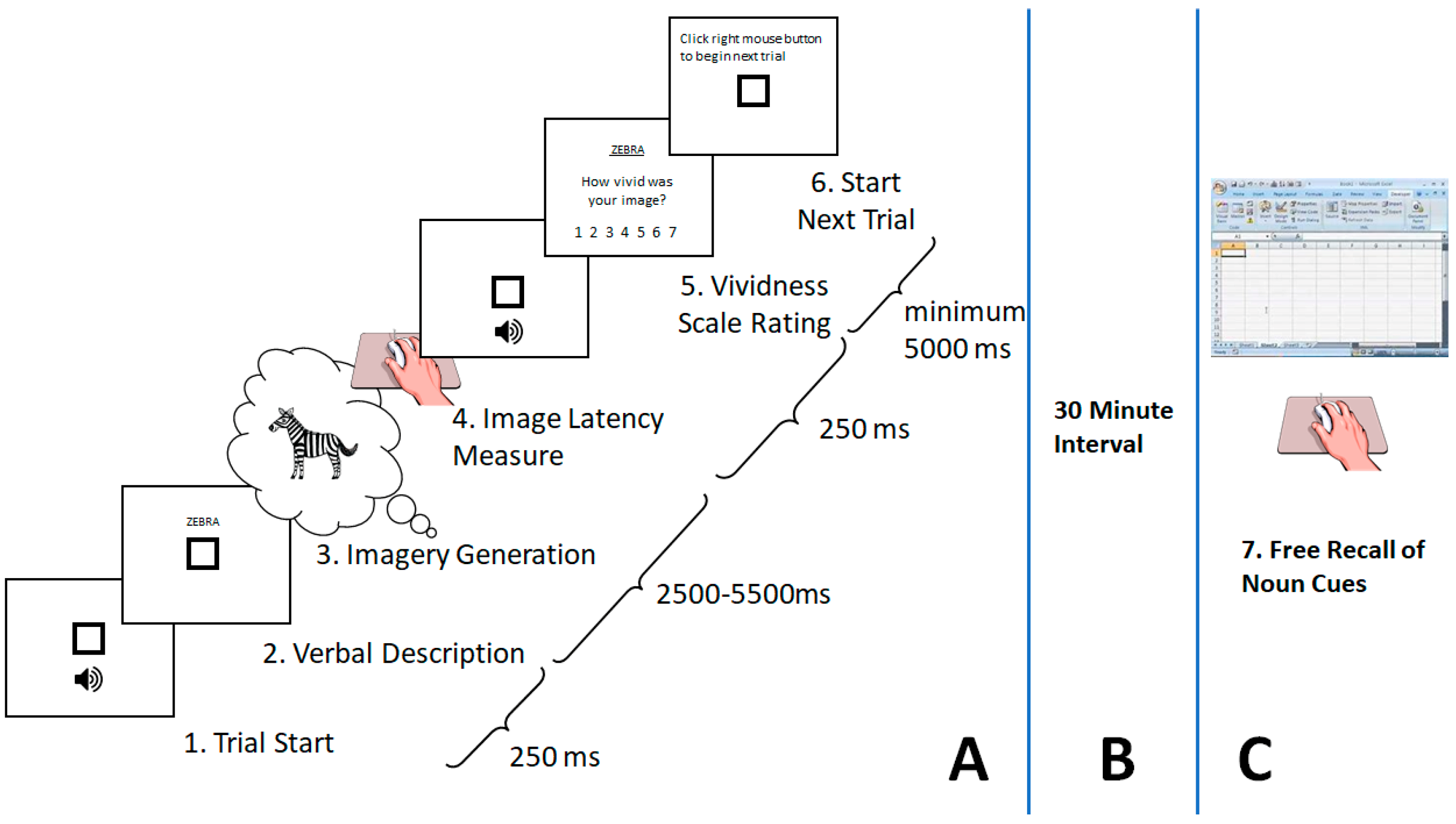
2. Method
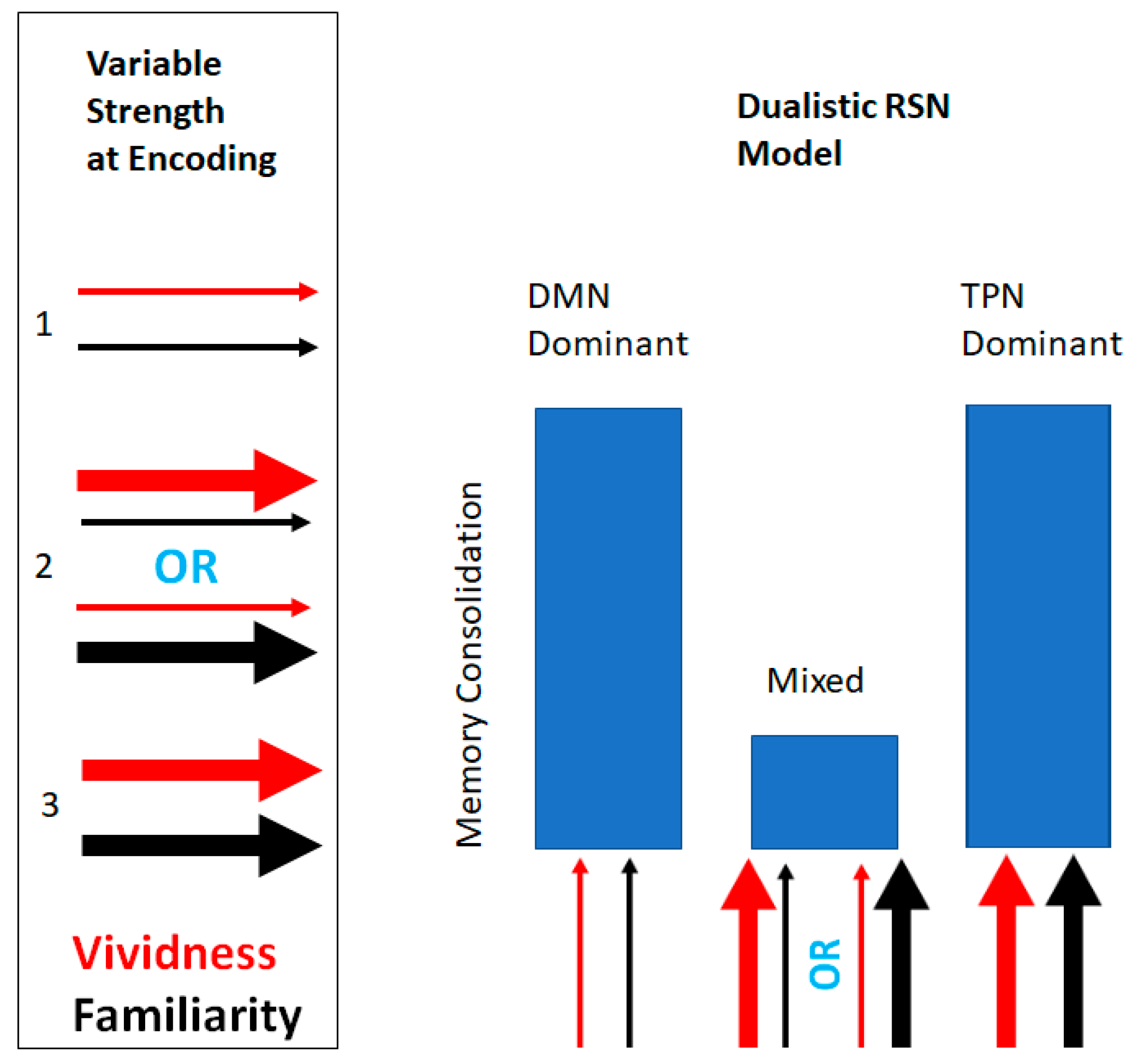
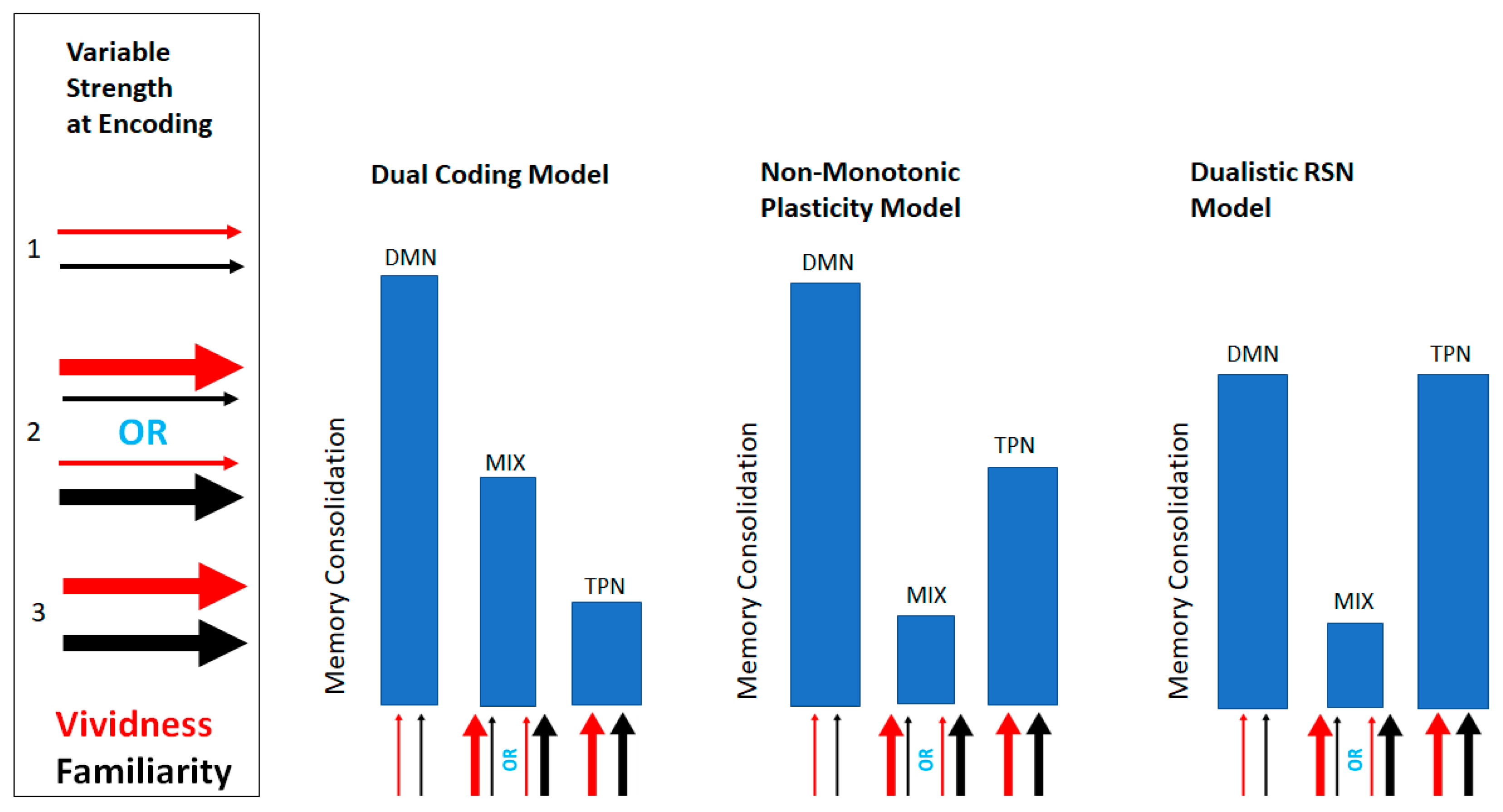
2.1. Design and Analytic Strategy
2.2. Stimuli
2.2.1. Initial Stimulus Selection
2.2.2. Vividness Rating Procedure
Sample of Vividness Raters
Image Generation Phase
Stimulus Familiarity Matching and Diagnostic Procedure
Filler Stimuli
2.3. Dependent Variables
2.3.1. Image Latency Times Measure
2.3.2. Free Incidental Recall Measure
3. Results
3.1. Stimuli Characteristics: Descriptive and Correlations
3.2. Mental Imagery Vividness and Familiarity
3.3. Mental Imagery Vividness and Reaction Time
3.4. Effects of Resting State Networks on Learning Outcomes
3.4.1. By-Item Analysis Approach
3.4.2. Linear Mixed Logistic Regression Approach
3.5. Effects of Resting State Networks on Mental Imagery Latency (Reaction Time)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Tulving, E.; McNulty, J.A.; Ozier, M. Vividness of words and learning in free-recall learning. Can. J. Psychol. Can. Psychol. 1965, 19, 242–252. [Google Scholar] [CrossRef]
- Baddeley, A.D.; Andrade, J. Working memory and the vividness of imagery. J. Exp. Psychol. Gen. 2000, 129, 126–145. [Google Scholar] [CrossRef] [PubMed]
- D’Angiulli, A.; Runge, M.; Faulkner, A.; Zakizadeh, J.; Chan, A.; Morcos, S. Vividness of Visual Imagery and Incidental Recall of Verbal Cues, When Phenomenological Availability Reflects Long-Term Memory Accessibility. Front. Psychol. 2013, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, P.W. The Role of Imagery in Incidental Learning. Br. J. Psychol. 1971, 62, 235–243. [Google Scholar] [CrossRef]
- Yonelinas, A.P. The Nature of Recollection and Familiarity: A Review of 30 Years of Research. J. Mem. Lang. 2002, 46, 441–517. [Google Scholar] [CrossRef] [Green Version]
- Mandler, G. Familiarity Breeds Attempts: A Critical Review of Dual-Process Theories of Recognition. Perspect. Psychol. Sci. 2008, 3, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellezza, F.S. Factors that affect vividness ratings. J. Ment. Imag. 1995, 19, 123–129. [Google Scholar]
- Paivio, A. Mental Representations: A Dual Coding Approach; Oxford University Press: New York, NY, USA, 1990; Volume 10. [Google Scholar]
- Newman, E.L.; Norman, K.A. Moderate Excitation Leads to Weakening of Perceptual Representations. Cereb. Cortex 2010, 20, 2760–2770. [Google Scholar] [CrossRef] [Green Version]
- Lewis-Peacock, J.A.; Norman, K.A. Competition between items in working memory leads to forgetting. Nat. Commun. 2014, 5, 5768. [Google Scholar] [CrossRef] [Green Version]
- Huijbers, W.; Schultz, A.P.; Vannini, P.; McLaren, D.G.; Wigman, S.E.; Ward, A.M.; Hedden, T.; Sperling, R.A. The encoding/retrieval flip: Interactions between memory performance and memory stage and relationship to intrinsic cortical networks. J. Cogn. Neurosci. 2013, 25, 1163–1179. [Google Scholar] [CrossRef]
- Kim, H. Encoding and retrieval along the long axis of the hippocampus and their relationships with dorsal attention and default mode networks: The HERNET model. Hippocampus 2015, 25, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Block, N. Two neural correlates of consciousness. Trends Cogn. Sci. 2005, 9, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Fox, M.D.; Greicius, M. Clinical Applications of Resting State Functional Connectivity. Front. Syst. Neurosci. 2010, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M. Resting-state functional connectivity in neuropsychiatric disorders. Curr. Opin. Neurol. 2008, 24, 424–430. [Google Scholar] [CrossRef]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Corbetta, M.; Van Essen, D.C.; Raichle, M.E. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. USA 2005, 102, 9673–9678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smallwood, J.; Spreng, R.N.; Andrews-Hanna, J.R.; Andrews-Hanna, J.R. The default network and self-generated thought: Component processes, dynamic control, and clinical relevance. Ann. N. Y. Acad. Sci. 2014, 1316, 29–52. [Google Scholar]
- Raichle, M.E. The Brain’s Default Mode Network. Ann. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef]
- Mantini, D.; Perrucci, M.G.; Del Gratta, C.; Romani, G.L.; Corbetta, M. Electrophysiological signatures of resting state networks in the human brain. Proc. Natl. Acad. Sci. USA 2007, 104, 13170–13175. [Google Scholar] [CrossRef] [Green Version]
- Kompus, K. Default mode network gates the retrieval of task-irrelevant incidental memories. Neurosci. Lett. 2011, 487, 318–321. [Google Scholar] [CrossRef]
- Dolcos, F.; LaBar, K.S.; Cabeza, R. Remembering one year later: Role of the amygdala and the medial temporal lobe memory system in retrieving emotional memories. Proc. Natl. Acad. Sci. USA 2005, 102, 2626–2631. [Google Scholar] [CrossRef] [Green Version]
- Diana, R.A.; Yonelinas, A.P.; Ranganath, C. Imaging recollection and familiarity in the medial temporal lobe: A three-component model. Trends Cogn. Sci. 2007, 11, 379–386. [Google Scholar] [CrossRef] [PubMed]
- St-Laurent, M.; Abdi, H.; Buchsbaum, B.R. Distributed Patterns of Reactivation Predict Vividness of Recollection. J. Cogn. Neurosci. 2015, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Weissman, D.H.; Roberts, K.C.; Visscher, K.M.; Woldorff, M.G.; Woldorff, M. The neural bases of momentary lapses in attention. Nat. Neurosci. 2006, 9, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Mowinckel, A.M.; Alnæs, D.; Pedersen, M.L.; Ziegler, S.; Fredriksen, M.; Kaufmann, T.; Sonuga-Barke, E.; Endestad, T.; Westlye, L.T.; Biele, G. Increased default-mode variability is related to reduced task-performance and is evident in adults with ADHD. NeuroImage Clin. 2017, 16, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q.; Kelly, A.C.; Biswal, B.B.; Margulies, D.S.; Shehzad, Z.; Shaw, D.; Ghaffari, M.; Rotrosen, J.; Adler, L.A.; Castellanos, F.X.; et al. Network homogeneity reveals decreased integrity of default-mode network in ADHD. J. Neurosci. Methods 2008, 169, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Assaf, M.; Jagannathan, K.; Calhoun, V.D.; Miller, L.; Stevens, M.C.; Sahl, R.; O’Boyle, J.G.; Schultz, R.T.; Pearlson, G.D. Abnormal functional connectivity of default mode sub-networks in autism spectrum disorder patients. NeuroImage 2010, 53, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Sheline, Y.I.; Barch, D.M.; Price, J.L.; Rundle, M.M.; Vaishnavi, S.N.; Snyder, A.Z.; Mintun, M.A.; Wang, S.; Coalson, R.S.; Raichle, M.E. The default mode network and self-referential processes in depression. Proc. Natl. Acad. Sci. USA 2009, 106, 1942–1947. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.-H.; Wang, P.-J.; Li, C.-B.; Hu, Z.-H.; Xi, Q.; Wu, W.-Y.; Tang, X.-W. Altered default mode network activity in patient with anxiety disorders: An fMRI study. Eur. J. Radiol. 2007, 63, 373–378. [Google Scholar] [CrossRef]
- Liao, W.; Zhang, Z.; Pan, Z.; Mantini, D.; Ding, J.; Duan, X.; Luo, C.; Wang, Z.; Tan, Q.; Lu, G.; et al. Default mode network abnormalities in mesial temporal lobe epilepsy: A study combining fMRI and DTI. Hum. Brain Mapp. 2011, 32, 883–895. [Google Scholar] [CrossRef]
- Bastos-Leite, A.J.; Ridgway, G.R.; Silveira, C.; Norton, A.; Reis, S.; Friston, K.J. Dysconnectivity within the default mode in first-episode schizophrenia: A stochastic dynamic causal modeling study with functional magnetic resonance imaging. Schizophr. Bull. 2015, 41, 144–153. [Google Scholar] [CrossRef]
- Camchong, J.; MacDonald, A.W.; Bell, C.; Mueller, B.A.; Lim, K.O. Altered functional and anatomical connectivity in schizophrenia. Schizophr. Bull. 2011, 37, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Pomarol-Clotet, E.; Salvador, R.; Sarró, S.; Gomar, J.; Vila, F.; Martínez, A.; Guerrero, A.; Ortiz-Gil, J.; Sans-Sansa, B.; Capdevila, A.; et al. Failure to deactivate in the prefrontal cortex in schizophrenia: Dysfunction of the default mode network? Psychol. Med. 2008, 38, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Garrity, A.G.; Pearlson, G.D.; McKiernan, K.; Lloyd, D.; Kiehl, K.A.; Calhoun, V.D. Aberrant “Default Mode” Functional Connectivity in Schizophrenia. Am. J. Psychiatry 2007, 164, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Sheline, Y.I.; Raichle, M.E. Resting State Functional Connectivity in Preclinical Alzheimer’s Disease: A Review. Boil. Psychiatry 2013, 74, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Sorg, C.; Riedl, V.; Mühlau, M.; Calhoun, V.D.; Eichele, T.; Läer, L.; Drzezga, A.; Förstl, H.; Kurz, A.; Zimmer, C.; et al. Selective changes of resting-state networks in individuals at risk for Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 18760–18765. [Google Scholar] [CrossRef] [PubMed]
- Glahn, D.C.; Winkler, A.M.; Kochunov, P.; Almasy, L.; Duggirala, R.; Carless, M.A.; Curran, J.C.; Olvera, R.L.; Laird, A.R.; Smith, S.M.; et al. Genetic control over the resting brain. Proc. Natl. Acad. Sci. USA 2010, 107, 1223–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulford, J.; Milton, F.; Salas, D.; Smith, A.; Simler, A.; Winlove, C.; Zeman, A. The neural correlates of visual imagery vividness—An fMRI study and literature review. Cortex 2018, 105, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Dissociating the roles of the default-mode, dorsal, and ventral networks in episodic memory retrieval. NeuroImage 2010, 50, 1648–1657. [Google Scholar] [CrossRef] [PubMed]
- Belardinelli, M.O.; Palmiero, M.; Sestieri, C.; Nardo, D.; Di Matteo, R.; Londei, A.; D’Ausilio, A.; Ferretti, A.; Del Gratta, C.; Romani, G. An fMRI investigation on image generation in different sensory modalities: The influence of vividness. Acta Psychol. 2009, 132, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, A.; Montaldi, D. Two separate, but interacting, neural systems for familiarity and novelty detection: A dual-route mechanism. Hippocampus 2014, 24, 516–527. [Google Scholar] [CrossRef]
- Bedny, M.; Aguirre, G.K.; Thompson-Schill, S.L. Item analysis in functional magnetic resonance imaging. NeuroImage 2007, 35, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, S.; Stadler, M.; Lindl, A.; Naumann, F.; Bühner, M. Analyzing longitudinal intervention studies with linear mixed models. TPM Test. Psychometr. Method. Appl. Psychol. 2019, 26, 101–119. [Google Scholar]
- Mental Imagery Data from Words. Available online: https://zenodo.org/record/2658726#.XPptDsTQiUl (accessed on 5 February 2019).
- D’Angiulli, A. Is the Spotlight an Obsolete Metaphor of “Seeing with the Mind’s Eye”? A Constructive Naturalistic Approach to the Inspection of Visual Mental Images. Imagin. Cogn. Personal. 2009, 28, 117–135. [Google Scholar] [CrossRef]
- D’Angiulli, A.; Reeves, A. Probing Vividness Increment through Imagery Interruption. Imagin. Cogn. Personal. 2003, 23, 63–78. [Google Scholar] [CrossRef]
- D’Angiulli, A. Mental image generation and the contrast sensitivity function. Cognition 2002, 85, B11–B19. [Google Scholar] [CrossRef]
- D’Angiulli, A.; Reeves, A. The Relationship between Self-Reported Vividness and Latency during Mental Size Scaling of Everyday Items: Phenomenological Evidence of Different Types of Imagery. Am. J. Psychol. 2007, 120, 521. [Google Scholar] [CrossRef]
- MRC Psycholinguistic Database. Available online: http://websites.psychology.uwa.edu.au/school/MRCDatabase/uwa_mrc.htm (accessed on 13 April 2013).
- D’Angiulli, A. Dissociating Vividness and Imageability. Imagin. Cogn. Personal. 2003, 23, 79–88. [Google Scholar] [CrossRef]
- Paivio, A. Imagery and Verbal Processes; Holt, Rinehart & Winston: Oxford, UK, 1971. [Google Scholar]
- Miller, L.M.; Roodenrys, S. The interaction of word frequency and concreteness in immediate serial recall. Mem. Cogn. 2009, 37, 850–865. [Google Scholar] [CrossRef] [Green Version]
- Tse, C.-S.; Altarriba, J. Testing the associative-link hypothesis in immediate serial recall: Evidence from word frequency and word imageability effects. Memory 2007, 15, 675–690. [Google Scholar] [CrossRef]
- Clark, C. MRC Psycholinguistic Database; The University of Western Australia School of Psychology: Perth, Australia, 1997. [Google Scholar]
- Ma, W.; Golinkoff, R.M.; Hirsh-Pasek, K.; McDonough, C.; Tardif, T. Imageability predicts the age of acquisition of verbs in Chinese children. J. Child Lang. 2009, 36, 405–423. [Google Scholar] [CrossRef]
- Barry, C.; Gerhand, S. Both concreteness and age-of-acquisition affect reading accuracy but only concreteness affects comprehension in a deep dyslexic patient. Brain Lang. 2003, 84, 84–104. [Google Scholar] [CrossRef]
- Ethical Conduct for Research Involving Humans. Available online: http://www.pre.ethics.gc.ca/pdf/eng/tcps2-2014/TCPS_2_FINAL_Web.pdf (accessed on 15 Decemeber 2014).
- Marks, D.F. New directions for mental imagery research. J. Ment. Imag. 1995, 19, 153–167. [Google Scholar]
- Wilson, M. MRC psycholinguistic database: Machine-usable dictionary, version 2.00. Behav. Res. Methods Instrum. Comput. 1988, 20, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Paivio, A.; Yuille, J.C.; Madigan, S.A. Concreteness, imagery, and meaningfulness values for 925 nouns. J. Exp. Psychol. 1968, 76, 1–25. [Google Scholar] [CrossRef]
- Toglia, M.P.; Battig, W.F. Handbook of Semantic Word Norms; Hillsdale, N.J., Ed.; Lawrence Erlbaum Associates: New York, NY, USA, 1978; ISBN 978-0-470-26378-5. [Google Scholar]
- Gilhooly, K.J.; Logie, R.H.; Logie, R. Age-of-acquisition, imagery, concreteness, familiarity, and ambiguity measures for 1944 words. Behav. Res. Methods 1980, 12, 395–427. [Google Scholar] [CrossRef]
- Coltheart, M. The MRC Psycholinguistic Database. Quart. J. Exper. Psychol. Sect. A 1981, 33, 497–505. [Google Scholar] [CrossRef]
- Craver-Lemley, C.; Reeves, A. Visual Imagery Selectively Reduces Vernier Acuity. Perception 1987, 16, 599–614. [Google Scholar] [CrossRef]
- D’Angiulli, A.; Reeves, A. Generating visual mental images: Latency and vividness are inversely related. Mem. Cogn. 2002, 30, 1179–1188. [Google Scholar] [CrossRef] [Green Version]
- Preacher, K.J.; Rucker, D.D.; Maccallum, R.C.; Nicewander, W.A. Use of the Extreme Groups Approach: A Critical Reexamination and New Recommendations. Psychol. Methods 2005, 10, 178–192. [Google Scholar] [CrossRef]
- Rosenthal, R.; Robert, R.; Rosnow, R.L. Contrast Analysis: Focused Comparisons in the Analysis of Variance; Cambridge University Press: Cambridge, UK, 1985. [Google Scholar]
- Hays, W. Statistics, 5th ed.; Wadsworth Publishing: Fort Worth, TX, USA, 1994; ISBN 978-0-03-074467-9. [Google Scholar]
- Meier, U. A note on the power of Fisher’s least significant difference procedure. Pharm. Stat. 2006, 5, 253–263. [Google Scholar] [CrossRef]
- Magezi, D.A. Linear mixed-effects models for within-participant psychology experiments: An introductory tutorial and free, graphical user interface (LMMgui). Front. Psychol. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Lei, X. Competition between frontoparietal control and default networks supports social working memory and empathy. Soc. Cogn. Affect. Neurosci. 2015, 10, 1144–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehm, L.P. Relationships among measures of visual imagery. Behav. Res. Ther. 1973, 11, 265–270. [Google Scholar] [CrossRef]
- Hale, S.M.; Simpson, H.M. Effects of eye movements on the rate of discovery and the vividness of visual images. Percept. Psychophys. 1971, 9, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Van Buuren, M.; Vink, M.; Kahn, R.S. Default-mode network dysfunction and self-referential processing in healthy siblings of schizophrenia patients. Schizophr. Res. 2012, 142, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield-Gabrieli, S.; Ford, J.M. Default Mode Network Activity and Connectivity in Psychopathology. Annu. Rev. Clin. Psychol. 2012, 8, 49–76. [Google Scholar] [CrossRef] [PubMed]
- Bossong, M.G.; Jansma, J.M.; van Hell, H.H.; Jager, G.; Kahn, R.S.; Ramsey, N.F. Default Mode Network in the Effects of Δ9-Tetrahydrocannabinol (THC) on Human Executive Function. PLoS ONE 2013, 8, e70074. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | M | SD | Min | Max |
|---|---|---|---|---|
| Vividness mean | 5.26 | 0.52 | 3.99 | 6.12 |
| RTs (ms) | 3892.59 | 524.24 | 2692.87 | 5271.39 |
| Familiarity mean | 5.60 | 0.74 | 3.82 | 6.84 |
| Recall probability | 0.40 | 0.22 | 0.08 | 0.9231 |
| Variables | Vividness | Reaction Time | Familiarity | Recall Probability |
|---|---|---|---|---|
| Vividness | −0.39 ** | 0.48 ** | −0.16 | |
| Reaction time | −0.39 ** | 0.04 | −0.01 | |
| Familiarity | 0.48 ** | 0.04 | 0.01 | |
| Recall Probability | −0.16 | −0.01 | 0.01 |
| Descriptive | |||||
|---|---|---|---|---|---|
| Variables | N | M | SE | Min | Max |
| TPN dominant | 18 | 0.40 | 0.06 | 0.08 | 0.92 |
| Mixed | 14 | 0.27 | 0.04 | 0.08 | 0.58 |
| DMN dominant | 18 | 0.49 | 0.04 | 0.15 | 0.77 |
| Variables | DRSNAP | ||||
|---|---|---|---|---|---|
| Variable Levels | TPN | Mixed | DMN | Total | |
| High | 0.06 (3) | 0 | 0.1 (5) | 0.16 (8) | |
| Recall | Medium | 0.14 (7) | 0.04 (2) | 0.18 (9) | 0.36 (18) |
| Low | 0.16 (8) | 0.24 (12) | 0.08 (4) | 0.48 (24) | |
| Total | 0.36 (18) | 0.28 (14) | 0.36 (18) | 1.00 (50) |
| Descriptive | |||||
|---|---|---|---|---|---|
| Variables | N | M | SE | 95% CI | 95% CI |
| TPN dominant | 519 | 0.36 | 0.02 | 0.32 | 0.40 |
| Mixed | 528 | 0.29 | 0.02 | 0.25 | 0.33 |
| DMN dominant | 253 | 0.36 | 0.03 | 0.30 | 0.42 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lefebvre, E.; D’Angiulli, A. Imagery-Mediated Verbal Learning Depends on Vividness–Familiarity Interactions: The Possible Role of Dualistic Resting State Network Activity Interference. Brain Sci. 2019, 9, 143. https://doi.org/10.3390/brainsci9060143
Lefebvre E, D’Angiulli A. Imagery-Mediated Verbal Learning Depends on Vividness–Familiarity Interactions: The Possible Role of Dualistic Resting State Network Activity Interference. Brain Sciences. 2019; 9(6):143. https://doi.org/10.3390/brainsci9060143
Chicago/Turabian StyleLefebvre, Etienne, and Amedeo D’Angiulli. 2019. "Imagery-Mediated Verbal Learning Depends on Vividness–Familiarity Interactions: The Possible Role of Dualistic Resting State Network Activity Interference" Brain Sciences 9, no. 6: 143. https://doi.org/10.3390/brainsci9060143