
Quercus acuta Thunb. (Fagaceae) and Its Component, Isoquercitrin, Inhibit HSV-1 Replication by Suppressing Virus-Induced ROS Production and NF-κB Activation

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Cultures and Viruses
2.3. Cell Viability by Cell Counting Kit-8 (CCK-8) Assay
2.4. Analysis of HSV-1 GFP Expression
2.5. Plaque Reduction Assay
2.6. Protein Detection by Western Blot Analysis
2.7. mRNA Measurement by Real-Time PCR
2.8. Measurement of the Accumulation of ROS
2.9. NF-κB Nuclear Localization for Immunofluorescence Staining
2.10. NF-κB DNA-Binding Activity by Electrophoretic Mobility Shift Assay (EMSA)
2.11. Ultrahigh-Performance Liquid Chromatography (UPLC) Coupled to High-Resolution Orbitrap Mass Spectrometry
2.12. Modeling of ICP27 Structure
2.13. Molecular Docking Simulation and Pharmacophore Analysis
2.14. Statistical Analysis
3. Results
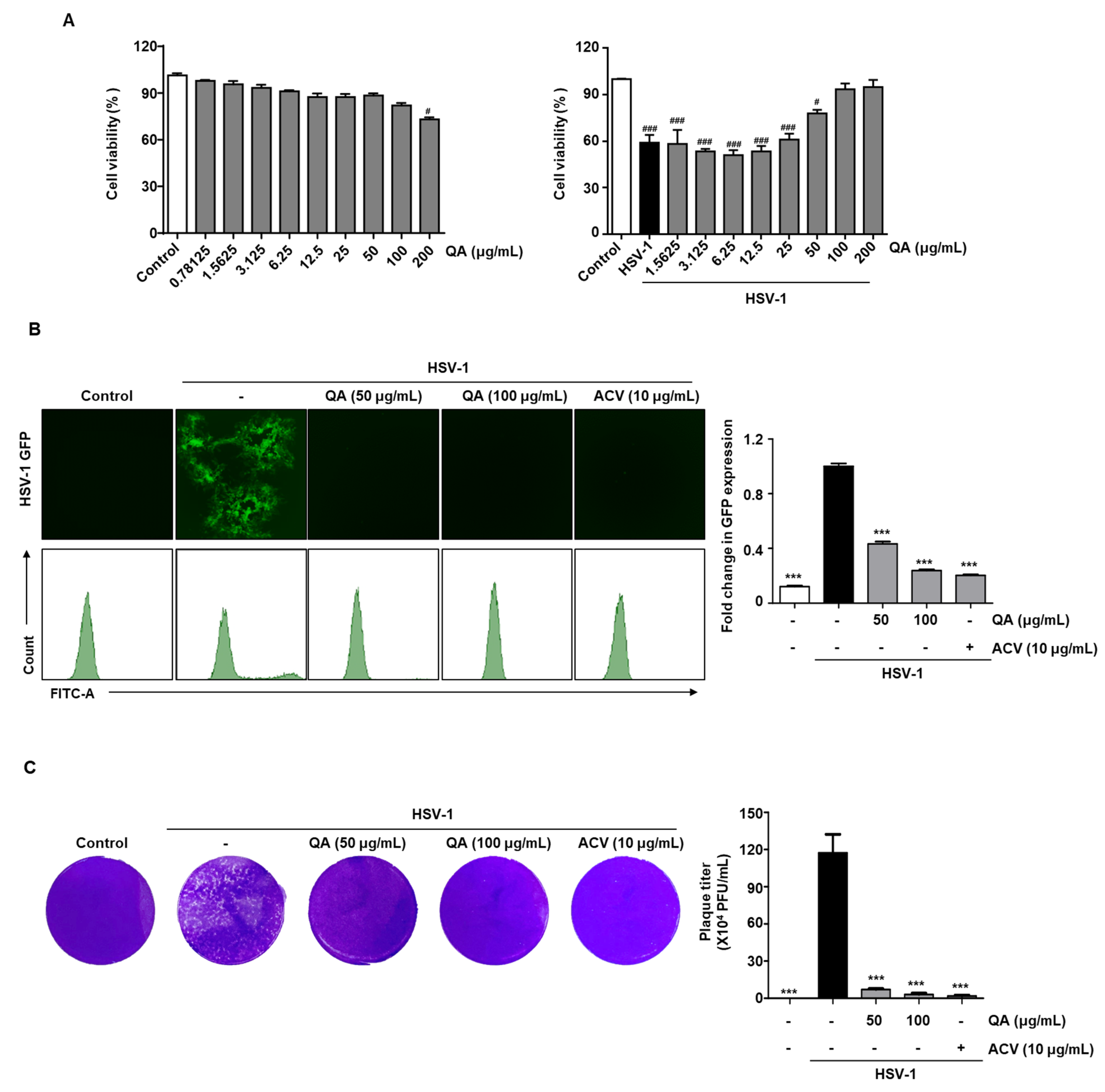
3.1. QA Exhibits Antiviral Effects upon Infection with HSV-1 in Vero Cells
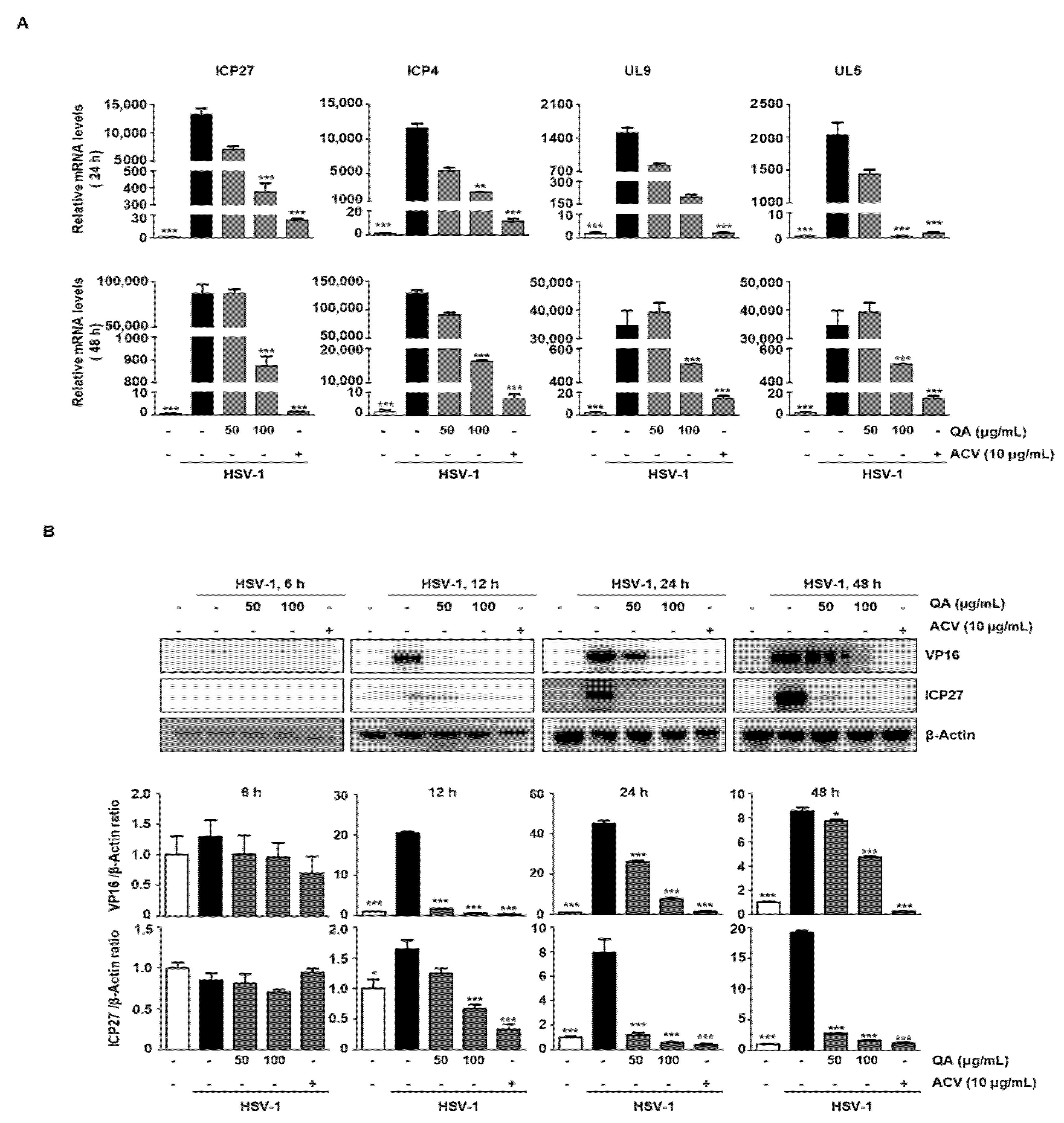
3.2. QA Reduces mRNA and Protein Levels of HSV-1 Related Genes
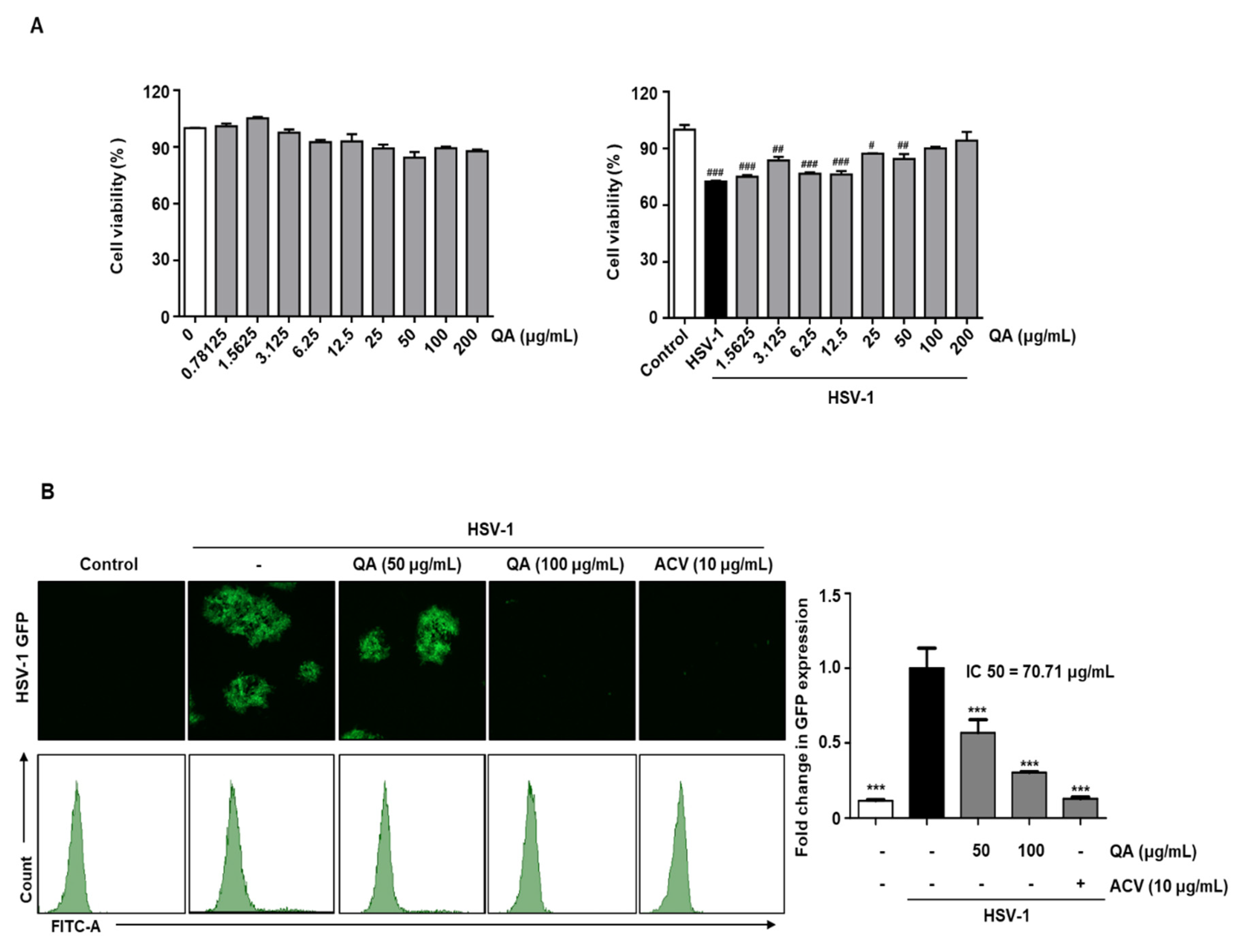
3.3. QA Exhibits Antiviral Effects upon Infection with HSV-1 in SK-N-SH Neuroblastoma Cells
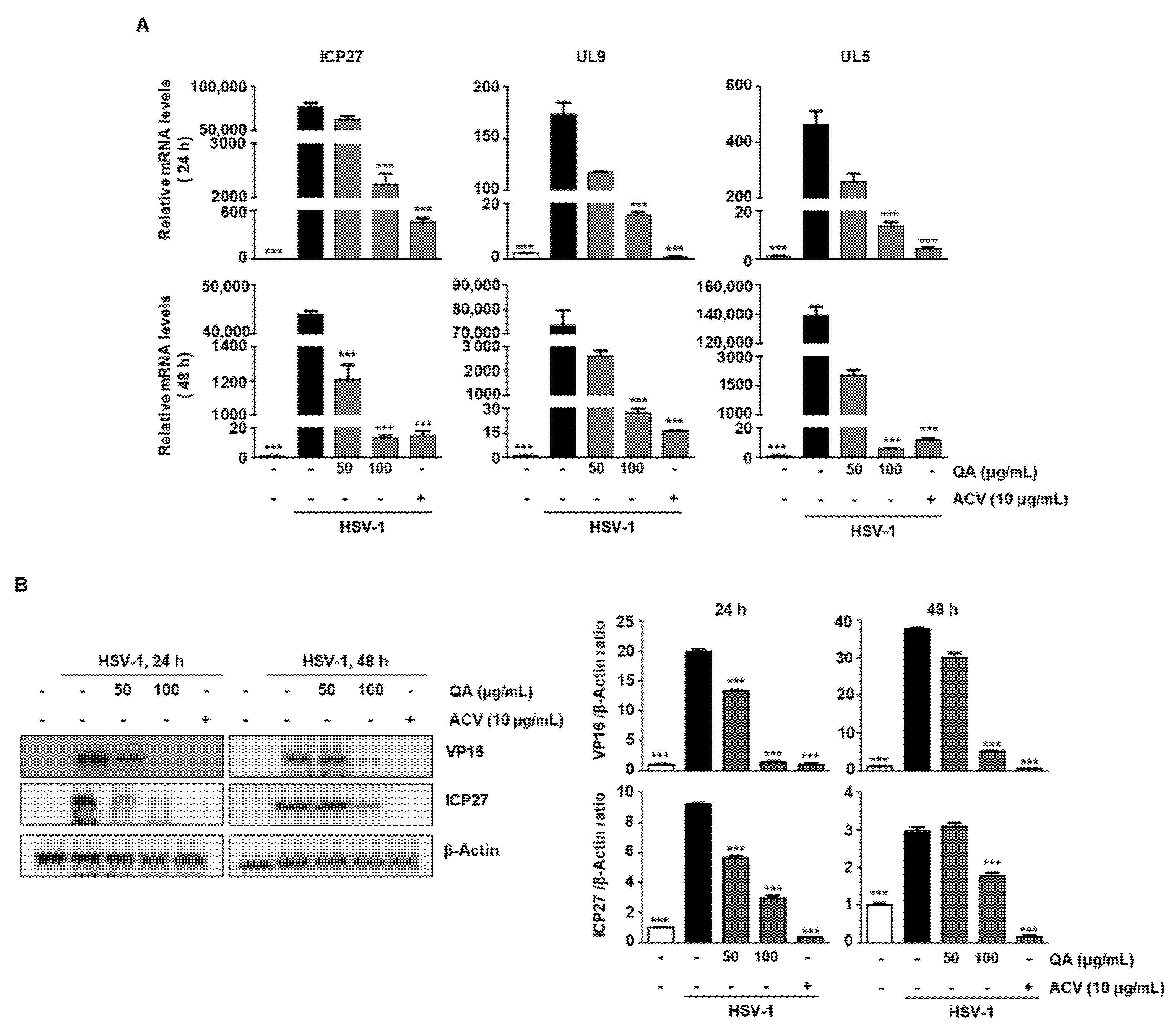
3.4. QA Exerts an Antiviral Effect by Inhibiting ICP27, an IE Gene, during HSV-1 Infection in SK-N-SH Cells
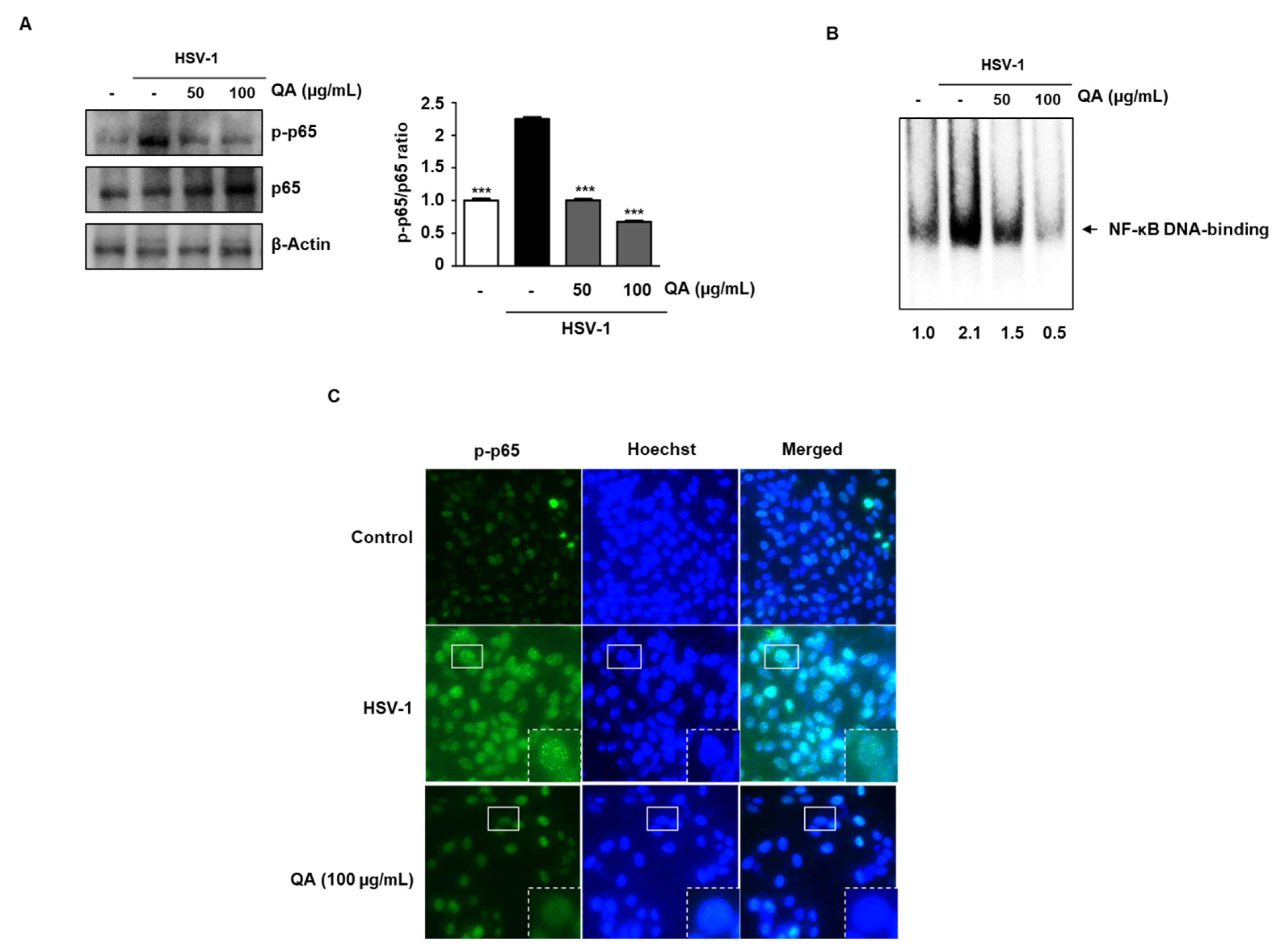
3.5. QA Inhibits NF-κB Phosphorylation Activity by HSV-1 Infection in SK-N-SH Cells
3.6. QA Induces Inhibition of ROS Production by HSV-1 Infection in SK-N-SH Cells
3.7. QA Suppresses the Expression of Inflammatory Cytokines by HSV-1 Infection
3.8. The HSV-1 Antiviral Effect by QA Treatment Indicates a Role for TBK1/IRF3 Activation
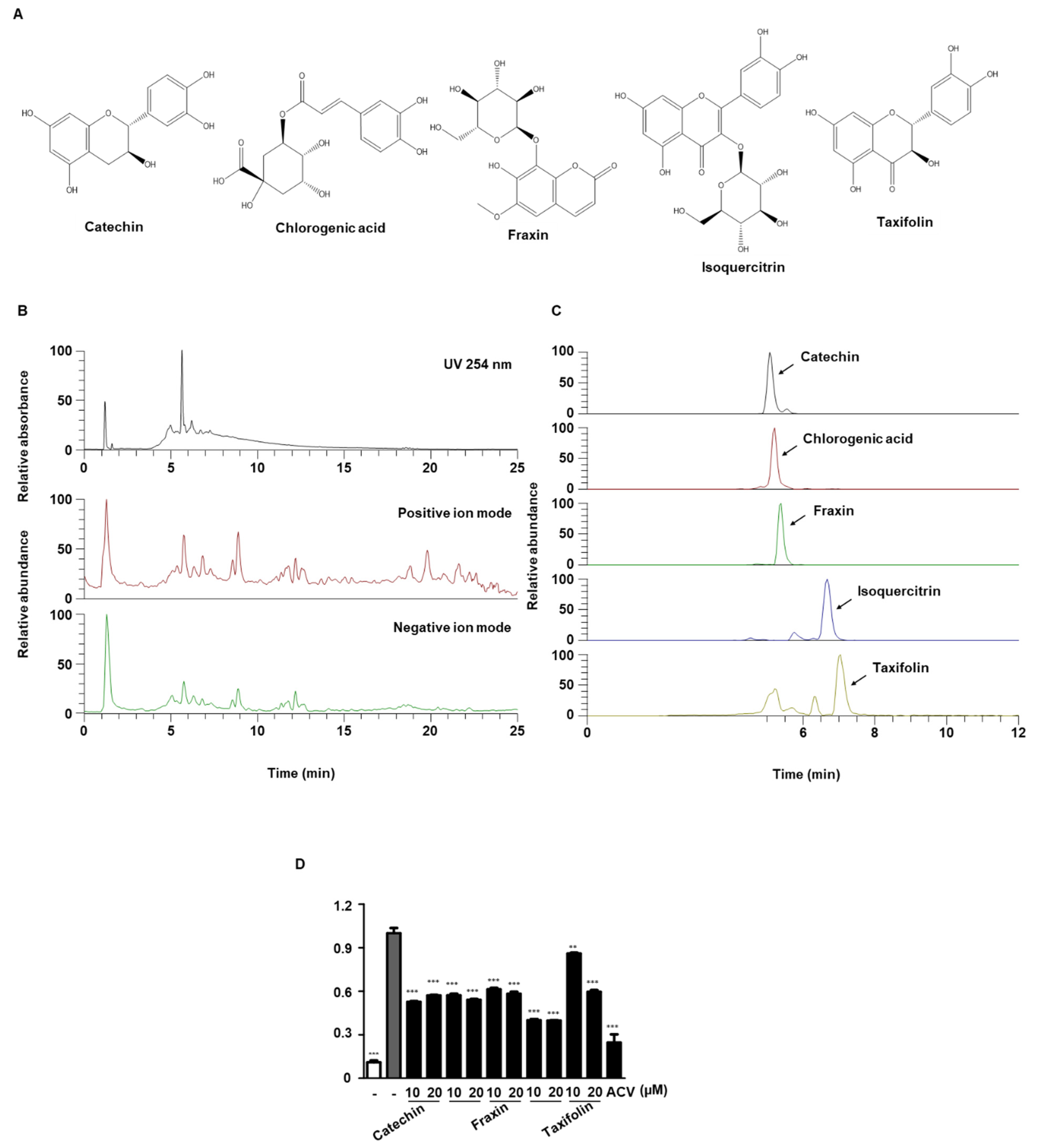
3.9. UPLC-MS/MS Analysis of QA
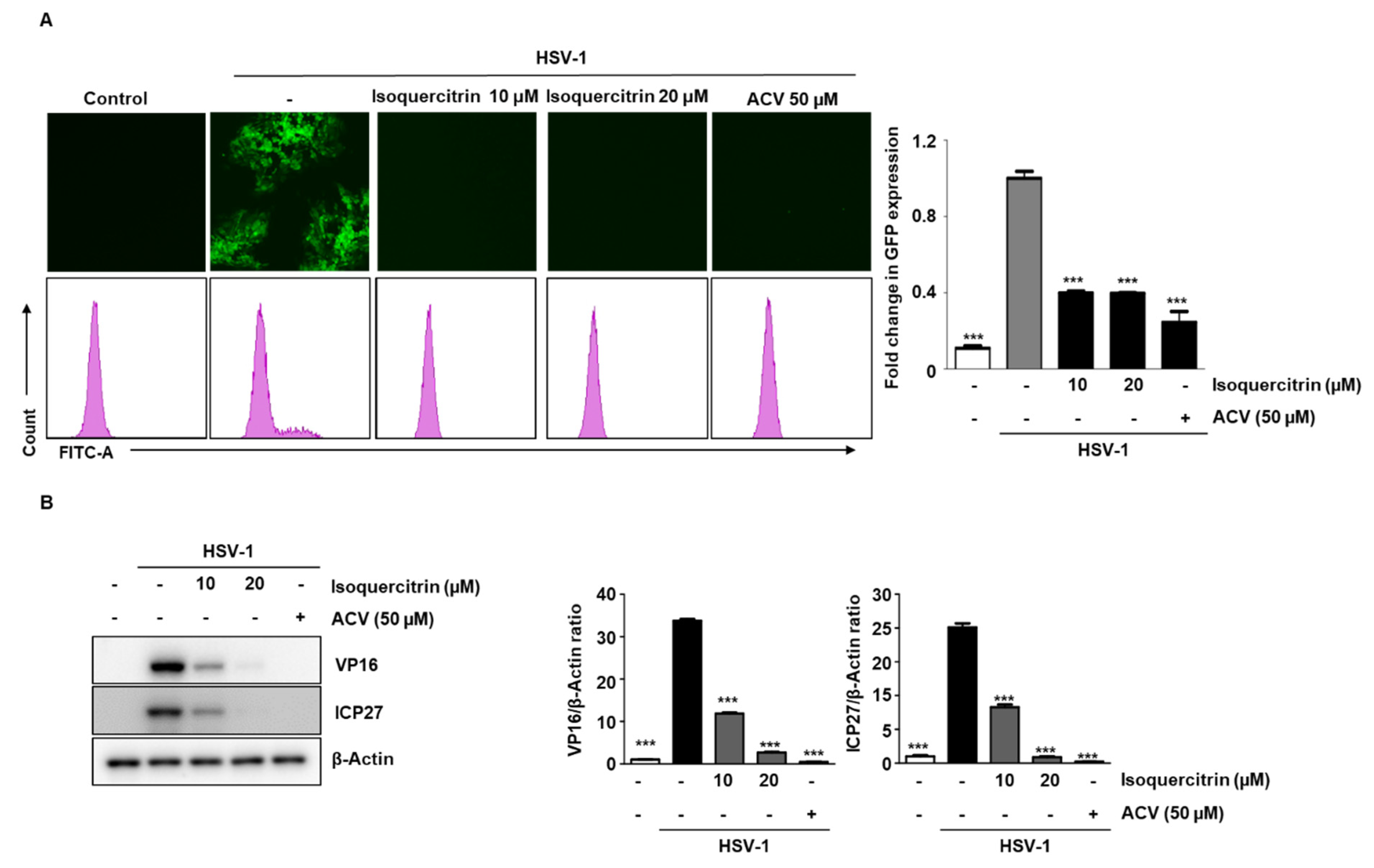
3.10. Isoquercitrin Isolated from QA Inhibits Viral Activity through ICP27 Inhibition during HSV-1 Infection
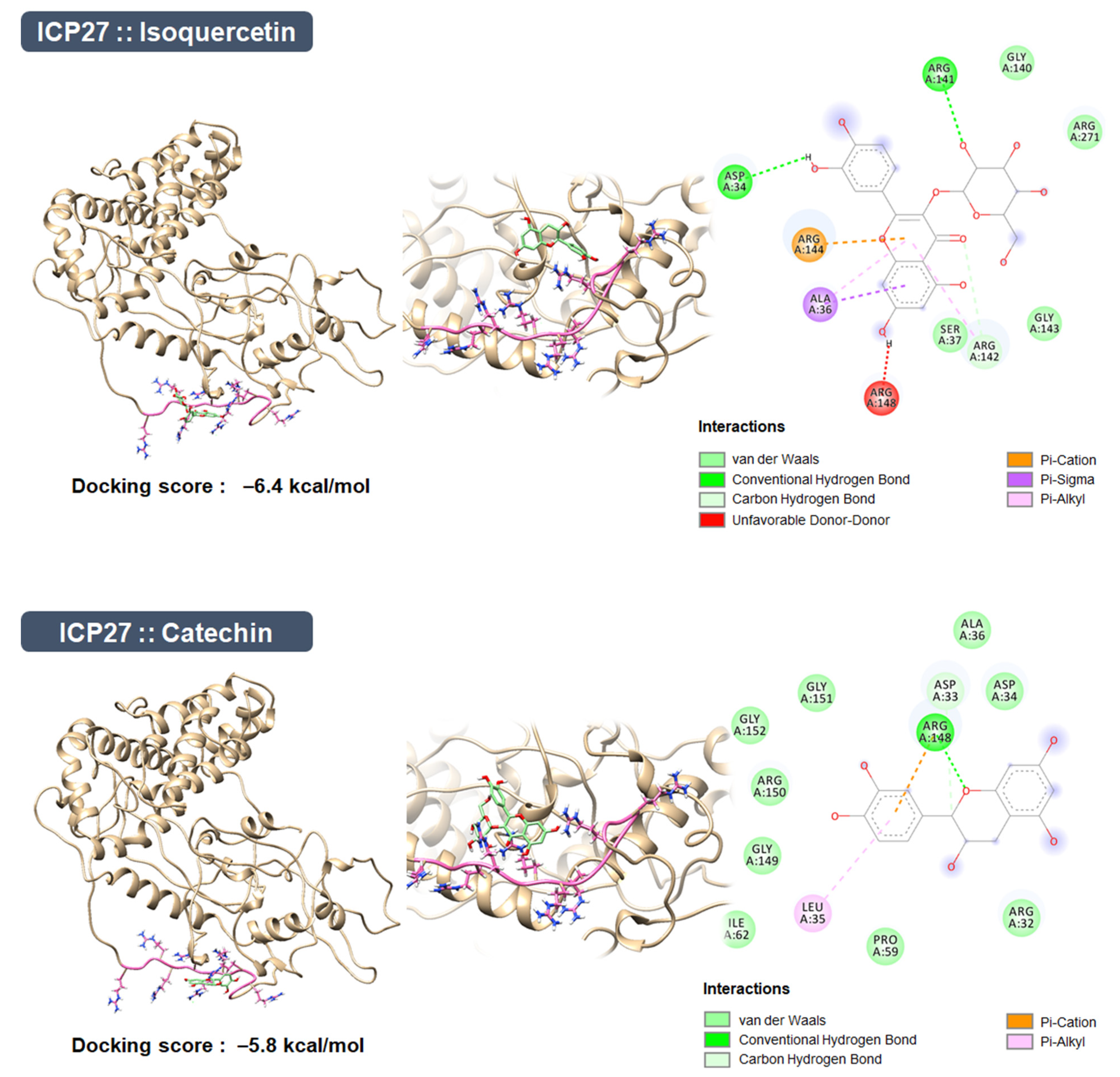
3.11. Construction of the ICP27 Structure and Molecular Docking Simulation
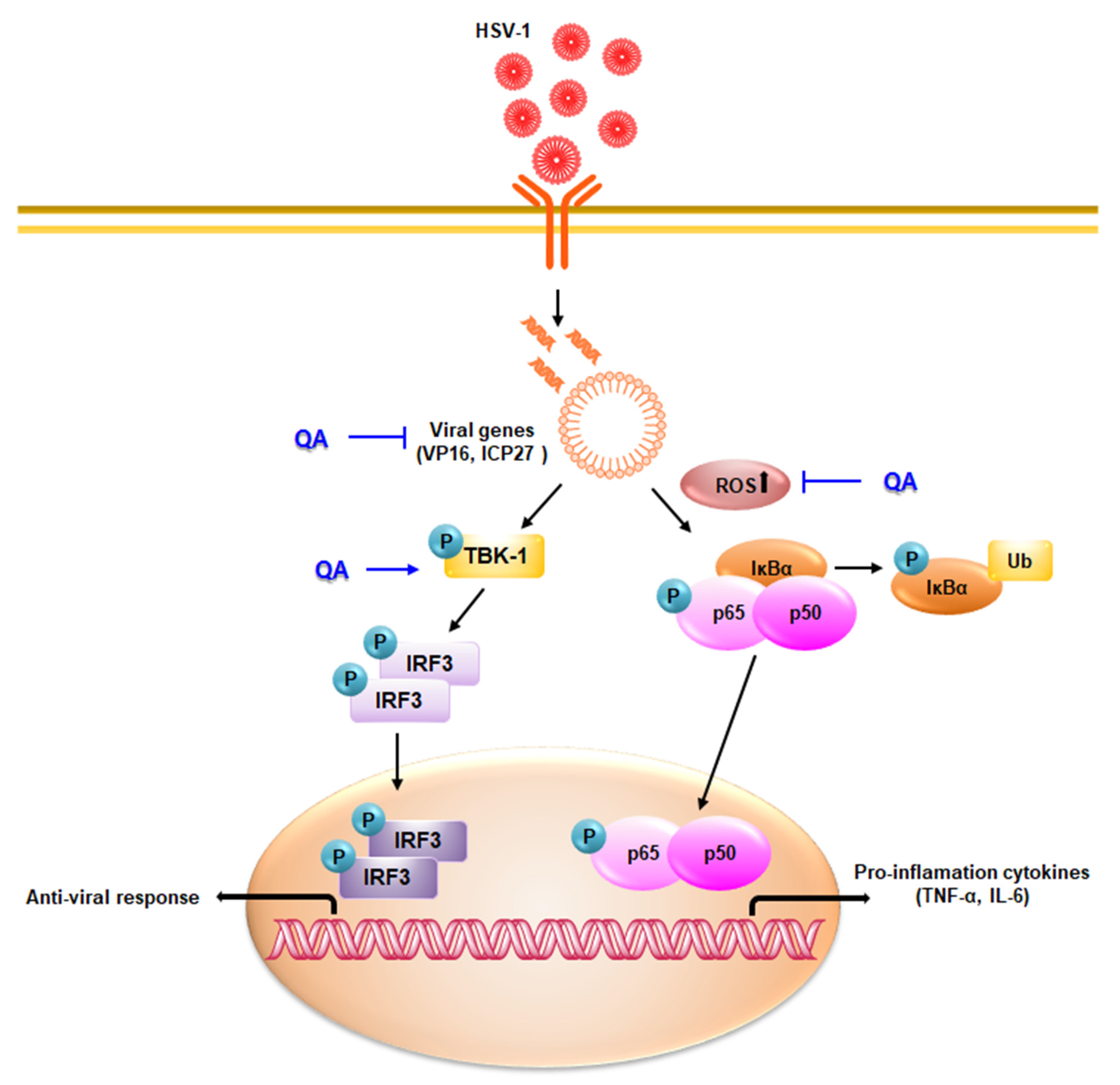
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Midak-Siewirska, A.; Karabin, K.; Chudzik, E.; Dzieciatkowski, T.; Przybylski, M.; Majewska, A.; Luczak, M.; Mlynarczyk, G. Application of real-time PCR assay for investigating the presence of herpes simplex virus type 1 DNA. Med. Dosw. Mikrobiol. 2010, 62, 85–92. [Google Scholar] [PubMed]
- Whitley, R.J.; Roizman, B. Herpes simplex virus infections. Lancet 2001, 357, 1513–1518. [Google Scholar] [CrossRef]
- Rozenberg, F.; Deback, C.; Agut, H. Herpes simplex encephalitis: From virus to therapy. Infect. Disord. Drug Targets 2011, 11, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Menendez, C.M.; Carr, D.J.J. Defining nervous system susceptibility during acute and latent herpes simplex virus-1 infection. J. Neuroimmunol. 2017, 308, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.P. Regulation of herpes simplex virus gene expression. Gene 2001, 271, 117–130. [Google Scholar] [CrossRef]
- Johnson, K.E.; Song, B.; Knipe, D.M. Role for herpes simplex virus 1 ICP27 in the inhibition of type I interferon signaling. Virology 2008, 374, 487–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, I.H.; Li, L.; Silva, L.; Sandri-Goldin, R.M. ICP27 recruits Aly/REF but not TAP/NXF1 to herpes simplex virus type 1 transcription sites although TAP/NXF1 is required for ICP27 export. J. Virol. 2005, 79, 3949–3961. [Google Scholar] [CrossRef] [Green Version]
- Paludan, S.R.; Bowie, A.G.; Horan, K.A.; Fitzgerald, K.A. Recognition of herpesviruses by the innate immune system. Nat. Rev. Immunol. 2011, 11, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Ni, L.; Wang, S.; Wang, K.; Lin, R.; Zheng, C. Herpes simplex virus 1-encoded tegument protein VP16 abrogates the production of beta interferon (IFN) by inhibiting NF-kappaB activation and blocking IFN regulatory factor 3 to recruit its coactivator CBP. J. Virol. 2013, 87, 9788–9801. [Google Scholar] [CrossRef] [Green Version]
- Christensen, M.H.; Jensen, S.B.; Miettinen, J.J.; Luecke, S.; Prabakaran, T.; Reinert, L.S.; Mettenleiter, T.; Chen, Z.J.; Knipe, D.M.; Sandri-Goldin, R.M.; et al. HSV-1 ICP27 targets the TBK1-activated STING signalsome to inhibit virus-induced type I IFN expression. EMBO J. 2016, 35, 1385–1399. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef]
- Ishikawa, H.; Barber, G.N. The STING pathway and regulation of innate immune signaling in response to DNA pathogens. Cell Mol. Life Sci. 2011, 68, 1157–1165. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Zhan, G.; Zheng, C. Evasion of host antiviral innate immunity by HSV-1, an update. Virol. J. 2016, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.C.; Feng, H.; Lin, Y.C.; Guo, X.R. New strategies against drug resistance to herpes simplex virus. Int. J. Oral Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef]
- De Clercq, E. Antivirals: Past, present and future. Biochem. Pharm. 2013, 85, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Boivin, G. Resistance of herpes simplex viruses to nucleoside analogues: Mechanisms, prevalence, and management. Antimicrob. Agents Chemother. 2011, 55, 459–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, I.S.; Park, D.H.; Bae, M.S.; Oh, D.S.; Kwon, N.H.; Kim, J.E.; Choi, C.Y.; Cho, S.S. In Vitro and In Vivo Studies on Quercus acuta Thunb. (Fagaceae) Extract: Active Constituents, Serum Uric Acid Suppression, and Xanthine Oxidase Inhibitory Activity. Evid. Based Complement. Altern. Med. 2017, 2017, 4097195. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Park, D.H.; Bae, M.S.; Song, S.H.; Seo, H.J.; Han, D.G.; Oh, D.S.; Jung, S.T.; Cho, Y.C.; Park, K.M.; et al. Analysis of the Active Constituents and Evaluation of the Biological Effects of Quercus acuta Thunb. (Fagaceae) Extracts. Molecules 2018, 23, 1772. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.H.; Park, K.H.; Kim, M.H.; Kim, H.H.; Kim, S.R.; Park, K.J.; Heo, J.H.; Lee, M.W. Anti-Oxidative and Anti-Inflammatory Effects of Phenolic Compounds from the Stems of Quercus acuta Thunberg. Asian J. Chem. 2014, 26, 4582–4586. [Google Scholar] [CrossRef]
- Serit, M.; Okubo, T.; Su, R.-H.; Hagiwara, N.; Kim, M.; Iwagawa, T.; Yamamoto, T. Antibacterial Compounds from Oak, Quercus acutaThunb. Agric. Biol. Chem. 2014, 55, 19–23. [Google Scholar] [CrossRef]
- Kim, T.I.; Kwon, E.B.; Oh, Y.C.; Go, Y.; Choi, J.G. Mori ramulus and its Major Component Morusin Inhibit Herpes Simplex Virus Type 1 Replication and the Virus-Induced Reactive Oxygen Species. Am. J. Chin. Med. 2021, 49, 163–179. [Google Scholar] [CrossRef]
- Hwang, Y.H.; Ma, J.Y. Preventive Effects of an UPLC-DAD-MS/MS Fingerprinted Hydroalcoholic Extract of Citrus aurantium in a Mouse Model of Ulcerative Colitis. Planta Med. 2018, 84, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhao, L.; Cai, W.; Shao, J.; Huang, X.; Liu, Y. An accurate and reproducible method for simultaneous determination of four flavonoids in EtOAc extracts from Sophora flavescens Ait. in rat plasma based on UHPLC Q-Exactive Mass spectrometry: Application to a pharmacokinetics study. Biomed. Chromatogr. 2019, 33, e4447. [Google Scholar] [CrossRef] [PubMed]
- Mador, N.; Goldenberg, D.; Cohen, O.; Panet, A.; Steiner, I. Herpes simplex virus type 1 latency-associated transcripts suppress viral replication and reduce immediate-early gene mRNA levels in a neuronal cell line. J. Virol. 1998, 72, 5067–5075. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, A.M.; McMahan, L.; Schaffer, P.A. Herpes simplex virus type 1 ICP27 deletion mutants exhibit altered patterns of transcription and are DNA deficient. J. Virol. 1989, 63, 18–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Guan, Y.; Sun, X.; Shi, L.; Liang, R.; Lv, X.; Xin, W. HSV-1 activates NF-kappaB in mouse astrocytes and increases TNF-alpha and IL-6 expression via Toll-like receptor 3. Neurol. Res. 2013, 35, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Marino-Merlo, F.; Papaianni, E.; Frezza, C.; Pedatella, S.; De Nisco, M.; Macchi, B.; Grelli, S.; Mastino, A. NF-kappaB-Dependent Production of ROS and Restriction of HSV-1 Infection in U937 Monocytic Cells. Viruses 2019, 11, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, A.K.; Chow, E.K.; Goodnough, J.B.; Yeh, W.C.; Cheng, G. Differential requirement for TANK-binding kinase-1 in type I interferon responses to toll-like receptor activation and viral infection. J. Exp. Med. 2004, 199, 1651–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Yi, Y.S.; Yang, Y.; Oh, J.; Jeong, D.; Cho, J.Y. The pivotal role of TBK1 in inflammatory responses mediated by macrophages. Mediat. Inflamm. 2012, 2012, 979105. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W. Negative regulation of TBK1-mediated antiviral immunity. FEBS Lett. 2013, 587, 542–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Han, F.; Song, A.; Wang, M.; Zhao, M.; Zhao, C. Rapid characterization of the chemical constituents of Cortex Fraxini by homogenate extraction followed by UHPLC coupled with Fourier transform ion cyclotron resonance mass spectrometry and GC-MS. J. Sep. Sci. 2016, 39, 4325–4334. [Google Scholar] [CrossRef]
- Spinola, V.; Pinto, J.; Llorent-Martinez, E.J.; Tomas, H.; Castilho, P.C. Evaluation of Rubus grandifolius L. (wild blackberries) activities targeting management of type-2 diabetes and obesity using in vitro models. Food Chem. Toxicol. 2019, 123, 443–452. [Google Scholar] [CrossRef]
- da Silva, N.A.; Rodrigues, E.; Mercadante, A.Z.; de Rosso, V.V. Phenolic compounds and carotenoids from four fruits native from the Brazilian Atlantic Forest. J. Agric. Food Chem. 2014, 62, 5072–5084. [Google Scholar] [CrossRef] [PubMed]
- Mears, W.E.; Rice, S.A. The RGG box motif of the herpes simplex virus ICP27 protein mediates an RNA-binding activity and determines in vivo methylation. J. Virol. 1996, 70, 7445–7453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri-Goldin, R.M. The many roles of the highly interactive HSV protein ICP27, a key regulator of infection. Future Microbiol. 2011, 6, 1261–1277. [Google Scholar] [CrossRef]
- Corbin-Lickfett, K.A.; Souki, S.K.; Cocco, M.J.; Sandri-Goldin, R.M. Three arginine residues within the RGG box are crucial for ICP27 binding to herpes simplex virus 1 GC-rich sequences and for efficient viral RNA export. J. Virol. 2010, 84, 6367–6376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, I.; Goshima, F.; Ito, H.; Koide, N.; Yoshida, T.; Yokochi, T.; Kimura, Y.; Nishiyama, Y. The vomeronasal chemosensory system as a route of neuroinvasion by herpes simplex virus. Virology 2005, 334, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcocci, M.E.; Napoletani, G.; Protto, V.; Kolesova, O.; Piacentini, R.; Li Puma, D.D.; Lomonte, P.; Grassi, C.; Palamara, A.T.; De Chiara, G. Herpes Simplex Virus-1 in the Brain: The Dark Side of a Sneaky Infection. Trends Microbiol. 2020, 28, 808–820. [Google Scholar] [CrossRef]
- Ounanian, A.; Guilbert, B.; Renversez, J.C.; Seigneurin, J.M.; Avrameas, S. Antibodies to viral antigens, xenoantigens, and autoantigens in Alzheimer’s disease. J. Clin. Lab. Anal. 1990, 4, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Leyton, L.; Hott, M.; Acuna, F.; Caroca, J.; Nunez, M.; Martin, C.; Zambrano, A.; Concha, M.I.; Otth, C. Nutraceutical activators of AMPK/Sirt1 axis inhibit viral production and protect neurons from neurodegenerative events triggered during HSV-1 infection. Virus Res. 2015, 205, 63–72. [Google Scholar] [CrossRef]
- Schachtele, S.J.; Hu, S.; Little, M.R.; Lokensgard, J.R. Herpes simplex virus induces neural oxidative damage via microglial cell Toll-like receptor-2. J. Neuroinflamm. 2010, 7, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Dosal, R.; Horan, K.A.; Rahbek, S.H.; Ichijo, H.; Chen, Z.J.; Mieyal, J.J.; Hartmann, R.; Paludan, S.R. HSV infection induces production of ROS, which potentiate signaling from pattern recognition receptors: Role for S-glutathionylation of TRAF3 and 6. PLoS Pathog. 2011, 7, e1002250. [Google Scholar] [CrossRef] [PubMed]
- Gregory, D.; Hargett, D.; Holmes, D.; Money, E.; Bachenheimer, S.L. Efficient replication by herpes simplex virus type 1 involves activation of the IkappaB kinase-IkappaB-p65 pathway. J. Virol. 2004, 78, 13582–13590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.Y.; Jouanguy, E.; Ugolini, S.; Smahi, A.; Elain, G.; Romero, P.; Segal, D.; Sancho-Shimizu, V.; Lorenzo, L.; Puel, A.; et al. TLR3 deficiency in patients with herpes simplex encephalitis. Science 2007, 317, 1522–1527. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | tR (min) | Molecular Formula | Precursor Ion (m/z) | Error (ppm) | MS/MS Fragments (m/z) | Identifications | ||
|---|---|---|---|---|---|---|---|---|
| Adduct | Expected | Measured | ||||||
| 1 | 5.07 | C15H14O6 | [M-H]− | 289.0718 | 289.0715 | −0.89 | 289, 245, 205, 125 | Catechin [18] a |
| 2 | 5.21 | C16H18O9 | [M+COOH-H]− | 399.0933 | 399.0930 | −0.81 | 191 | Chlorogenic Acid [32] a |
| 3 | 5.34 | C16H18O10 | [M-H]− | 369.0827 | 369.0822 | −1.45 | 369, 207, 192 | Fraxin [31] a |
| 4 | 6.67 | C21H20O12 | [M-H]− | 463.0882 | 463.0878 | −0.80 | 463, 300, 301, 151 | Isoquercitrin [33] a |
| 5 | 6.99 | C15H12O7 | [M-H]− | 303.0510 | 303.0508 | −0.92 | 285, 177, 125 | Taxifolin [18] a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.; Kim, Y.S.; Hwang, Y.-H.; Yang, H.J.; Li, W.; Kwon, E.-B.; Kim, T.I.; Go, Y.; Choi, J.-G. Quercus acuta Thunb. (Fagaceae) and Its Component, Isoquercitrin, Inhibit HSV-1 Replication by Suppressing Virus-Induced ROS Production and NF-κB Activation. Antioxidants 2021, 10, 1638. https://doi.org/10.3390/antiox10101638
Kim B, Kim YS, Hwang Y-H, Yang HJ, Li W, Kwon E-B, Kim TI, Go Y, Choi J-G. Quercus acuta Thunb. (Fagaceae) and Its Component, Isoquercitrin, Inhibit HSV-1 Replication by Suppressing Virus-Induced ROS Production and NF-κB Activation. Antioxidants. 2021; 10(10):1638. https://doi.org/10.3390/antiox10101638
Chicago/Turabian StyleKim, Buyun, Young Soo Kim, Youn-Hwan Hwang, Hye Jin Yang, Wei Li, Eun-Bin Kwon, Tae In Kim, Younghoon Go, and Jang-Gi Choi. 2021. "Quercus acuta Thunb. (Fagaceae) and Its Component, Isoquercitrin, Inhibit HSV-1 Replication by Suppressing Virus-Induced ROS Production and NF-κB Activation" Antioxidants 10, no. 10: 1638. https://doi.org/10.3390/antiox10101638