
Improving Cognition with Nutraceuticals Targeting TGF-β1 Signaling


 , ,
, ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Nutraceuticals and Mental Health: Focus on Cognitive Function
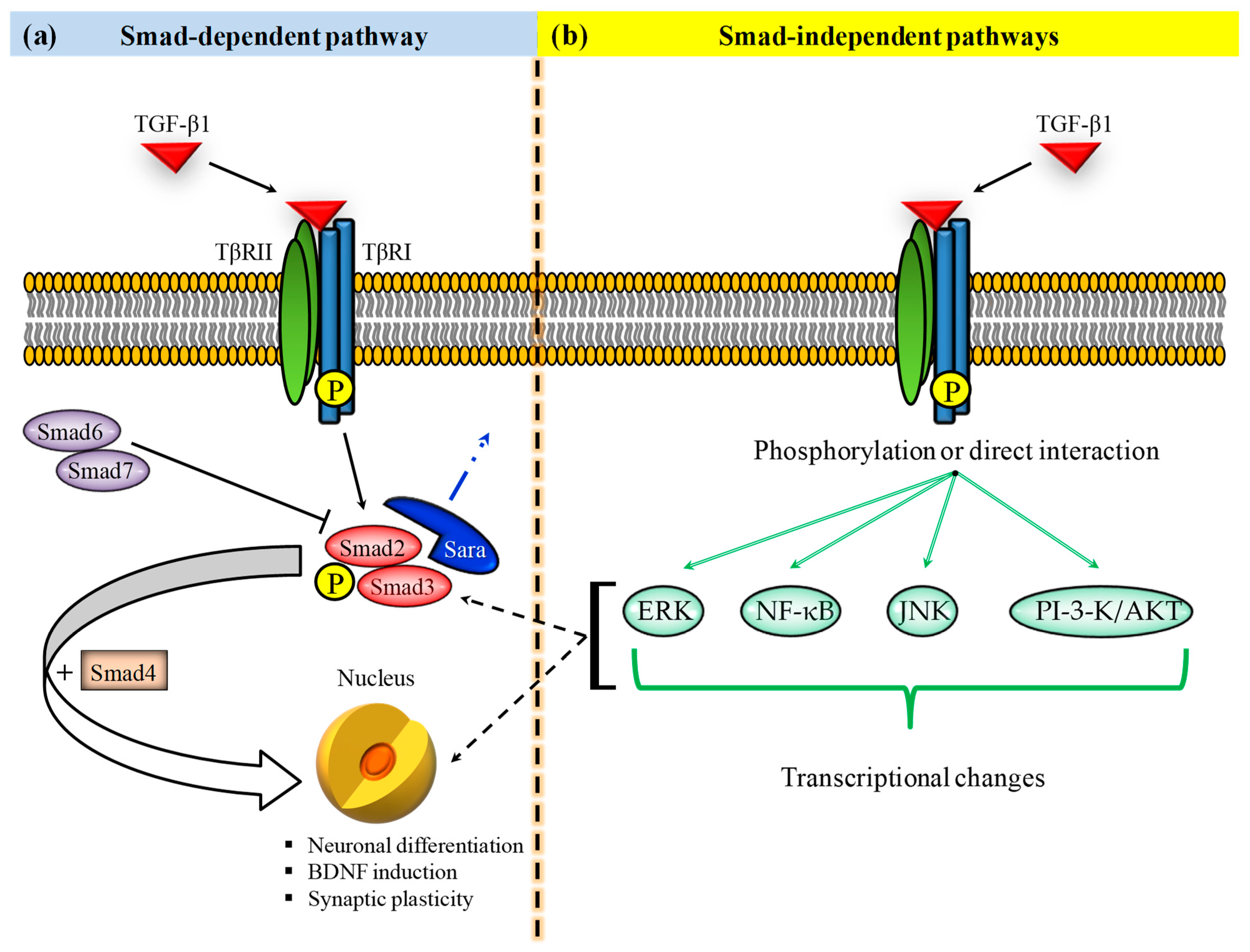
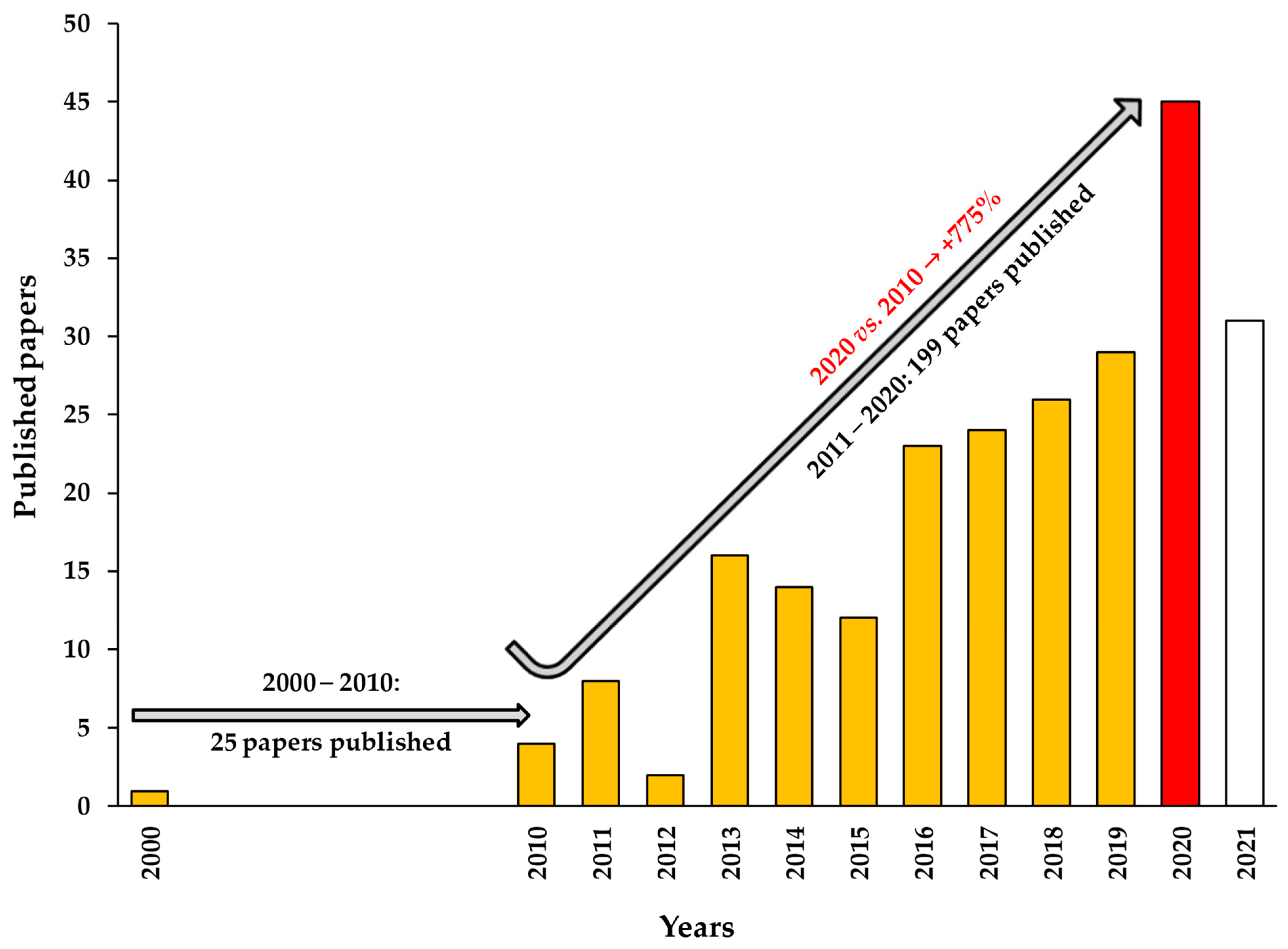
2. TGF-β1 in Cognitive Disorders
3. Nutraceuticals Targeting TGF-β1 Pathway: Evidence from Preclinical Studies
3.1. Medicinal Herbs
3.2. Omega-3 and Omega-6 Fatty Acids
3.3. Multifunctional Nutraceuticals Able to Target TGF-β1 Signaling: Focus on Carnosine and Its Therapeutic Potential in Cognitive Disorders
Increasing Carnosine Delivery and Its Bioavailability: Focus on Vesicular, Nanoparticulates Systems and Derivatives
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- De Felice, S. The Nutraceutical Revolution: Fueling a Powerful, New International Market. 1989. Available online: https://fimdefelice.org (accessed on 5 May 2021).
- González-Sarrías, A.; Larrosa, M.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. Nutraceuticals for older people: Facts, fictions and gaps in knowledge. Maturitas 2013, 75, 313–334. [Google Scholar] [CrossRef]
- Saiwal, N.; Dahiya, M.; Dureja, H. Nutraceutical insight into vegetables and their potential for nutrition mediated healthcare. Curr. Nutr. Food Sci. 2019, 15, 441–453. [Google Scholar] [CrossRef]
- Medalia, A.; Revheim, N. Dealing with Cognitive Dysfunction Associated with Psychiatric Disabilities: A Handbook for Families and Friends of Individuals with Psychiatric Disorders; New York State Office of Mental Health: New York, NY, USA, 2002. [Google Scholar]
- Al Dahhan, N.Z.; De Felice, F.G.; Munoz, D.P. Potentials and pitfalls of cross-translational models of cognitive impairment. Front. Behav. Neurosci. 2019, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Clements, S.; Corney, S.; Humin, Y.; Karmas, R.; Henderson, C. Cognitive Functioning: Supporting People with Mental Health Conditions and Cognitive Impairment. 2015. Available online: www.mhcc.org.au (accessed on 5 May 2021).
- Godos, J.; Caraci, F.; Castellano, S.; Currenti, W.; Galvano, F.; Ferri, R.; Grosso, G. Association between dietary flavonoids intake and cognitive function in an Italian cohort. Biomolecules 2020, 10, 1300. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P. The impact of fruit flavonoids on memory and cognition. Br. J. Nutr. 2010, 104 (Suppl. 3), S40–S47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godos, J.; Currenti, W.; Angelino, D.; Mena, P.; Castellano, S.; Caraci, F.; Galvano, F.; Del Rio, D.; Ferri, R.; Grosso, G. Diet and mental health: Review of the recent updates on molecular mechanisms. Antioxidants 2020, 9, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irwin, M.R.; Olmstead, R.; Carroll, J.E. Sleep disturbance, sleep duration, and inflammation: A systematic review and meta-analysis of cohort studies and experimental sleep deprivation. Biol. Psychiatry 2016, 80, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Philip, P.; Sagaspe, P.; Taillard, J.; Mandon, C.; Constans, J.; Pourtau, L.; Pouchieu, C.; Angelino, D.; Mena, P.; Martini, D. Acute intake of a grape and blueberry polyphenol-rich extract ameliorates cognitive performance in healthy young adults during a sustained cognitive effort. Antioxidants 2019, 8, 650. [Google Scholar] [CrossRef] [Green Version]
- Valls-Pedret, C.; Lamuela-Raventós, R.M.; Medina-Remón, A.; Quintana, M.; Corella, D.; Pintó, X.; Martínez-González, M.Á.; Estruch, R.; Ros, E. Polyphenol-rich foods in the mediterranean diet are associated with better cognitive function in elderly subjects at high cardiovascular risk. J. Alzheimer Dis. 2012, 29, 773–782. [Google Scholar] [CrossRef] [Green Version]
- Angelino, D.; Godos, J.; Ghelfi, F.; Tieri, M.; Titta, L.; Lafranconi, A.; Marventano, S.; Alonzo, E.; Gambera, A.; Sciacca, S.; et al. Fruit and vegetable consumption and health outcomes: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2019, 70, 652–667. [Google Scholar] [CrossRef]
- Tieri, M.; Ghelfi, F.; Vitale, M.; Vetrani, C.; Marventano, S.; Lafranconi, A.; Godos, J.; Titta, L.; Gambera, A.; Alonzo, E.; et al. Whole grain consumption and human health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2020, 71, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Martini, D.; Godos, J.; Marventano, S.; Tieri, M.; Ghelfi, F.; Titta, L.; Lafranconi, A.; Trigueiro, H.; Gambera, A.; Alonzo, E.; et al. Nut and legume consumption and human health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2021. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Annweiler, C.; Herrmann, F.R.; Fantino, B.; Brugg, B.; Beauchet, O. Effectiveness of the combination of memantine plus vitamin D on cognition in patients with Alzheimer disease: A pre-post pilot study. Cogn. Behav. Neurol. 