
Pressurized Hot Liquid Extraction with 15% v/v Glycerol-Water as An Effective Environment-Friendly Process to Obtain Durvillaea incurvata and Lessonia spicata Phlorotannin Extracts with Antioxidant and Antihyperglycemic Potential


 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Seaweed Samples
2.3. Liquid Extraction Methods: PHLE and Maceration with 60% Acetone
2.4. Total Solids Content of the PHLE Extracts
2.5. Total Polyphenol Content
2.6. Antioxidant Capacity
2.6.1. DPPH Radical Scavenging Activity (DPPH RSA)
2.6.2. Oxygen Radical Absorbance Capacity (ORAC)
2.7. Inhibition of Carbohydrate-Hydrolyzing Enzymes
2.7.1. Sample Preparation
2.7.2. Inhibition of α-Glucosidase Activity
2.7.3. Inhibition of α-Amylase Activity
2.8. Contaminant Content
2.8.1. Mannitol
2.8.2. Hydroxymethylfurfural
2.8.3. Heavy Metals
2.9. Statistical Analysis
3. Results and Discussion
3.1. Effectivity of PHLE with 15% v/v Glycerol-Water against Maceration with 60% Acetone
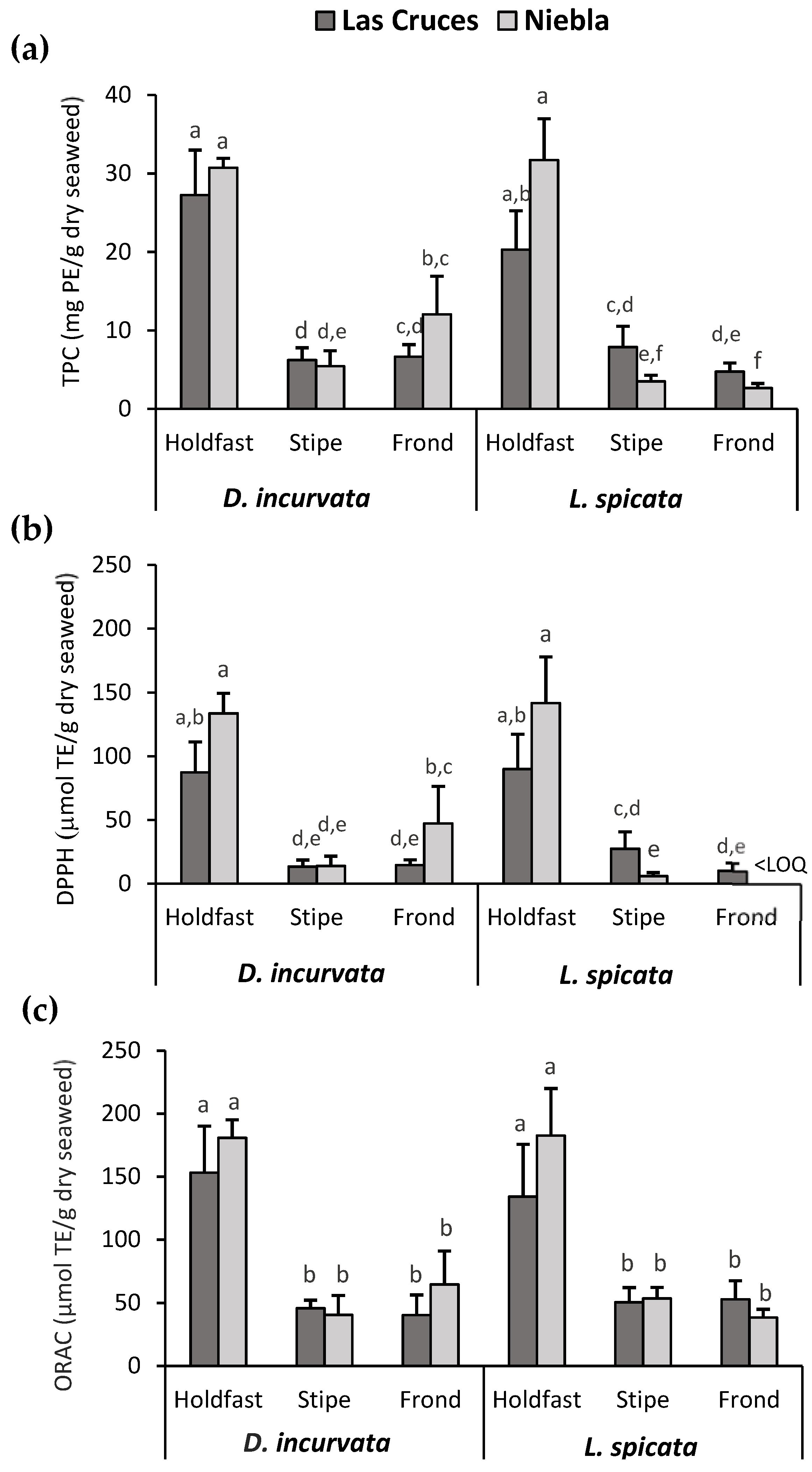
3.2. Effect of the Species, Harvesting Area, and Thallus Part Used on the TPC and AC of D. incurvata and L. spicata
3.3. Extraction Yield, TPC, and AC of the PHLE Extract
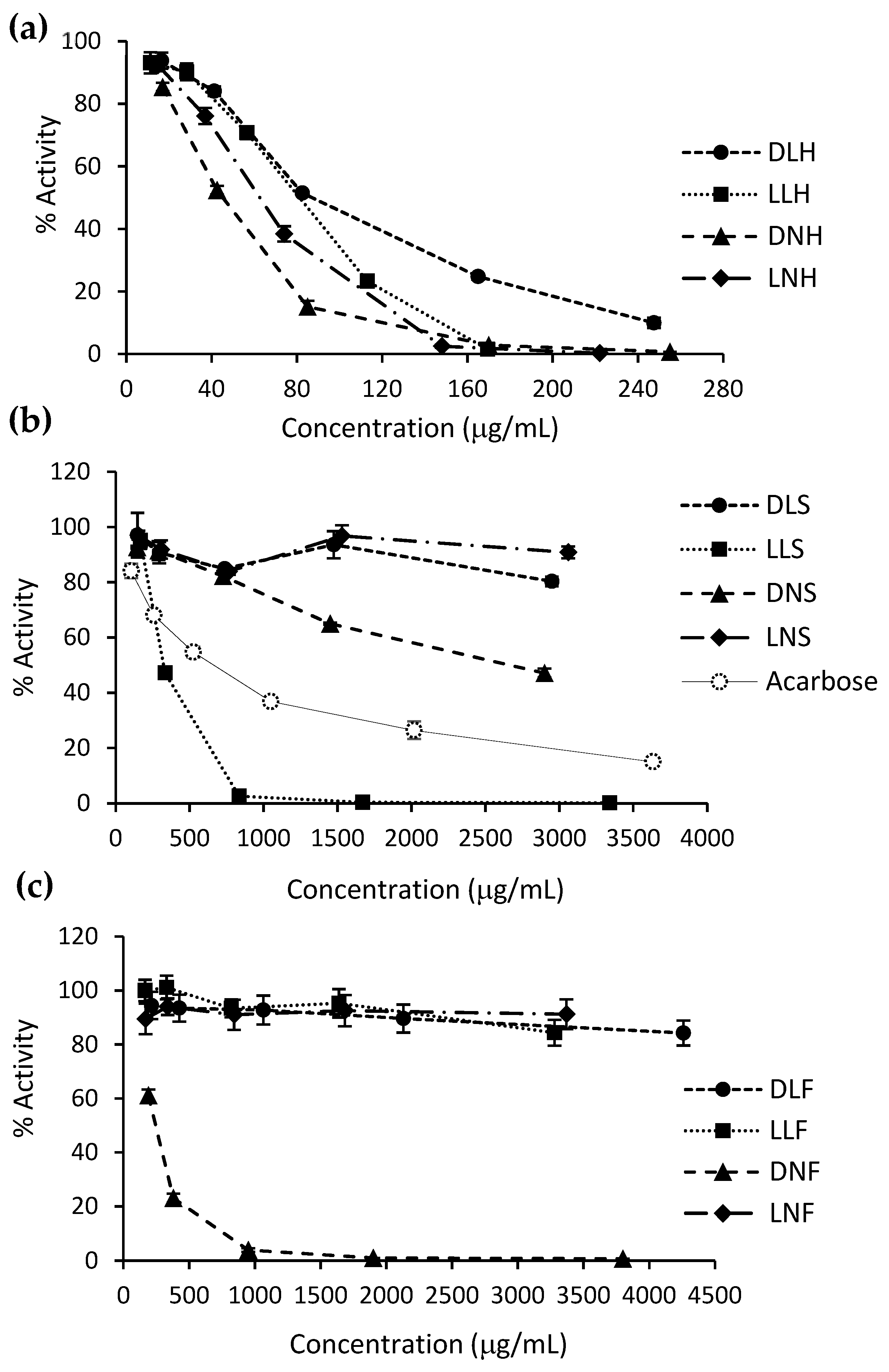
3.4. Carbohydrate-Hydrolyzing Enzymes’ Inhibitory Activity
3.5. Presence of Contaminants: Mannitol, HMF, and Heavy Metals
3.5.1. Mannitol and HMF in PHLE Extracts
3.5.2. Heavy Metals in Dry Seaweeds and PHLE Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peñalver, R.; Lorenzo, J.M.; Ros, G.; Amarowicz, R.; Pateiro, M.; Nieto, G. Seaweeds as a Functional Ingredient for a Healthy Diet. Mar. Drugs 2020, 18, 301. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Ramírez, C.; Pino, M.; Collins, M.B.; Rossen, J.; Pino-Navarro, J.D. Monte Verde: Seaweed, food, medicine, and the peopling of South America. Science 2008, 320, 784–786. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J.; Levine, I.A. Seaweed in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2016; pp. 370–371. [Google Scholar]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic Potential of Marine Brown Algae—a Mini Review. J. Diabetes Res. 2020, 2020, 1230218. [Google Scholar] [CrossRef]
- Rocha, D.H.A.; Seca, A.M.L.; Pinto, D. Seaweed Secondary Metabolites In Vitro and In Vivo Anticancer Activity. Mar. Drugs 2018, 16, 410. [Google Scholar] [CrossRef] [Green Version]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [Green Version]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, N.C.; Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Brown Macroalgae as Valuable Food Ingredients. Antioxidants 2019, 8, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjeewa, K.K.A.; Kim, E.A.; Son, K.T.; Jeon, Y.J. Bioactive properties and potentials cosmeceutical applications of phlorotannins isolated from brown seaweeds: A review. J. Photochem. Photobiol. B Biol. 2016, 162, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jonsdottir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Olafsdottir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Koivikko, R. Brown Algal Phlorotannins: Improving and Applying Chemical Methods; University of Turku: Turku, Finland, 2008. [Google Scholar]
- Schoenwaelder, M.; Clayton, M.N. Physode formation in embryos of Phyllospora comosa and Hormosira banksii (Phaeophyceae). Phycologia 2000, 39, 9. [Google Scholar] [CrossRef]
- Mannino, A.; Micheli, C. Ecological Function of Phenolic Compounds from Mediterranean Fucoid Algae and Seagrasses: An Overview on the Genus Cystoseira sensu lato and Posidonia oceanica (L.) Delile. J. Mar. Sci. Eng. 2020, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.A.; Lee, J.H.; Han, J.S. A phlorotannin constituent of Ecklonia cava alleviates postprandial hyperglycemia in diabetic mice. Pharm. Biol. 2017, 55, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- You, H.N.; Lee, H.A.; Park, M.H.; Lee, J.H.; Han, J.S. Phlorofucofuroeckol A isolated from Ecklonia cava alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2015, 752, 92–96. [Google Scholar] [CrossRef]
- Lee, S.H.; Ko, S.C.; Kang, M.C.; Lee, D.H.; Jeon, Y.J. Octaphlorethol A, a marine algae product, exhibits antidiabetic effects in type 2 diabetic mice by activating AMP-activated protein kinase and upregulating the expression of glucose transporter 4. Food Chem. Toxicol. 2016, 91, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.E.; Islam, N.; Ahn, B.R.; Chowdhury, S.S.; Sohn, H.S.; Jung, H.A.; Choi, J.S. Protein tyrosine phosphatase 1B and a-glucosidase inhibitory Phlorotannins from edible brown algae, Ecklonia stolonifera and Eisenia bicyclis. Biosc. Biotechnol. Biochem. 2011, 75, 1472–1480. [Google Scholar] [CrossRef] [Green Version]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of Phlorotannins Extraction from Fucus vesiculosus and Evaluation of Their Potential to Prevent Metabolic Disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanniou, A.; Serrano-León, E.; Vandanjon, L.; Ibañez, E.; Mendiola, J.A.; Cerantola, S.; Kervarec, N.; LaBarre, S.; Marchal, L.; Stiger-Pouvreau, V. Green improved processes to extract bioactive phenolic compounds from brown macroalga using Sargassum muticum as model. Talanta 2013, 104, 44–52. [Google Scholar] [CrossRef]
- Getachew, A.T.; Jacobsen, C.; Holdt, S.L. Emerging Technologies for the Extraction of Marine Phenolics: Opportunities and Challenges. Mar. Drugs 2020, 18, 389. [Google Scholar] [CrossRef]
- Zakaria, S.M.; Mazlina, S.; Kamal, M. Subcritical Water Extraction of Bioactive Compounds from Plants and Algae: Applications in Pharmaceutical and Food Ingredients. Food Eng. Rev. 2016, 8, 23–34. [Google Scholar] [CrossRef]
- Huamán-Castilla, N.L.; Mariotti-Celis, M.S.; Martínez-Cifuentes, M.; Pérez-Correa, J.R. Glycerol as Alternative Co-Solvent for Water Extraction of Polyphenols from Carmenere Pomace: Hot Pressurized Liquid Extraction and Computational Chemistry Calculations. Biomolecules 2020, 10, 474. [Google Scholar] [CrossRef] [Green Version]
- Huamán-Castilla, N.L.; Martínez-Cifuentes, M.; Camilo, C.; Pedreschi, F.; Mariotti-Celis, M.; Pérez-Correa, J.R. The Impact of Temperature and Ethanol Concentration on the Global Recovery of Specific Polyphenols in an Integrated HPLE/RP Process on Carmenere Pomace Extracts. Molecules 2019, 24, 3145. [Google Scholar] [CrossRef] [Green Version]
- Ozkaynak Kanmaz, E. 5-Hydroxymethylfurfural (HMF) formation during subcritical water extraction. Food Sci. Biotechnol. 2018, 27, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Makris, D.P.; Lalas, S. Glycerol and Glycerol-Based Deep Eutectic Mixtures as Emerging Green Solvents for Polyphenol Extraction: The Evidence So Far. Molecules 2020, 25, 5842. [Google Scholar] [CrossRef] [PubMed]
- Huamán-Castilla, N.L.; Campos, D.; García-Ríos, D.; Parada, J.; Martínez-Cifuentes, M.; Mariotti-Celis, M.S.; Pérez-Correa, J.R. Chemical Properties of Vitis Vinifera Carménère Pomace Extracts Obtained by Hot Pressurized Liquid Extraction, and Their Inhibitory Effect on Type 2 Diabetes Mellitus Related Enzymes. Antioxidants 2021, 10, 472. [Google Scholar] [CrossRef] [PubMed]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef]
- Fraser, C.I.; Velásquez, M.; Nelson, W.A.; Macaya, E.C.; Hay, C.H. The Biogeographic Importance of Buoyancy in Macroalgae: A Case Study of the Southern Bull-Kelp Genus Durvillaea (Phaeophyceae), Including Descriptions of Two New Species. J. Phycol. 2019, 56, 23–36. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Beltrán, J.; Hiriart-Bertrand, L.; Flores, V.; de Reviers, B.; Correa, J.A.; Santelices, B. Identification of Cryptic Species in the Lessonia Nigrescens Complex (Phaeophyceae, Laminariales). J. Phycol. 2012, 48, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Tierney, M.; Smyth, T.; Rai, D.; Soler-Vila, A.; Croft, A.; Brunton, N. Enrichment of polyphenol contents and antioxidant activities of Irish brown macroalgae using food-friendly techniques based on polarity and molecular size. Food Chem. 2013, 139, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and Identification of Phlorotannins from the Brown Alga Sargassum fusiforme (Harvey) Setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.; Prior, R. High-Throughput Assay of Oxygen Radical Absorbance Capacity (ORAC) Using a Multichannel Liquid Handling System Coupled with a Microplate Fluorescence Reader in 96-Well Format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef]
- Pacheco, L.V.; Parada, J.; Pérez-Correa, J.R.; Mariotti-Celis, M.S.; Erpel, F.; Zambrano, A.; Palacios, M. Bioactive Polyphenols from Southern Chile Seaweed as Inhibitors of Enzymes for Starch Digestion. Mar. Drugs 2020, 18, 353. [Google Scholar] [CrossRef] [PubMed]
- ICH. FDA Guidance for Industry: Q2B Validation of Analytical Procedures: Methodology; Food and Drug Administration: Rockville, MD, USA, 1996. [Google Scholar]
- Toker, O.S.; Dogan, M.; Ersoz, N.B.; Yilmaz, M.T. Optimization of the content of 5-hydroxymethylfurfural (HMF) formed in some molasses types: HPLC-DAD analysis to determine effect of different storage time and temperature levels. Ind. Crop. Prod. 2013, 50, 137–144. [Google Scholar] [CrossRef]
- Leege, P.; Millner, P.; Thompson, W.; Watson, M. Test methods for the Examination of Composting and Compost; United States Department of Agriculture and Composting Council Research and Education Foundation: Raleigh, NC, USA, 2002. [Google Scholar]
- SISS. Determinación de Arsénico total por Método espectrofotometría de absorción atómica con generación de hidruros. In Manual de Métodos de Ensayo Para Agua Potable, 2nd ed.; Superitendencia de Servicios Sanitarios: Santiago, Chile, 2007; pp. 101–110. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 2nd ed.; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 2012; p. 724. [Google Scholar]
- Hagerman, A.E. Extraction of tannin from fresh and preserved leaves. J. Chem. Ecol. 1988, 14, 453–461. [Google Scholar] [CrossRef]
- Zhong, Y.; Shahidi, F. Methods for the assessment of antioxidant activity in foods. In Handbook of Antioxidants for Food Preservation; Sahidi, F., Ed.; Woodhead Publishing: Cambridge, UK, 2015. [Google Scholar]
- Schaich, K.; Tian, X.; XIe, J. Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays. J. Funct. Foods 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Vásquez, J. The Brown Seaweeds Fishery in Chile. In Fisheries and Aquaculture in the Modern World; Mikkola, H., Ed.; IntechOpen: London, UK, 2016; pp. 123–141. [Google Scholar]
- Ummat, V.; Tiwari, B.K.; Jaiswal, A.K.; Condon, K.; Garcia-Vaquero, M.; O’Doherty, J.; O’Donnell, C.; Rajauria, G. Optimisation of Ultrasound Frequency, Extraction Time and Solvent for the Recovery of Polyphenols, Phlorotannins and Associated Antioxidant Activity from Brown Seaweeds. Mar. Drugs 2020, 18, 250. [Google Scholar] [CrossRef] [PubMed]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Barba, F.J.; Lorenzo, J.M. Antioxidant Potential of Extracts Obtained from Macro-(Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata) and Micro-Algae (Chlorella vulgaris and Spirulina platensis) Assisted by Ultrasound. Medicines 2018, 5, 33. [Google Scholar] [CrossRef] [Green Version]
- Heffernan, N.; Brunton, N.P.; FitzGerald, R.J.; Smyth, T.J. Profiling of the molecular weight and structural isomer abundance of macroalgae-derived phlorotannins. Mar. Drugs 2015, 13, 509–528. [Google Scholar] [CrossRef]
- Olate-Gallegos, C.; Barriga, A.; Vergara, C.; Fredes, C.; García, P.; Giménez, B.; Robert, P. Identification of Polyphenols from Chilean Brown Seaweeds Extracts by LC-DAD-ESI-MS/MS. J. Aquat. Food Prod. Technol. 2019, 28, 17. [Google Scholar] [CrossRef]
- Beratto-Ramos, A.; Castillo-Felices, R.; Troncoso-León, N.; Agurto-Muñoz, A.; Agurto-Muñoz, C. Selection criteria for high-value biomass: Seasonal and morphological variation of polyphenolic content and antioxidant capacity in two brown macroalgae. J. Appl. Phycol. 2018, 31, 653–664. [Google Scholar] [CrossRef]
- Pinteus, S.; Silva, J.; Alves, C.; Horta, A.; Fino, N.; Rodrigues, A.I.; Mendes, S.; Pedrosa, R. Cytoprotective effect of seaweeds with high antioxidant activity from the Peniche coast (Portugal). Food Chem. 2017, 218, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Múzquiz de la Garza, A.R.; Tapia-Salazar, M.; Maldonado-Muniz, M.; de la Rosa-Millán, J.; Gutiérrez-Uribe, J.A.; Santos-Zea, L.; Barba-Dávila, B.A.; Ricque-Marie, D.; Cruz-Suárez, L.E. Nutraceutical Potential of Five Mexican Brown Seaweeds. Biomed. Res. Int. 2019, 2019, 3795160. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A. Postprandial hyperglycemia and diabetes complications: Is it time to treat? Diabetes 2005, 54, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krentz, A.J.; Bailey, C.J. Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs 2005, 65, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The a-amylase and a-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Kongstad, K.T.; Wiese, S.; Jager, A.K.; Staerk, D. Edible seaweed as future functional food: Identification of a-glucosidase inhibitors by combined use of high-resolution a-glucosidase inhibition profiling and HPLC-HRMS-SPE-NMR. Food Chem. 2016, 203, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Neto, R.T.; Marcal, C.; Queirós, A.S.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Apostolidis, E.; Lee, C.M. In vitro potential of Ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J. Food Sci. 2010, 75, H97–H102. [Google Scholar] [CrossRef]
- Parada, J.; Pérez-Correa, J.R.; Pérez-Jiménez, J. Design of low glycemic response foods using polyphenols from seaweed. J. Funct. Foods 2019, 56, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Daub, C.D.; Mabate, B.; Malgas, S.; Pletschke, B.I. Fucoidan from Ecklonia maxima is a powerful inhibitor of the diabetes-related enzyme, α-glucosidase. Int. J. Biol. Macromol. 2020, 151, 412–420. [Google Scholar] [CrossRef]
- Zaharudin, N.; Staerk, D.; Dragsted, L.O. Inhibition of a-glucosidase activity by selected edible seaweeds and fucoxanthin. Food Chem. 2019, 270, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Plaza, M.; Amigo-Benavente, M.; Del Castillo, M.D.; Ibáñez, E.; Herrero, M. Neoformation of antioxidants in glycation model systems treated under subcritical water extraction conditions. Food Res. Int. 2010, 43, 1123–1129. [Google Scholar] [CrossRef]
- Chades, T.; Scully, S.M.; Ingvadottir, E.M.; Orlygsson, J. Fermentation of Mannitol Extracts from Brown Macro Algae by Thermophilic Clostridia. Front. Microbiol. 2018, 9, 1931. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Ross, A.B.; Anastasakis, K.; Hodgson, E.M.; Gallagher, J.A.; Jones, J.M.; Donnison, I.S. Seasonal variation in the chemical composition of the bioenergy feedstock Laminaria digitata for thermochemical conversion. Bioresour. Technol. 2011, 102, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Hamid, S.S.; Wakayama, M.; Ichihara, K.; Sakurai, K.; Ashino, Y.; Kadowaki, R.; Soga, T.; Tomita, M. Metabolome profiling of various seaweed species discriminates between brown, red, and green algae. Planta 2019, 249, 1921–1947. [Google Scholar] [CrossRef]
- Lenhart, A.; Chey, W.D. A Systematic Review of the Effects of Polyols on Gastrointestinal Health and Irritable Bowel Syndrome. Adv. Nutr. 2017, 8, 587–596. [Google Scholar] [CrossRef]
- André, P.; Villain, F. Free radical scavenging properties of mannitol and its role as a constituent of hyaluronic acid fillers: A literature review. Int. J. Cosmet. Sci. 2017, 39, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Abraham, K.; Gurtler, R.; Berg, K.; Heinemeyer, G.; Lampen, A.; Appel, K.E. Toxicology and risk assessment of 5-Hydroxymethylfurfural in food. Mol. Nutr. Food Res. 2011, 55, 667–678. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Sunwoo, I.Y.; Jeong, G.T.; Kim, S.K. Detoxification of Hydrolysates of the Red Seaweed Gelidium amansii for Improved Bioethanol Production. Appl. Biochem. Biotechnol. 2019, 188, 977–990. [Google Scholar] [CrossRef]
- Yang, C.F.; Huang, C.R. