
An Overview on the Therapeutic Function of Foods Enriched with Plant Sterols in Diabetes Management

 ,
,
Abstract
:1. Introduction
2. Diabetes
2.1. Epidemics and Thrifty Genes
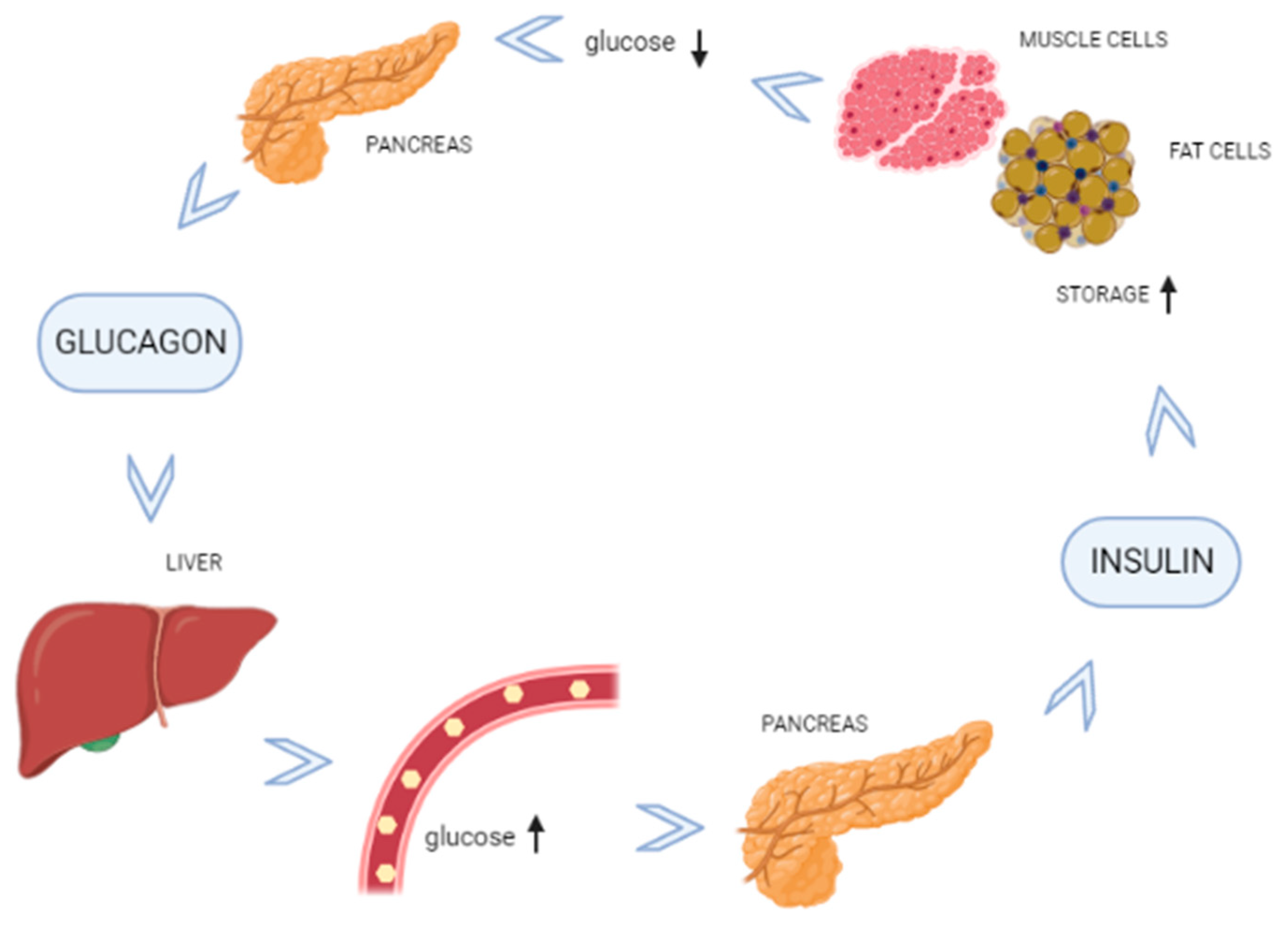
2.2. Blood Glucose Regulation
3. Phytosterols
- Polyphenols include flavonoids, phenolic acids, tannins, and stilbenes.
- Terpenoids include carotenoids and non-carotenoids.
- Thiols includes glucosinolates and indoles (Table 1).