2012, 25, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Winnick, R.; Winnick, T. Carnosine-anserine synthetase of muscle I. Preparation and properties of a soluble enyzme from chick muscle. Biochim. Biophys. Acta 1959, 31, 47–55. [Google Scholar] [CrossRef]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G.; et al. Modulation of pro-oxidant and pro-inflammatory activities of m1 macrophages by the natural dipeptide carnosine. Int. J. Mol. Sci. 2020, 21, 776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, K.; Mousa, A.; de Courten, B. Effects of supplementation with carnosine and other histidine-containing dipeptides on chronic disease risk factors and outcomes: Protocol for a systematic review of randomised controlled trials. BMJ Open 2018, 8, e020623. [Google Scholar] [CrossRef] [PubMed]
- Rokicki, J.; Li, L.; Imabayashi, E.; Kaneko, J.; Hisatsune, T.; Matsuda, H. Daily carnosine and anserine supplementation alters verbal episodic memory and resting state network connectivity in healthy elderly adults. Front. Aging Neurosci. 2015, 7, 219. [Google Scholar] [CrossRef] [Green Version]
- Szcześniak, D.; Budzeń, S.; Kopeć, W.; Rymaszewska, J. Anserine and carnosine supplementation in the elderly: Effects on cognitive functioning and physical capacity. Arch. Gerontol. Geriatr. 2014, 59, 485–490. [Google Scholar] [CrossRef]
- Hisatsune, T.; Kaneko, J.; Kurashige, H.; Cao, Y.; Satsu, H.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H. Effect of anserine/carnosine supplementation on verbal episodic memory in elderly people. J. Alzheimer Dis. 2016, 50, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Baraniuk, J.N.; El-Amin, S.; Corey, R.; Rayhan, R.; Timbol, C. Carnosine treatment for gulf war illness: A randomized controlled trial. Glob. J. Health Sci. 2013, 5, 69–81. [Google Scholar] [CrossRef]
- Masuoka, N.; Yoshimine, C.; Hori, M.; Tanaka, M.; Asada, T.; Abe, K.; Hisatsune, T. Effects of anserine/carnosine supplementation on mild cognitive impairment with apoe4. Nutrients 2019, 11, 1626. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Godos, J.; Castellano, S.; Micek, A.; Murabito, P.; Galvano, F.; Ferri, R.; Grosso, G.; Caraci, F. The therapeutic potential of carnosine/anserine supplementation against cognitive decline: A systematic review with meta-analysis. Biomedicines 2021, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Cornelli, U. Treatment of Alzheimer’s disease with a cholinesterase inhibitor combined with antioxidants. Neurodegener. Dis. 2010, 7, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, A.N.; Harrington, R.J.; Tsai, A.; Liao, P.; Harrington, M.G. Free amino acid and dipeptide changes in the body fluids from Alzheimer’s disease subjects. Amino Acids 2007, 32, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Vivien, D.; Ali, C. Transforming growth factor-β signalling in brain disorders. Cytokine Growth Factor Rev. 2006, 17, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Bosco, P.; Signorelli, M.; Spada, R.S.; Cosentino, F.I.; Toscano, G.; Bonforte, C.; Muratore, S.; Prestianni, G.; Panerai, S. The cc genotype of transforming growth factor-β1 increases the risk of late-onset Alzheimer’s disease and is associated with ad-related depression. Eur. Neuropsychopharmacol. 2012, 22, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Wrighton, K.H.; Lin, X.; Feng, X.H. Phospho-control of tgf-beta superfamily signaling. Cell Res. 2009, 19, 8–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuzillet, C.; de Gramont, A.; Tijeras-Raballand, A.; de Mestier, L.; Cros, J.; Faivre, S.; Raymond, E. Perspectives of tgf-β inhibition in pancreatic and hepatocellular carcinomas. Oncotarget 2014, 5, 78–94. [Google Scholar] [CrossRef] [Green Version]
- Massagué, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.H.; Derynck, R. Specificity and versatility in tgf-beta signaling through smads. Annu. Rev. Cell Dev. Biol. 2005, 21, 659–693. [Google Scholar] [CrossRef] [Green Version]
- Liu, F. Receptor-regulated smads in tgf-beta signaling. Front. Biosci. 2003, 8, s1280–s1303. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.E. Smad-dependent and smad-independent pathways in tgf-beta family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- König, H.G.; Kögel, D.; Rami, A.; Prehn, J.H. Tgf-β1 activates two distinct type i receptors in neurons: Implications for neuronal nf-κb signaling. J. Cell Biol. 2005, 168, 1077–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.E. Non-smad signaling pathways of the tgf-β family. Cold Spring Harb. Perspect. Biol. 2017, 9, a022129. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Battaglia, G.; Busceti, C.; Biagioni, F.; Mastroiacovo, F.; Bosco, P.; Drago, F.; Nicoletti, F.; Sortino, M.A.; Copani, A. Tgf-beta 1 protects against abeta-neurotoxicity via the phosphatidylinositol-3-kinase pathway. Neurobiol. Dis. 2008, 30, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Spampinato, S.; Sortino, M.A.; Bosco, P.; Battaglia, G.; Bruno, V.; Drago, F.; Nicoletti, F.; Copani, A. Dysfunction of tgf-β1 signaling in Alzheimer’s disease: Perspectives for neuroprotection. Cell Tissue Res. 2012, 347, 291–301. [Google Scholar] [CrossRef]
- Meyers, E.A.; Kessler, J.A. Tgf-β family signaling in neural and neuronal differentiation, development, and function. Cold Spring Harb. Perspect. Biol. 2017, 9, a022244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sometani, A.; Kataoka, H.; Nitta, A.; Fukumitsu, H.; Nomoto, H.; Furukawa, S. Transforming growth factor-beta1 enhances expression of brain-derived neurotrophic factor and its receptor, trkb, in neurons cultured from rat cerebral cortex. J. Neurosci. Res. 2001, 66, 369–376. [Google Scholar] [CrossRef]
- Caraci, F.; Spampinato, S.F.; Morgese, M.G.; Tascedda, F.; Salluzzo, M.G.; Giambirtone, M.C.; Caruso, G.; Munafò, A.; Torrisi, S.A.; Leggio, G.M.; et al. Neurobiological links between depression and ad: The role of tgf-β1 signaling as a new pharmacological target. Pharm. Res. 2018, 130, 374–384. [Google Scholar] [CrossRef]
- Caraci, F.; Gulisano, W.; Guida, C.A.; Impellizzeri, A.A.; Drago, F.; Puzzo, D.; Palmeri, A. A key role for tgf-β1 in hippocampal synaptic plasticity and memory. Sci. Rep. 2015, 5, 11252. [Google Scholar] [CrossRef] [Green Version]
- Koeglsperger, T.; Li, S.; Brenneis, C.; Saulnier, J.L.; Mayo, L.; Carrier, Y.; Selkoe, D.J.; Weiner, H.L. Impaired glutamate recycling and glun2b-mediated neuronal calcium overload in mice lacking tgf-β1 in the cns. Glia 2013, 61, 985–1002. [Google Scholar] [CrossRef] [Green Version]
- Tesseur, I.; Zou, K.; Esposito, L.; Bard, F.; Berber, E.; Can, J.V.; Lin, A.H.; Crews, L.; Tremblay, P.; Mathews, P.; et al. Deficiency in neuronal tgf-beta signaling promotes neurodegeneration and Alzheimer’s pathology. J. Clin. Investig. 2006, 116, 3060–3069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueberham, U.; Hilbrich, I.; Ueberham, E.; Rohn, S.; Glöckner, P.; Dietrich, K.; Brückner, M.K.; Arendt, T. Transcriptional control of cell cycle-dependent kinase 4 by smad proteins—Implications for Alzheimer’s disease. Neurobiol. Aging 2012, 33, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.G.; Musso, N.; Giambirtone, M.; Grasso, M.; Spampinato, S.F.; Merlo, S.; Drago, F.; Lazzarino, G.; Sortino, M.A.; et al. Carnosine prevents aβ-induced oxidative stress and inflammation in microglial cells: A key role of tgf-β1. Cells 2019, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Torrisi, S.A.; Geraci, F.; Tropea, M.R.; Grasso, M.; Caruso, G.; Fidilio, A.; Musso, N.; Sanfilippo, G.; Tascedda, F.; Palmeri, A.; et al. Fluoxetine and vortioxetine reverse depressive-like phenotype and memory deficits induced by aβ(1-42) oligomers in mice: A key role of transforming growth factor-β1. Front. Pharm. 2019, 10, 693. [Google Scholar] [CrossRef]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.M.; Kim, Y.K. The role of il-12 and tgf-beta1 in the pathophysiology of major depressive disorder. Int. Immunopharmacol. 2006, 6, 1298–1304. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Xu, H.; Zhu, S.; Wang, H.; He, J.; Zhang, H.; Guo, H.; Kong, J.; Huang, Q.; et al. Fluoxetine improves behavioral performance by suppressing the production of soluble β-amyloid in app/ps1 mice. Curr. Alzheimer Res. 2014, 11, 672–680. [Google Scholar] [CrossRef]
- Caraci, F.; Tascedda, F.; Merlo, S.; Benatti, C.; Spampinato, S.F.; Munafò, A.; Leggio, G.M.; Nicoletti, F.; Brunello, N.; Drago, F.; et al. Fluoxetine prevents aβ(1-42)-induced toxicity via a paracrine signaling mediated by transforming-growth-factor-β1. Front. Pharm. 2016, 7, 389. [Google Scholar] [CrossRef] [Green Version]
- Dobolyi, A.; Vincze, C.; Pál, G.; Lovas, G. The neuroprotective functions of transforming growth factor beta proteins. Int. J. Mol. Sci. 2012, 13, 8219–8258. [Google Scholar] [CrossRef]
- Lacmann, A.; Hess, D.; Gohla, G.; Roussa, E.; Krieglstein, K. Activity-dependent release of transforming growth factor-beta in a neuronal network in vitro. Neuroscience 2007, 150, 647–657. [Google Scholar] [CrossRef]
- Battaglia, G.; Cannella, M.; Riozzi, B.; Orobello, S.; Maat-Schieman, M.L.; Aronica, E.; Busceti, C.L.; Ciarmiello, A.; Alberti, S.; Amico, E.; et al. Early defect of transforming growth factor β1 formation in huntington’s disease. J. Cell Mol. Med. 2011, 15, 555–571. [Google Scholar] [CrossRef] [Green Version]
- da Cunha, A.; Jefferson, J.A.; Jackson, R.W.; Vitković, L. Glial cell-specific mechanisms of tgf-beta 1 induction by il-1 in cerebral cortex. J. Neuroimmunol. 1993, 42, 71–85. [Google Scholar] [CrossRef]
- Yao, H.; Zhang, Y.; Shu, H.; Xie, B.; Tao, Y.; Yuan, Y.; Shang, Y.; Yuan, S.; Zhang, J. Hyperforin promotes post-stroke neuroangiogenesis via astrocytic il-6-mediated negative immune regulation in the ischemic brain. Front. Cell Neurosci. 2019, 13, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, G. The role of proteases in transforming growth factor-beta activation. Int. J. Biochem. Cell Biol. 2008, 40, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Matias, I.; Diniz, L.P.; Buosi, A.; Neves, G.; Stipursky, J.; Gomes, F.C.A. Flavonoid hesperidin induces synapse formation and improves memory performance through the astrocytic tgf-β1. Front. Aging Neurosci. 2017, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Matias, I.; Buosi, A.S.; Gomes, F.C. Functions of flavonoids in the central nervous system: Astrocytes as targets for natural compounds. Neurochem. Int. 2016, 95, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; van Praag, H.; Gage, F.H. Activity-dependent regulation of neuronal plasticity and self repair. Prog. Brain Res. 2000, 127, 35–48. [Google Scholar]
- Cobianchi, S.; Arbat-Plana, A.; Lopez-Alvarez, V.M.; Navarro, X. Neuroprotective effects of exercise treatments after injury: The dual role of neurotrophic factors. Curr. Neuropharmacol. 2017, 15, 495–518. [Google Scholar] [CrossRef] [Green Version]
- Baj, G.; D’Alessandro, V.; Musazzi, L.; Mallei, A.; Sartori, C.R.; Sciancalepore, M.; Tardito, D.; Langone, F.; Popoli, M.; Tongiorgi, E. Physical exercise and antidepressants enhance bdnf targeting in hippocampal ca3 dendrites: Further evidence of a spatial code for bdnf splice variants. Neuropsychopharmacology 2012, 37, 1600–1611. [Google Scholar] [CrossRef] [Green Version]
- Guerrera, C.S.; Furneri, G.; Grasso, M.; Caruso, G.; Castellano, S.; Drago, F.; Di Nuovo, S.; Caraci, F. Antidepressant drugs and physical activity: A possible synergism in the treatment of major depression? Front. Psychol. 2020, 11, 857. [Google Scholar] [CrossRef] [PubMed]
- Szymura, J.; Kubica, J.; Wiecek, M.; Pera, J. The immunomodulary effects of systematic exercise in older adults and people with parkinson’s disease. J. Clin. Med. 2020, 9, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, S. Nanoparticle-based delivery of phytochemical compounds against major maladies: Cancer, diabetes, and cardiovascular disease. In Plant-Derived Bioactives; Springer: Singapore, 2020; pp. 591–619. [Google Scholar]
- Sarris, J. Herbal medicines in the treatment of psychiatric disorders: 10-year updated review. Phytother. Res. 2018, 32, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Phillipson, J.D. Phytochemistry and medicinal plants. Phytochemistry 2001, 56, 237–243. [Google Scholar] [CrossRef]
- Vacca, R.A.; Valenti, D.; Caccamese, S.; Daglia, M.; Braidy, N.; Nabavi, S.M. Plant polyphenols as natural drugs for the management of down syndrome and related disorders. Neurosci. Biobehav. Rev. 2016, 71, 865–877. [Google Scholar] [CrossRef]
- Mao, T.K.; Van De Water, J.; Keen, C.L.; Schmitz, H.H.; Gershwin, M.E. Cocoa flavonols and procyanidins promote transforming growth factor-beta1 homeostasis in peripheral blood mononuclear cells. Exp. Biol. Med. 2003, 228, 93–99. [Google Scholar] [CrossRef]
- Matias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte heterogeneity: Impact to brain aging and disease. Front. Aging Neurosci. 2019, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Kasper, S.; Caraci, F.; Forti, B.; Drago, F.; Aguglia, E. Efficacy and tolerability of hypericum extract for the treatment of mild to moderate depression. Eur. Neuropsychopharmacol. 2010, 20, 747–765. [Google Scholar] [CrossRef]
- Ben-Eliezer, D.; Yechiam, E. Hypericum perforatum as a cognitive enhancer in rodents: A meta-analysis. Sci. Rep. 2016, 6, 35700. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Wang, F.; Xiu, C.; Zhang, J.; Li, Y. Hypericum perforatum extract attenuates behavioral, biochemical, and neurochemical abnormalities in aluminum chloride-induced Alzheimer’s disease rats. Biomed. Pharm. 2017, 91, 931–937. [Google Scholar] [CrossRef]
- Trofimiuk, E.; Walesiuk, A.; Braszko, J.J. St John’s wort (Hypericum perforatum) diminishes cognitive impairment caused by the chronic restraint stress in rats. Pharm. Res. 2005, 51, 239–246. [Google Scholar] [CrossRef]
- Süntar, I.P.; Akkol, E.K.; Yilmazer, D.; Baykal, T.; Kirmizibekmez, H.; Alper, M.; Yeşilada, E. Investigations on the in vivo wound healing potential of Hypericum perforatum L. J. Ethnopharmacol. 2010, 127, 468–477. [Google Scholar] [CrossRef]
- Sayar, H.; Gergerlioglu, N.; Seringec, N.; Ozturk, P.; Bulbuloglu, E.; Karabay, G. Comparison of efficacy of topical phenytoin with hypericin in second-degree burn wound healing: An experimental study in rats. Med. Sci. Monit. Basic Res. 2014, 20, 36–46. [Google Scholar]
- Yechiam, E.; Ben-Eliezer, D.; Ashby, N.J.S.; Bar-Shaked, M. The acute effect of Hypericum perforatum on short-term memory in healthy adults. Psychopharmacology 2019, 236, 613–623. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, P.; Liu, H.; Yao, H.; Yao, S.; Yuan, S.Y.; Zhang, J.C. Hyperforin improves post-stroke social isolation-induced exaggeration of psd and psa via tgf-β. Int. J. Mol. Med. 2019, 43, 413–425. [Google Scholar] [CrossRef]
- Goyarzu, P.; Malin, D.H.; Lau, F.C.; Taglialatela, G.; Moon, W.D.; Jennings, R.; Moy, E.; Moy, D.; Lippold, S.; Shukitt-Hale, B.; et al. Blueberry supplemented diet: Effects on object recognition memory and nuclear factor-kappa b levels in aged rats. Nutr. Neurosci. 2004, 7, 75–83. [Google Scholar] [CrossRef]
- Joseph, J.A.; Denisova, N.A.; Arendash, G.; Gordon, M.; Diamond, D.; Shukitt-Hale, B.; Morgan, D. Blueberry supplementation enhances signaling and prevents behavioral deficits in an Alzheimer disease model. Nutr. Neurosci. 2003, 6, 153–162. [Google Scholar] [CrossRef]
- Rubio-Perez, J.M.; Morillas-Ruiz, J.M. A review: Inflammatory process in Alzheimer’s disease, role of cytokines. Sci. World J. 2012, 2012, 756357. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Nowak, G.; Caso, J.R.; Leza, J.C.; Song, C.; Kubera, M.; Klein, H.; Galecki, P.; Noto, C.; Glaab, E.; et al. Toward omics-based, systems biomedicine, and path and drug discovery methodologies for depression-inflammation research. Mol. Neurobiol. 2016, 53, 2927–2935. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Li, C.; Zug, C.; Qu, H.; Schluesener, H.; Zhang, Z. Hesperidin ameliorates behavioral impairments and neuropathology of transgenic app/ps1 mice. Behav. Brain Res. 2015, 281, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-X.; Deng, Y.-Y.; Li, F.; Liu, B.; Liu, H.-Y.; Shi, J.-S.; Gong, Q.-H. Icariin, a major constituent of flavonoids from Epimedium brevicornum, protects against cognitive deficits induced by chronic brain hypoperfusion via its anti-amyloidogenic effect in rats. Pharmacol. Biochem. Behav. 2015, 138, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Angeloni, C.; Barbalace, M.C.; Hrelia, S. Icariin and its metabolites as potential protective phytochemicals against Alzheimer’s disease. Front. Pharm. 2019, 10, 271. [Google Scholar] [CrossRef] [PubMed]
- Sheng, C.; Xu, P.; Zhou, K.; Deng, D.; Zhang, C.; Wang, Z. Icariin attenuates synaptic and cognitive deficits in an aβ(1-42)-induced rat model of Alzheimer’s disease. Biomed. Res. Int. 2017, 2017, 7464872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.P. Omega-3 fatty acids in psychiatry: A review. Ann. Clin. Psychiatry 2000, 12, 159–165. [Google Scholar] [CrossRef]
- Moriguchi, T.; Salem, N., Jr. Recovery of brain docosahexaenoate leads to recovery of spatial task performance. J. Neurochem. 2003, 87, 297–309. [Google Scholar] [CrossRef]
- Heude, B.; Ducimetière, P.; Berr, C. Cognitive decline and fatty acid composition of erythrocyte membranes—The eva study. Am. J. Clin. Nutr. 2003, 77, 803–808. [Google Scholar] [CrossRef] [Green Version]
- Helland, I.B.; Smith, L.; Saarem, K.; Saugstad, O.D.; Drevon, C.A. Maternal supplementation with very-long-chain n-3 fatty acids during pregnancy and lactation augments children’s iq at 4 years of age. Pediatrics 2003, 111, e39–e44. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, M.; Katakura, M.; Tanabe, Y.; Al Mamun, A.; Inoue, T.; Hossain, S.; Arita, M.; Shido, O. N-3 fatty acids effectively improve the reference memory-related learning ability associated with increased brain docosahexaenoic acid-derived docosanoids in aged rats. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 203–209. [Google Scholar] [CrossRef]
- Loef, M.; Walach, H. The omega-6/omega-3 ratio and dementia or cognitive decline: A systematic review on human studies and biological evidence. J. Nutr. Gerontol. Geriatr. 2013, 32, 1–23. [Google Scholar] [CrossRef]
- Takeuchi, T.; Fukumoto, Y.; Harada, E. Influence of a dietary n-3 fatty acid deficiency on the cerebral catecholamine contents, eeg and learning ability in rat. Behav. Brain Res. 2002, 131, 193–203. [Google Scholar] [CrossRef]
- Gamoh, S.; Hashimoto, M.; Sugioka, K.; Hossain, M.S.; Hata, N.; Misawa, Y.; Masumura, S. Chronic administration of docosahexaenoic acid improves reference memory-related learning ability in young rats. Neuroscience 1999, 93, 237–241. [Google Scholar] [CrossRef]
- Minami, M.; Kimura, S.; Endo, T.; Hamaue, N.; Hirafuji, M.; Togashi, H.; Matsumoto, M.; Yoshioka, M.; Saito, H.; Watanabe, S.; et al. Dietary docosahexaenoic acid increases cerebral acetylcholine levels and improves passive avoidance performance in stroke-prone spontaneously hypertensive rats. Pharm. Biochem. Behav. 1997, 58, 1123–1129. [Google Scholar] [CrossRef]
- Fontani, G.; Corradeschi, F.; Felici, A.; Alfatti, F.; Migliorini, S.; Lodi, L. Cognitive and physiological effects of omega-3 polyunsaturated fatty acid supplementation in healthy subjects. Eur. J. Clin. Investig. 2005, 35, 691–699. [Google Scholar] [CrossRef]
- Nunes, B.; Pinho, C.; Sousa, C.; Melo, A.R.; Bandarra, N.; Silva, M.C. Relevance of omega-3 and omega-6/omega-3 ratio in preventing cognitive impairment. Acta Med. Port. 2017, 30, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.J.; Spite, M. Resolvins: Anti-inflammatory and proresolving mediators derived from omega-3 polyunsaturated fatty acids. Annu. Rev. Nutr. 2012, 32, 203–227. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, K. Fatty acids as modulators of the immune response. Annu. Rev. Nutr. 2006, 26, 45–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrucci, L.; Cherubini, A.; Bandinelli, S.; Bartali, B.; Corsi, A.; Lauretani, F.; Martin, A.; Andres-Lacueva, C.; Senin, U.; Guralnik, J.M. Relationship of plasma polyunsaturated fatty acids to circulating inflammatory markers. J. Clin. Endocrinol. Metab. 2006, 91, 439–446. [Google Scholar] [CrossRef]
- Sharma, A.; Belna, J.; Espat, J.; Rodriguez, G.; Cannon, V.T.; Hurteau, J.A. Effects of omega-3 fatty acids on components of the transforming growth factor beta-1 pathway: Implication for dietary modification and prevention in ovarian cancer. Am. J. Obstet. Gynecol. 2009, 200, 516-e1. [Google Scholar] [CrossRef]
- Han, S.C.; Koo, D.H.; Kang, N.J.; Yoon, W.J.; Kang, G.J.; Kang, H.K.; Yoo, E.S. Docosahexaenoic acid alleviates atopic dermatitis by generating tregs and il-10/tgf-β-modified macrophages via a tgf-β-dependent mechanism. J. Investig. Dermatol. 2015, 135, 1556–1564. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Feng, Z.P.; Peng, H.Y.; Fu, P. Omega-3 polyunsaturated fatty acids alleviate adenine-induced chronic renal failure via regulating ros production and tgf-β/smad pathway. Eur. Rev. Med. Pharm. Sci. 2018, 22, 5024–5032. [Google Scholar]
- Hida, M.; Fujita, H.; Ishikura, K.; Omori, S.; Hoshiya, M.; Awazu, M. Eicosapentaenoic acid inhibits pdgf-induced mitogenesis and cyclin d1 expression via tgf-beta in mesangial cells. J. Cell. Physiol. 2003, 196, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Thienprasert, A.; Samuhaseneetoo, S.; Popplestone, K.; West, A.L.; Miles, E.A.; Calder, P.C. Fish oil n-3 polyunsaturated fatty acids selectively affect plasma cytokines and decrease illness in thai schoolchildren: A randomized, double-blind, placebo-controlled intervention trial. J. Pediatr. 2009, 154, 391–395. [Google Scholar] [CrossRef]
- Krauss-Etschmann, S.; Hartl, D.; Rzehak, P.; Heinrich, J.; Shadid, R.; Ramírez-Tortosa, M.D.C.; Campoy, C.; Pardillo, S.; Schendel, D.J.; Decsi, T.; et al. Decreased cord blood il-4, il-13, and ccr4 and increased tgf-beta levels after fish oil supplementation of pregnant women. J. Allergy Clin. Immunol. 2008, 121, 464–470.e466. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, S.; Shimada, T.; Sugioka, K.; Yamasaki, H.; Fujii, Y.; Ishibashi, Y.; Oka, J.; Shido, O. Docosahexaenoic acid provides protection from impairment of learning ability in Alzheimer’s disease model rats. J. Neurochem. 2002, 81, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Gamoh, S.; Hashimoto, M.; Hossain, S.; Masumura, S. Chronic administration of docosahexaenoic acid improves the performance of radial arm maze task in aged rats. Clin. Exp. Pharm. Physiol. 2001, 28, 266–270. [Google Scholar] [CrossRef]
- Cole, G.M.; Lim, G.P.; Yang, F.; Teter, B.; Begum, A.; Ma, Q.; Harris-White, M.E.; Frautschy, S.A. Prevention of Alzheimer’s disease: Omega-3 fatty acid and phenolic anti-oxidant interventions. Neurobiol. Aging 2005, 26 (Suppl. 1), 133–136. [Google Scholar] [CrossRef] [Green Version]
- Lukiw, W.J.; Cui, J.G.; Marcheselli, V.L.; Bodker, M.; Botkjaer, A.; Gotlinger, K.; Serhan, C.N.; Bazan, N.G. A role for docosahexaenoic acid-derived neuroprotectin d1 in neural cell survival and Alzheimer disease. J. Clin. Investig. 2005, 115, 2774–2783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, M.; Li, Y.; Tang, H.; Zhang, C.; Li, W.; Zhang, Y.; Li, Y.; Zhao, Y.; Song, C. Endogenous omega (n)-3 fatty acids in fat-1 mice attenuated depression-like behavior, imbalance between microglial m1 and m2 phenotypes, and dysfunction of neurotrophins induced by lipopolysaccharide administration. Nutrients 2018, 10, 1351. [Google Scholar] [CrossRef] [Green Version]
- McNamara, R.K.; Jandacek, R.; Rider, T.; Tso, P.; Chu, W.J.; Weber, W.A.; Welge, J.A.; Strawn, J.R.; Adler, C.M.; DelBello, M.P. Effects of fish oil supplementation on prefrontal metabolite concentrations in adolescents with major depressive disorder: A preliminary 1h mrs study. Nutr. Neurosci. 2016, 19, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Prokopieva, V.D.; Yarygina, E.G.; Bokhan, N.A.; Ivanova, S.A. Use of carnosine for oxidative stress reduction in different pathologies. Oxidative Med. Cell. Longev. 2016, 2016, 2939087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, M.; Kobayashi, N.; Sugizaki, T.; Shimoda, M.; Kawahara, M.; Tanaka, K.-I. Carnosine suppresses neuronal cell death and inflammation induced by 6-hydroxydopamine in an in vitro model of Parkinson’s disease. PLoS ONE 2020, 15, e0240448. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.G.; Fidilio, A.; O’Donnell, F.; Musso, N.; Lazzarino, G.; Grasso, M.; Amorini, A.M.; Tascedda, F.; Bucolo, C.; et al. Carnosine decreases pma-induced oxidative stress and inflammation in murine macrophages. Antioxidants 2019, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fresta, C.G.; Hogard, M.L.; Caruso, G.; Melo Costa, E.E.; Lazzarino, G.; Lunte, S.M. Monitoring carnosine uptake by raw 264.7 macrophage cells using microchip electrophoresis with fluorescence detection. Anal. Methods 2017, 9, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Benatti, C.; Musso, N.; Fresta, C.G.; Fidilio, A.; Spampinato, G.; Brunello, N.; Bucolo, C.; Drago, F.; Lunte, S.M.; et al. Carnosine protects macrophages against the toxicity of aβ1-42 oligomers by decreasing oxidative stress. Biomedicines 2021, 9, 477. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.G.; Grasso, M.; Santangelo, R.; Lazzarino, G.; Lunte, S.M.; Caraci, F. Inflammation as the common biological link between depression and cardiovascular diseases: Can carnosine exert a protective role? Curr. Med. Chem. 2020, 27, 1782–1800. [Google Scholar] [CrossRef]
- Famenini, S.; Rigali, E.A.; Olivera-Perez, H.M.; Dang, J.; Chang, M.T.; Halder, R.; Rao, R.V.; Pellegrini, M.; Porter, V.; Bredesen, D.; et al. Increased intermediate m1-m2 macrophage polarization and improved cognition in mild cognitive impairment patients on ω-3 supplementation. FASEB J. 2017, 31, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Pinto, B.; Morelli, G.; Rastogi, M.; Savardi, A.; Fumagalli, A.; Petretto, A.; Bartolucci, M.; Varea, E.; Catelani, T.; Contestabile, A.; et al. Rescuing over-activated microglia restores cognitive performance in juvenile animals of the dp(16) mouse model of down syndrome. Neuron 2020, 108, 887–904.e12. [Google Scholar] [CrossRef] [PubMed]
- Malpetti, M.; Kievit, R.A.; Passamonti, L.; Jones, P.S.; Tsvetanov, K.A.; Rittman, T.; Mak, E.; Nicastro, N.; Bevan-Jones, W.R.; Su, L.; et al. Microglial activation and tau burden predict cognitive decline in Alzheimer’s disease. Brain 2020, 143, 1588–1602. [Google Scholar] [CrossRef]
- Caruso, G.; Caraci, F.; Jolivet, R.B. Pivotal role of carnosine in the modulation of brain cells activity: Multimodal mechanism of action and therapeutic potential in neurodegenerative disorders. Prog. Neurobiol. 2019, 175, 35–53. [Google Scholar] [CrossRef]
- Ansurudeen, I.; Sunkari, V.G.; Grünler, J.; Peters, V.; Schmitt, C.P.; Catrina, S.B.; Brismar, K.; Forsberg, E.A. Carnosine enhances diabetic wound healing in the db/db mouse model of type 2 diabetes. Amino Acids 2012, 43, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Han, K.; Park, S.; Cho, H.; Lee, D.Y.; Kim, J.W.; Seo, J.A.; Kim, S.G.; Baik, S.H.; Park, Y.G.; et al. Incidence and risk factors for dementia in type 2 diabetes mellitus: A nationwide population-based study in Korea. Diabetes Metab. J. 2020, 44, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Distefano, D.A.; Parlascino, P.; Fresta, C.G.; Lazzarino, G.; Lunte, S.M.; Nicoletti, V.G. Receptor-mediated toxicity of human amylin fragment aggregated by short- and long-term incubations with copper ions. Mol. Cell. Biochem. 2017, 425, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.; Mousa, A.; Berk, M.; Chia, W.L.; Ukropec, J.; Majid, A.; Ukropcová, B.; de Courten, B. The potential of carnosine in brain-related disorders: A comprehensive review of current evidence. Nutrients 2019, 11, 1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everaert, I.; Mooyaart, A.; Baguet, A.; Zutinic, A.; Baelde, H.; Achten, E.; Taes, Y.; De Heer, E.; Derave, W. Vegetarianism, female gender and increasing age, but not cndp1 genotype, are associated with reduced muscle carnosine levels in humans. Amino Acids 2011, 40, 1221–1229. [Google Scholar] [CrossRef]
- Qiu, J.; Hauske, S.J.; Zhang, S.; Rodriguez-Niño, A.; Albrecht, T.; Pastene, D.O.; van den Born, J.; van Goor, H.; Ruf, S.; Kohlmann, M.; et al. Identification and characterisation of carnostatine (san9812), a potent and selective carnosinase (cn1) inhibitor with in vivo activity. Amino Acids 2019, 51, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Bermúdez, M.L.; Skelton, M.R.; Genter, M.B. Intranasal carnosine attenuates transcriptomic alterations and improves mitochondrial function in the thy1-asyn mouse model of Parkinson’s disease. Mol. Genet. Metab. 2018, 125, 305–313. [Google Scholar] [CrossRef]
- Jadhav, K.R.; Gambhire, M.N.; Shaikh, I.M.; Kadam, V.J.; Pisal, S.S. Nasal drug delivery system-factors affecting and applications. Curr. Drug Ther. 2007, 2, 27–38. [Google Scholar] [CrossRef]
- Vecchio, G.; La Mendola, D.; Rizzarelli, E. The synthesis and conformation of β-cyclodextrins functionalized with enantiomers of boc-carnosine. J. Supramol. Chem. 2001, 1, 87–95. [Google Scholar] [CrossRef]
- Lanza, V.; Bellia, F.; D’Agata, R.; Grasso, G.; Rizzarelli, E.; Vecchio, G. New glycoside derivatives of carnosine and analogs resistant to carnosinase hydrolysis: Synthesis and characterization of their copper(ii) complexes. J. Inorg. Biochem. 2011, 105, 181–188. [Google Scholar] [CrossRef]
- Bellia, F.; Oliveri, V.; Rizzarelli, E.; Vecchio, G. New derivative of carnosine for nanoparticle assemblies. Eur. J. Med. Chem. 2013, 70, 225–232. [Google Scholar] [CrossRef]
- Farid, R.M.; Gaafar, P.M.E.; Hazzah, H.A.; Helmy, M.W.; Abdallah, O.Y. Chemotherapeutic potential of l-carnosine from stimuli-responsive magnetic nanoparticles against breast cancer model. Nanomedicine 2020, 15, 891–911. [Google Scholar] [CrossRef] [PubMed]
- Khramtsov, P.; Barkina, I.; Kropaneva, M.; Bochkova, M.; Timganova, V.; Nechaev, A.; Byzov, I.; Zamorina, S.; Yermakov, A.; Rayev, M. Magnetic nanoclusters coated with albumin, casein, and gelatin: Size tuning, relaxivity, stability, protein corona, and application in nuclear magnetic resonance immunoassay. Nanomaterials 2019, 9, 1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durmus, Z.; Kavas, H.; Baykal, A.; Sozeri, H.; Alpsoy, L.; Çelik, S.; Toprak, M. Synthesis and characterization of l-carnosine coated iron oxide nanoparticles. J. Alloy. Compd. 2011, 509, 2555–2561. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, Y.; Wang, L.; Li, G.; Gao, J.; Wang, Y. Development of l-carnosine functionalized iron oxide nanoparticles loaded with dexamethasone for simultaneous therapeutic potential of blood brain barrier crossing and ischemic stroke treatment. Drug Deliv. 2021, 28, 380–389. [Google Scholar] [CrossRef]
- Maherani, B.; Arab-Tehrany, E.; Kheirolomoom, A.; Cleymand, F.; Linder, M. Influence of lipid composition on physicochemical properties of nanoliposomes encapsulating natural dipeptide antioxidant l-carnosine. Food Chem. 2012, 134, 632–640. [Google Scholar] [CrossRef]
- Maestrelli, F.; Landucci, E.; De Luca, E.; Nerli, G.; Bergonzi, M.C.; Piazzini, V.; Pellegrini-Giampietro, D.E.; Gullo, F.; Becchetti, A.; Tadini-Buoninsegni, F.; et al. Niosomal formulation of a lipoyl-carnosine derivative targeting trpa1 channels in brain. Pharmaceutics 2019, 11, 669. [Google Scholar] [CrossRef] [Green Version]
- Moulahoum, H.; Sanli, S.; Timur, S.; Zihnioglu, F. Potential effect of carnosine encapsulated niosomes in bovine serum albumin modifications. Int. J. Biol. Macromol. 2019, 137, 583–591. [Google Scholar] [CrossRef]
- Kim, E.S.; Kim, D.; Nyberg, S.; Poma, A.; Cecchin, D.; Jain, S.A.; Kim, K.A.; Shin, Y.J.; Kim, E.H.; Kim, M.; et al. Lrp-1 functionalized polymersomes enhance the efficacy of carnosine in experimental stroke. Sci. Rep. 2020, 10, 699. [Google Scholar] [CrossRef]
- Bellia, F.; Vecchio, G.; Rizzarelli, E. Carnosinases, their substrates and diseases. Molecules 2014, 19, 2299–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awan, Z.A.; Fahmy, U.A.; Badr-Eldin, S.M.; Ibrahim, T.S.; Asfour, H.Z.; Al-Rabia, M.W.; Alfarsi, A.; Alhakamy, N.A.; Abdulaal, W.H.; Al Sadoun, H.; et al. The enhanced cytotoxic and pro-apoptotic effects of optimized simvastatin-loaded emulsomes on mcf-7 breast cancer cells. Pharmaceutics 2020, 12, 597. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Delivery System | References | |
|---|---|---|
| Carnosine derivatives |
| [134] |
| [135] | |
| [136] | |
| Nanoparticulate systems |
| [137,138,139] |
| [140] | |
| Vesicular systems |
| [141] |
| [142,143] | |
| [144] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grasso, M.; Caruso, G.; Godos, J.; Bonaccorso, A.; Carbone, C.; Castellano, S.; Currenti, W.; Grosso, G.; Musumeci, T.; Caraci, F. Improving Cognition with Nutraceuticals Targeting TGF-β1 Signaling. Antioxidants 2021, 10, 1075. https://doi.org/10.3390/antiox10071075
Grasso M, Caruso G, Godos J, Bonaccorso A, Carbone C, Castellano S, Currenti W, Grosso G, Musumeci T, Caraci F. Improving Cognition with Nutraceuticals Targeting TGF-β1 Signaling. Antioxidants. 2021; 10(7):1075. https://doi.org/10.3390/antiox10071075
Chicago/Turabian StyleGrasso, Margherita, Giuseppe Caruso, Justyna Godos, Angela Bonaccorso, Claudia Carbone, Sabrina Castellano, Walter Currenti, Giuseppe Grosso, Teresa Musumeci, and Filippo Caraci. 2021. "Improving Cognition with Nutraceuticals Targeting TGF-β1 Signaling" Antioxidants 10, no. 7: 1075. https://doi.org/10.3390/antiox10071075
APA StyleGrasso, M., Caruso, G., Godos, J., Bonaccorso, A., Carbone, C., Castellano, S., Currenti, W., Grosso, G., Musumeci, T., & Caraci, F. (2021). Improving Cognition with Nutraceuticals Targeting TGF-β1 Signaling. Antioxidants, 10(7), 1075. https://doi.org/10.3390/antiox10071075