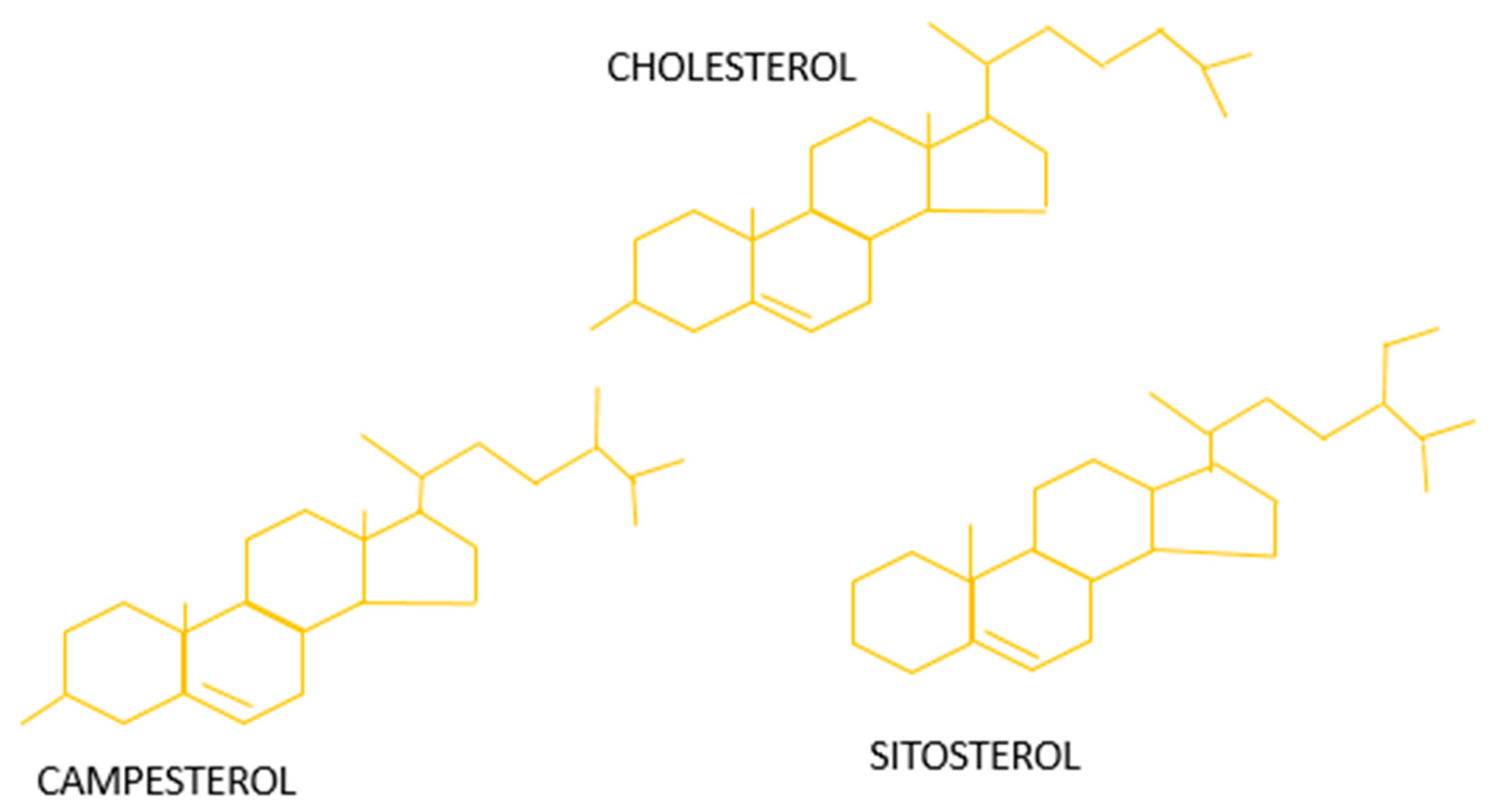
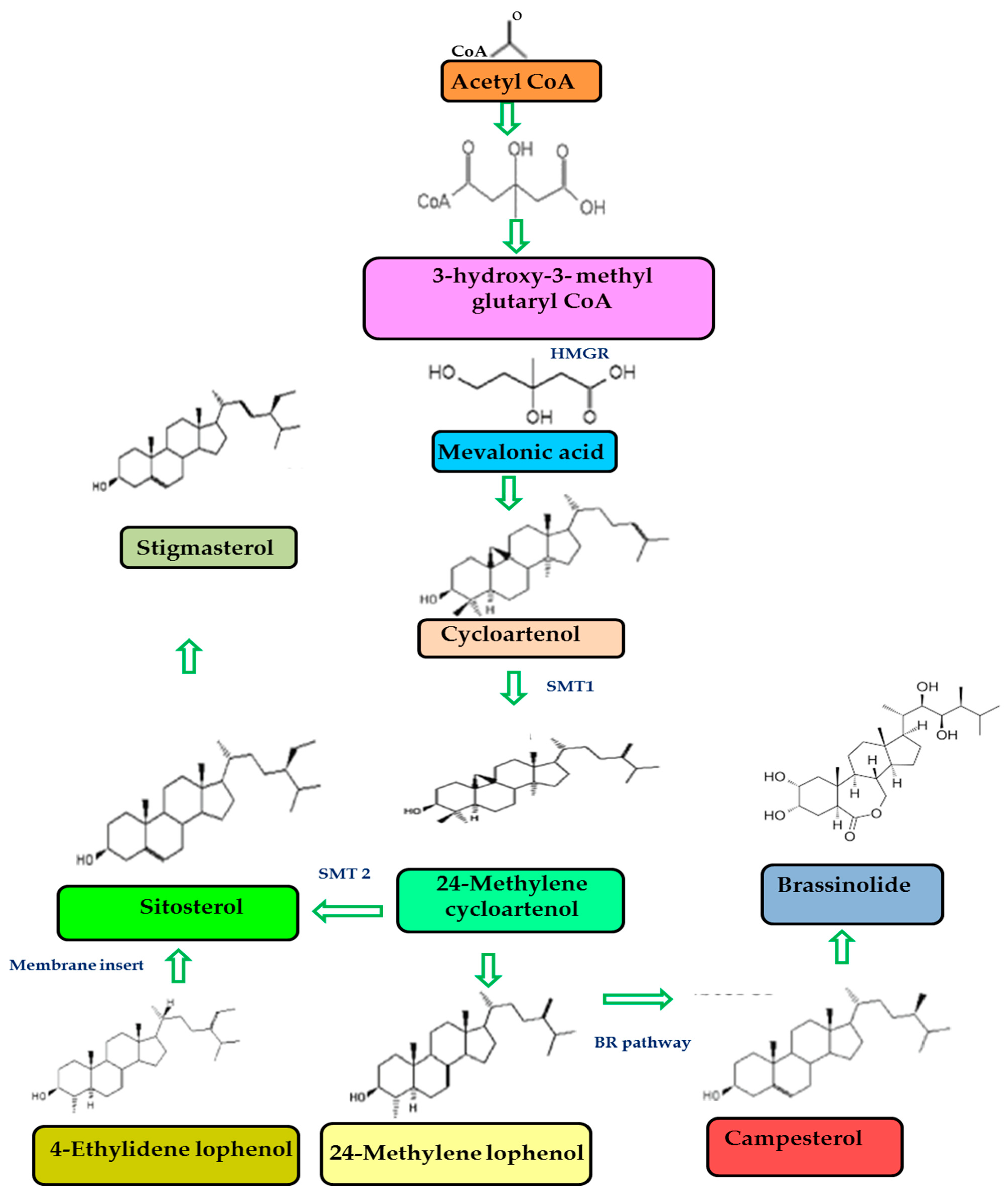
3.1. Phytosterols Chemical Structure and Bioavailability
3.2. Mechanism of Action
4. Diabetes and Cholesterol
4.1. Genetic and Epigenetic Modulations in Diabetes
4.2. Plant Sterols and Anti-Cholesterol Activity
4.3. Plant Sterols and Anti-Diabetic Effects

{kind=link}
{kind=link}
{kind=link}
| Botanical Name | Common Name | Components Used | Animal Studies | Effects | Reference |
|---|---|---|---|---|---|
| Aloe barbadensis | Aloe vera | Leaves | Diabetic rats | Significant reduction in the levels of the enzymes that facilitate carbohydrate metabolism | [81] |
| Cuminum cyminum | Cumin | seeds | Diabetic rats | Improves insulin secretion | [82] |
| Nigella sativum | Black cumin | Seeds | Rats | Improves glucose tolerance | [83] |
| Chloroxylon switenia | Indian satinwood | Barks | Diabetic albino rats | Decreases blood glucose level | [84] |
| Forsythia suspense | Weeping forsythia | Fruits | STZ induced Kunming mice | Significant reduction in blood glucose level | [85] |
| Coccinia grandis | Scarlet gourds | Leaf | Diabetic Wistar rats | Improves insulin-secretagogue and cytoprotective activities | [86] |
| Afzelia africana | African mahogany | Stem | Diabetic Wistar rats | Reduces hyperglycemia | [93] |
| Urtica dioica | Common nettle | Leaf | Fructose induced Insulin resistance Wistar rats | Significantly reduces hyperglycemia and insulin resistance | [87] |
| Anacardium accidentale | Cashew tree | Leaf | Diabetes induced female albino Wistar rats | Significant reduction in the levels of serum glucose, glycosylated haemoglobin, FIRI, and serum insulin | [88] |
| Pleurotus ostreatus | Oyster mushroom | Diabetes induced male Wistar rats | Significant reduction in blood glucose level | [92] | |
| Uvaria chamae | Bush banana | Root | Diabetes induced albino rats | Significant improvement in the regeneration of islets of Langerhans | [94] |
| Cinnamomum zeylanicum | Cinnamon | Bark | STZ-induced rats | Significantly diminishes α-glucosidase activity | [73] |
| Ocimum basilicum | Basil | Leaves | Significantly inhibits α amylase activity in a dose-dependent manner | [97] | |
| Corchorus olitorius | Jute | Leaves | Significantly inhibits the enzymatic activities of α-amylase, α-glucosidase, and ACE | [99] | |
| Ficus deltoidea | Fig | Leaves, Flowers | STZ-induced diabetic rats | Significantly lowers the blood glucose level | [101] |
| Holarrhena antidysenterica | Bitter oleander | Seeds | Starch-loaded normoglycemic rats | Interferes with starch digestion | [75] |
| Olea europaea | Olive | Leaves | STZ-induced diabetic rats | Inhibits α amylase activity | [72,74] |
| Glycine max | Soybean | Soybean | Significantly lowers the levels of α-amylase, α- glucosidase and ACE | [100] |
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- American Diabetes Association: Diagnosis and classification of diabetes mellitus. Diabetes Care 2009, 32, S62–S67. [CrossRef] [Green Version]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Purnell, J.Q. Definitions, Classification, and Epidemiology of Obesity. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., Kaltsas, G., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2018. [Google Scholar]
- Ahirwar, R.; Mondal, P.R. Prevalence of obesity in India: A systematic review. Diabetes Metab. Syndr. 2019, 13, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, D.A.; Puglisi, M.J.; Hasty, A.H. Impact of increased adipose tissue mass on inflammation: Insulin resistance, and dyslipidemia. Curr. Diabetes Rep. 2009, 9, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitner, D.R.; Frühbeck, G.; Yumuk, V.; Schindler, K.; Micic, D.; Woodward, E.; Toplak, H. Obesity and Type 2 Diabetes: Two Diseases with a Need for Combined Treatment Strategies—EASO Can Lead the Way. Obes. Facts 2017, 10, 483–492. [Google Scholar] [CrossRef]
- Cabral, C.E.; Klein, M. Phytosterols in the Treatment of Hypercholesterolemia and Prevention of Cardiovascular Diseases. Arq. Bras. Cardiol. 2017, 109, 475–482. [Google Scholar] [CrossRef]
- Martianto, D.; Bararah, A.; Andarwulan, N.; Średnicka-Tober, D. Cross-Sectional Study of Plant Sterols Intake as a Basis for Designing Appropriate Plant Sterol-Enriched Food in Indonesia. Nutrients 2021, 13, 452. [Google Scholar] [CrossRef]
- Tipton, C.M. Susruta of India: An unrecognized contributor to the history of exercise physiology. J. Appl. Physiol. 2008, 104, 1553–1556. [Google Scholar] [CrossRef] [Green Version]
- Dean, L.; McEntyre, J. The Genetic Landscape of Diabetes [Internet]; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2004. [Google Scholar]
- Wolfsdorf, J.I.; Sperling, M.A. Diabetes Mellitus. In Textbook of Clinical Pediatrics; Elzouki, A.Y., Harfi, H.A., Nazer, H.M., Stapleton, F.B., Oh, W., Whitley, R.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Speakman, J.R. Thrifty genes for obesity: An attractive but flawed idea, and an alternative perspective: The ‘drifty gene’ hypothesis. Int. J. Obes. 2008, 32, 1611–1617. [Google Scholar]
- Neel, J.V. Diabetes mellitus: A “thrifty” genotype rendered detrimental by progress. Am. J. Hum. Genet. 1962, 14, 353–362. [Google Scholar] [PubMed]
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes. 2015, 6, 850–867. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, M.; Ignacimuthu, S. Hypocholesterolemic effect of Indian medicinal plants—A review. Med. Chem. 2015, 5, 40–49. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Anti-oxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Yang, R.; Xue, L.; Zhang, L.; Wang, X.; Qi, X.; Jiang, J.; Yu, L.; Wang, X.; Zhang, W.; Zhang, Q.; et al. Phytosterol Contents of Edible Oils and Their Contributions to Estimated Phytosterol Intake in the Chinese Diet. Foods 2019, 8, 334. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, L.; Hu, G.Q.; Zhang, N.; Zhu, X.D.; Yang, K.Y.; Jin, F.; Shi, M.; Chen, Y.P.; Hu, W.H.; et al. Gemcitabine and Cisplatin Induction Chemotherapy in Nasopharyngeal Carcinoma. N. Engl. J. Med. 2019, 381, 1124–1135. [Google Scholar] [CrossRef]
- Ostlund, R.E., Jr. Phytosterols in human nutrition. Annu. Rev. Nutr. 2002, 22, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Martins, C.M.; Fonseca, F.A.; Ballus, C.A.; Figueiredo-Neto, A.M.; Meinhart, A.D.; de Godoy, H.T.; Izar, M.C. Common sources and composition of phytosterols and their estimated intake by the population in the city of São Paulo, Brazil. Nutrition 2013, 29, 865–871. [Google Scholar] [CrossRef]
- Nowicka, B.; Kruk, J. Occurrence biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta 2010, 1797, 1587–1605. [Google Scholar] [CrossRef] [Green Version]
- Griebel, T.; Zeier, J. A role for beta-sitosterol to stigmasterol conversion in plant-pathogen interactions. Plant J. 2010, 63, 254–268. [Google Scholar] [CrossRef]
- Sardinia, S.; Manali, A.; Gohari, A.R.; Abdollahi, M. The story of beta-sitosterol-a review. Eur. J. Med. Plants 2014, 4, 590–609. [Google Scholar]
- Dufourc, E.J. Sterols and membrane dynamics. J. Chem. Biol. 2008, 1, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, A.; Altabella, T.; Arró, M.; Boronat, A. Emerging roles for conjugated sterols in plants. Prog. Lipid Res. 2017, 67, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Knol, D.; Cardinault, N.; Nowicki, M.; Bott, R.; Antona, C.; Borel, P.; Bernard, J.P.; Duchateau, G.; Lairon, D. Phytosterol ester processing in the small intestine: Impact on cholesterol availability for absorption and chylomicron cholesterol incorporation in healthy humans. J. Lipid Res. 2011, 52, 1256–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Quispe, C.; Sharifi-Rad, J.; Cruz-Martins, N.; Nigam, M.; Mishra, A.P.; Konovalov, D.A.; Orobinskaya, V.; Abu-Reidah, I.M.; Zam, W.; et al. Phytosterols: From Preclinical Evidence to Potential Clinical Applications. Front. Pharmacol. 2021, 11, 599959. [Google Scholar] [CrossRef]
- Vezza, T.; Canet, F.; de Marañón, A.M.; Bañuls, C.; Rocha, M.; Víctor, V.M. Phytosterols: Nutritional Health Players in the Management of Obesity and Its Related Disorders. Antioxidants 2020, 9, 1266. [Google Scholar] [CrossRef]
- Jones, P.; Shamloo, M.; MacKay, D.S.; Rideout, T.C.; Myrie, S.B.; Plat, J.; Roullet, J.B.; Baer, D.J.; Calkins, K.L.; Davis, H.R.; et al. Progress and perspectives in plant sterol and plant stanol research. Nutr. Rev. 2018, 76, 725–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.; Ellegård, L.; Jessup, W.; Jones, P.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2013, 232, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed.; Wiley: Chichester, UK; New York, NY, USA, 2002. [Google Scholar]
- Kongduang, D.; Wungsintaweekul, J.; de-Eknamkul, W. Biosynthesis of β-sitosterol and stigmasterol proceeds exclusively via the mevalonate pathway in cell suspension cultures of Croton stellatopilosus. Tetrahedron Lett. 2008, 49, 4067–4072. [Google Scholar] [CrossRef]
- Miras-Moreno, B.; Sabater-Jara, A.B.; Pedreño, M.A.; Almagro, L. Bioactivity of Phytosterols and Their Production in Plant in Vitro Cultures. J. Agric. Food Chem. 2016, 64, 7049–7058. [Google Scholar] [CrossRef]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef]
- Jesch, E.D.; Carr, T.P. Food Ingredients That Inhibit Cholesterol Absorption. Prev. Nutr. Food Sci. 2017, 22, 67–80. [Google Scholar]
- Gylling, H.; Simonen, P. Phytosterols, Phytostanols, and Lipoprotein Metabolism. Nutrients 2015, 7, 7965–7977. [Google Scholar] [CrossRef] [Green Version]
- Cohn, J.S.; Kamili, A.; Wat, E.; Chung, R.W.; Tandy, S. Reduction in intestinal cholesterol absorption by various food components: Mechanisms and implications. Atheroscler. Suppl. 2010, 11, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; Barreto Arantes, M.; Menezes de Faria Pereira, S.; Leandro da Cruz, L.; de Souza Passos, M.; Pereira de Moraes, L.; Vieira, I.; Barros de Oliveira, D. Plants as Sources of Anti-Inflammatory Agents. Molecules 2020, 25, 3726. [Google Scholar] [CrossRef]
- De Jong, A.; Plat, J.; Bast, A.; Godschalk, R.W.; Basu, S.; Mensink, R.P. Effects of plant sterol and stanol ester consumption on lipid metabolism, anti-oxidant status and markers of oxidative stress, endothelial function and low-grade inflammation in patients on current statin treatment. Eur. J. Clin. Nutr. 2008, 62, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Dumolt, J.H.; Rideout, T.C. The Lipid-lowering Effects and Associated Mechanisms of Dietary Phytosterol Supplementation. Curr. Pharm. Des. 2017, 23, 5077–5085. [Google Scholar] [CrossRef]
- Rideout, T.C.; Marinangeli, C.; Harding, S.V. Triglyceride-Lowering Response to Plant Sterol and Stanol Consumption. J. AOAC Int. 2015, 98, 707–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Smet, E.; Mensink, R.P.; Plat, J. Effects of plant sterols and stanols on intestinal cholesterol metabolism: Suggested mechanisms from past to present. Mol. Nutr. Food Res. 2012, 56, 1058–1072. [Google Scholar] [CrossRef]
- Chen, H.C. Molecular mechanisms of sterol absorption. J. Nutr. 2001, 131, 2603–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brufau, G.; Kuipers, F.; Lin, Y.; Trautwein, E.A.; Groen, A.K. A reappraisal of the mechanism by which plant sterols promote neutral sterol loss in mice. PLoS ONE 2011, 6, e21576. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.B. Recent advances in understanding the STSL locus and ABCG5/ABCG8 biology. Curr. Opin. Lipidol. 2014, 25, 169–175. [Google Scholar] [CrossRef]
- Solati, Z.; Moghadasian, M.H. Use of Animal Models in Plant Sterol and Stanol Research. J. AOAC Int. 2015, 98, 691–696. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5280794, Stigmasterol. 2021. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Stigmasterol (accessed on 23 November 2021).
- Tabish, S.A. Is Diabetes Becoming the Biggest Epidemic of the Twenty-first Century? Int. J. Health Sci. 2007, 1, V–VIII. [Google Scholar]
- Guariguata, L.; Whiting, D.R.; Hambleton, I.; Beagley, J.; Linnenkamp, U.; Shaw, J.E. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res. Clin. Pract. 2014, 103, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Chadt, A.; Scherneck, S.; Joost, H.G.; Al-Hasani, H. Molecular links between Obesity and Diabetes: “Diabesity”. [Updated 23 January 2018]. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK279051/ (accessed on 23 November 2021).
- Schofield, J.D.; Liu, Y.; Rao-Balakrishna, P.; Malik, R.A.; Soran, H. Diabetes Dyslipidemia. Diabetes Ther. Res. Treat. Educ. Diabetes Relat. Disord. 2016, 7, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Borén, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: Pathophysiological, genetic, and therapeutic insights: A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2020, 41, 2313–2330. [Google Scholar] [CrossRef]
- Kwak, S.H.; Park, K.S. Recent progress in genetic and epigenetic research on type 2 diabetes. Exp. Mol. Med. 2016, 48, e220. [Google Scholar] [CrossRef] [Green Version]
- Gu, H.F. Genetic and Epigenetic Studies in Diabetic Kidney Disease. Front. Genet. 2019, 10, 507. [Google Scholar] [CrossRef]
- Stankov, K.; Benc, D.; Draskovic, D. Genetic and epigenetic factors in etiology of diabetes mellitus type 1. Pediatrics 2013, 132, 1112–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rosa, S.; Arcidiacono, B.; Chiefari, E.; Brunetti, A.; Indolfi, C.; Foti, D.P. Type 2 Diabetes Mellitus and Cardiovascular Disease: Genetic and Epigenetic Links. Front. Endocrinol. 2018, 9, 2. [Google Scholar] [CrossRef]
- Sommese, L.; Benincasa, G.; Schiano, C.; Marfella, R.; Grimaldi, V.; Sorriento, A.; Lucchese, R.; Fiorito, C.; Sardu, C.; Nicoletti, G.F.; et al. Genetic and epigenetic-sensitive regulatory network in immune response: A putative link between HLA-G and diabetes. Expert Rev. Endocrinol. Metab. 2019, 14, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Schwenk, R.W.; Vogel, H.; Schürmann, A. Genetic and epigenetic control of metabolic health. Mol. Metab. 2013, 2, 337–347. [Google Scholar] [CrossRef]
- Othman, R.A.; Moghadasian, M.H. Beyond cholesterol-lowering effects of plant sterols: Clinical and experimental evidence of anti-inflammatory properties. Nutr. Rev. 2011, 69, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Jones, P.J.; Abumweis, S.S. Plant sterols: Factors affecting their efficacy and safety as functional food ingredients. Lipids Health Dis. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abumweis, S.S.; Barake, R.; Jones, P.J. Plant sterols/stanols as cholesterol lowering agents: A meta-analysis of randomized controlled trials. Food Nutr. Res. 2008, 52. [Google Scholar] [CrossRef]
- Demonty, I.; Ras, R.T.; Van der Knaap, H.C.; Duchateau, G.S.; Meijer, L.; Zock, P.L.; Geleijnse, J.M.; Trautwein, E.A. Continuous dose-response relationship of the LDL-cholesterol-lowering effect of phytosterol intake. J. Nutr. 2009, 139, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Musa-Veloso, K.; Paulionis, L.; Poon, T.; Lee, H.Y. The effects of almond consumption on fasting blood lipid levels: A systematic review and meta-analysis of randomised controlled trials. J. Nutr. Sci. 2016, 5, e34. [Google Scholar] [CrossRef] [Green Version]
- Genser, B.; Silbernagel, G.; De Backer, G.; Bruckert, E.; Carmena, R.; Chapman, M.J.; Deanfield, J.; Descamps, O.S.; Rietzschel, E.R.; Dias, K.C.; et al. Plant sterols and cardiovascular disease: A systematic review and meta-analysis. Eur. Heart J. 2012, 33, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Talati, R.; Sobieraj, D.M.; Makanji, S.S.; Phung, O.J.; Coleman, C.I. The comparative efficacy of plant sterols and stanols on serum lipids: A systematic review and meta-analysis. J. Am. Diet. Assoc. 2010, 110, 719–726. [Google Scholar] [CrossRef]
- Naumann, E.; Plat, J.; Mensink, R.P. Changes in serum concentrations of noncholesterol sterols and lipoproteins in healthy subjects do not depend on the ratio of plant sterols to stanols in the diet. J. Nutr. 2003, 133, 2741–2747. [Google Scholar] [CrossRef] [Green Version]
- Kothari, S.; Dhami-Shah, H.; Shah, S.R. Antidiabetic Drugs and Statins in Nonalcoholic Fatty Liver Disease. J. Clin. Exp. Hepatol. 2019, 9, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2017, 8, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Govindappa, M. A review on the role of plant(s) extracts and its phytochemicals for the management of diabetes. J. Diabetes Metab. 2015, 6, 1–38. [Google Scholar]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P. Indian herbs and herbal drugs used for the treatment of diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, N. A Review of Recent Investigations on Medicinal Herbs Possessing Anti-Diabetic Properties. J. Nutr. Disord. Ther. 2011, 1, 2. [Google Scholar] [CrossRef]
- Mentreddy, S.R. Medicinal plant species with potential anti-diabetic properties. J. Sci. Food Agric. 2007, 87, 743–750. [Google Scholar] [CrossRef]
- Governa, P.; Baini, G.; Borgonetti, V.; Cettolin, G.; Giachetti, D.; Magnano, A.R.; Miraldi, E.; Biagi, M. Phytotherapy in the Management of Diabetes: A Review. Molecules 2018, 23, 105. [Google Scholar] [CrossRef] [Green Version]
- Gaikwad, B.S.; Krishna Mohan, G.; Sandhya Rani, M. Phytochemicals for diabetes management. Pharm. Crop. 2014, 5, 11–28. [Google Scholar] [CrossRef] [Green Version]
- Dembinska-Kiec, A.; Mykkänen, O.; Kiec-Wilk, B.; Mykkänen, H. Antioxidant phytochemicals against type 2 diabetes. Br. J. Nutr. 2008, 99, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Gweon, O.C.; Seo, Y.J.; Im, J.; Kang, M.J.; Kim, M.J.; Kim, J.I. Antioxidant effect of garlic and aged black garlic in animal model of type 2 diabetes mellitus. Nutr. Res. Pract. 2009, 3, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, A.K.; Rao, J.M. Diabetes mellitus and multiple therapeutic approaches of phytochemicals: Present status and future prospects. Curr. Sci. 2002, 83, 30–38. [Google Scholar]
- Zhang, X.; Yang, S.; Chen, J.; Su, Z. Unraveling the Regulation of Hepatic Gluconeogenesis. Front. Endocrinol. 2019, 9, 802. [Google Scholar] [CrossRef]
- Misawa, E.; Tanaka, M.; Nomaguchi, K.; Yamada, M.; Toida, T.; Takase, M.; Iwatsuki, K.; Kawada, T. Administration of phytosterols isolated from Aloe vera gel reduce visceral fat mass and improve hyperglycemia in Zucker diabetic fatty (ZDF) rats. Obes. Res. Clin. Pract. 2008, 2, 239–245. [Google Scholar] [CrossRef]
- Patil, S.B.; Takalikar, S.S.; Joglekar, M.M.; Haldavnekar, V.S.; Arvindekar, A.U. Insulinotropic and β-cell protective action of cuminaldehyde, cuminol and an inhibitor isolated from Cuminumcyminum in streptozotocin-induced diabetic rats. Br. J. Nutr. 2013, 110, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Meddah, B.; Ducroc, R.; El Abbes Faouzi, M.; Eto, B.; Mahraoui, L.; Benhaddou-Andaloussi, A.; Martineau, L.C.; Cherrah, Y.; Haddad, P.S. Nigella sativa inhibits intestinal glucose absorption and improves glucose tolerance in rats. J. Ethnopharmacol. 2009, 121, 419–424. [Google Scholar] [CrossRef]
- Jayaprasad, B.; Sharavanan, P.S.; Sivaraj, R. Antidiabetic effect of Chloroxylonswietenia bark extracts on streptozotocin-induced diabetic rats. Beni-Suef Univ. J. Basic Appl. Sci. 2016, 5, 61–69. [Google Scholar]
- Zhang, Y.; Feng, F.; Chen, T.; Li, Z.; Shen, Q.W. Antidiabetic and antihyperlipidemic activities of Forsythia suspensa (Thunb.) Vahl (fruit) in streptozotocin-induced diabetes mice. J. Ethnopharmacol. 2016, 192, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.I.; Chopda, M.Z.; Patil, R.H.; Vishwakarma, K.S.; Maheshwari, V.L. In vivo anti-diabetic and anti-oxidant activities of Coccinia grandis leaf extract against streptozotocin-induced diabetes in experimental rats. Asian Pac. J. Trop. Dis. 2016, 6, 298–304. [Google Scholar] [CrossRef]
- Ahangarpour, A.; Mohammadian, M.; Dianat, M. Antidiabetic effect of hydro-alcholic urtica dioica leaf extract in male rats with fructose-induced insulin resistance. Iran J. Med. Sci. 2012, 37, 181–186. [Google Scholar]
- Jaiswal, Y.S.; Tatke, P.A.; Gabhe, S.Y.; Vaidya, A.B. Antidiabetic activity of extracts of Anacardium occidentale Linn. leaves on n-streptozotocin diabetic rats. J. Tradit. Complement. Med. 2016, 7, 421–427. [Google Scholar] [CrossRef]
- Liu, C.T.; Hsu, T.W.; Chen, K.M.; Tan, Y.P.; Lii, C.K.; Sheen, L.Y. The Antidiabetic Effect of Garlic Oil is Associated with Ameliorated Oxidative Stress but Not Ameliorated Level of Pro-inflammatory Cytokines in Skeletal Muscle of Streptozotocin-induced Diabetic Rats. J. Tradit. Complement. Med. 2012, 2, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Antu, K.A.; Riya, M.P.; Nair, A.; Mishra, A.; Srivastava, A.K.; Raghu, K.G. Symplocos cochinchinensis enhances insulin sensitivity via the down regulation of lipogenesis and insulin resistance in high energy diet rat model. J. Ethnopharmacol. 2016, 193, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Cheng, D.; Zhang, Z. Antidiabetic activity of Helicteresangustifolia root. Pharm. Biol. 2016, 54, 938–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.Y.; Korivi, M.; Yang, H.T.; Huang, C.C.; Chaing, Y.Y.; Tsai, Y.C. Effect of Pleurotus tuber-regium polysaccharides supplementation on the progression of diabetes complications in obese-diabetic rats. Chin. J. Physiol. 2014, 57, 198–208. [Google Scholar] [CrossRef] [Green Version]
- Oyedemi, S.O.; Adewusi, E.A.; Aiyegoro, O.A.; Akinpelu, D.A. Antidiabetic and haematological effect of aqueous extract of stem bark of Afzelia africana (Smith) on streptozotocin-induced diabetic Wistar rats. Asian Pac. J. Trop. Biomed. 2011, 1, 353–358. [Google Scholar]
- Emordi, J.E.; Agbaje, E.O.; Oreagba, I.A.; Iribhogbe, O.I. Antidiabetic and hypolipidemic activities of hydroethanolic root extract of Uvariachamae in streptozotocin induced diabetic albino rats. BMC Complement. Altern. Med. 2016, 16, 468. [Google Scholar] [CrossRef] [Green Version]
- Palsamy, P.; Subramanian, S. Resveratrol, a natural phytoalexin, normalizes hyperglycemia in streptozotocin-nicotinamide induced experimental diabetic rats. Biomed. Pharmacother. 2008, 62, 598–605. [Google Scholar] [CrossRef]
- El-Beshbishy, H.; Bahashwan, S. Hypoglycemic effect of basil (Ocimum basilicum) aqueous extract is mediated through inhibition of α-glucosidase and α-amylase activities: An in vitro study. Toxicol. Ind. Health 2012, 28, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, A.R.; Haque, M. Preparation of Medicinal Plants: Basic Extraction and Fractionation Procedures for Experimental Purposes. J. Pharm. Bioallied Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademiluyi, A.O.; Akinyemi, A.J.; Henle, T.; Saliu, J.A.; Schwarzenbolz, U. Inhibitory effect of polyphenol-rich extracts of jute leaf (Corchorus olitorius) on key enzyme linked to type 2 diabetes (α-amylase and α-glucosidase) and hypertension (angiotensin I converting) in vitro. J. Funct. Foods 2012, 4, 450–458. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G.; Aragbaiye, F.P.; Oyeleye, S.I.; Ogunsuyi, O.B. Antioxidant properties and in vitro α-amylase and α-glucosidase inhibitory properties of phenolics constituents from different varieties of Corchorus spp. J. Taibah Univ. Med. Sci. 2015, 10, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Ademiluyi, A.O.; Oboh, G. Soybean phenolic-rich extracts inhibit key-enzymes linked to type 2 diabetes (α-amylase and α-glucosidase) and hypertension (angiotensin I converting enzyme) in vitro. Exp. Toxicol. Pathol. 2013, 65, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Choo, C.Y.; Sulong, N.Y.; Man, F.; Wong, T.W. Vitexin and isovitexin from the leaves of Ficusdeltoidea with in-vivo α-glucosidase inhibition. J. Ethnopharmacol. 2012, 142, 776–781. [Google Scholar] [CrossRef]
| Plant-Derived Foods | Scientific Names | Total PS (mg/100 g) | Reference |
|---|---|---|---|
| Zucchini | Cucurbita pepo L. | 0.63 | [18] |
| Eggplant | Solanum melongena L. | 0.5 | [18] |
| Broccoli | Brassica oleracea var. Italica | 4–50 | [18] |
| Carrot | Daucus carota L. | 16–30 | [18] |
| Cauliflower | Brassica oleracea var. Botrytis L. | 44 | [18] |
| Endive | Cichorium endıvia L. | 16–20 | [19] |
| Spinach | Tetragonia expansa | 16 | [19] |
| White cabbage | Brassica oleracea var. Capital | 27.4 | [19] |
| Tomato | Lycopersicum esculentum Mill. | 9–10 | [19] |
| Green bean | Phaseolus vulgaris | 10–12 | [19] |
| Brown rice | Oryza sativa | 18–20 | [19] |
| Polished rice | Oryza sativa | 9–10 | [19] |
| Green pea | Pisum sativum | 25 | [19] |
| Brown beans | Phaseolus vulgaris L. | 16 | [20] |
| Black beans | Phaseolus vulgaris L. | 15 | [20] |
| Chickpeas | Cicer arietinum | 120 | [20] |
| Lentil | Lens esculenta | 117 | [20] |
| Soybean | Glycine max L. Merr. | 32–35 | [20] |
| Black soybean | Glycine max L. Merr. | 17–18 | [21] |
| Linseed | Linum usitatissimum | 44–45 | [21] |
| Acai | Euterpe oleracea | 14 | [21] |
| Avocado | Persea americana | 75 | [21] |
| Pineapple | Ananas comosus L. Merril | 3–5 | [21] |
| Banana | Musa acuminata × Musa balbisiana | 12–16 | [21] |
| Coconut | Cocos nucifera | 14 | [22] |
| Guava | Psidium guajava L. | 3–5 | [22] |
| Orange | Citrus sinensis | 23–24 | [22] |
| Apple | Malus Domestica | 13–18 | [22] |
| Papaya | Carica papaya | 4–5 | [22] |
| Mango | Mangifera indica L. | 1–2 | [22] |
| Strawberry | Fragaria vesca L. | 10–15 | [22] |
| Canola (Rapeseed) | Brassica napus | 250–878 | [22] |
| Coconut | Cocos nucifera | 73–75 | [22] |
| Sunflower | Helianthus annuus | 400–500 | [22] |
| Corn | Zea mays | 686–1400 | [22] |
| Soybean | Glycine max | 203–328 | [22] |
| Olive | Olea europaea | 114–162 | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayaraman, S.; Roy, A.; Vengadassalapathy, S.; Sekar, R.; Veeraraghavan, V.P.; Rajagopal, P.; Rengasamy, G.; Mukherjee, R.; Sekar, D.; Manjunathan, R. An Overview on the Therapeutic Function of Foods Enriched with Plant Sterols in Diabetes Management. Antioxidants 2021, 10, 1903. https://doi.org/10.3390/antiox10121903
Jayaraman S, Roy A, Vengadassalapathy S, Sekar R, Veeraraghavan VP, Rajagopal P, Rengasamy G, Mukherjee R, Sekar D, Manjunathan R. An Overview on the Therapeutic Function of Foods Enriched with Plant Sterols in Diabetes Management. Antioxidants. 2021; 10(12):1903. https://doi.org/10.3390/antiox10121903
Chicago/Turabian StyleJayaraman, Selvaraj, Anitha Roy, Srinivasan Vengadassalapathy, Ramya Sekar, Vishnu Priya Veeraraghavan, Ponnulakshmi Rajagopal, Gayathri Rengasamy, Raktim Mukherjee, Durairaj Sekar, and Reji Manjunathan. 2021. "An Overview on the Therapeutic Function of Foods Enriched with Plant Sterols in Diabetes Management" Antioxidants 10, no. 12: 1903. https://doi.org/10.3390/antiox10121903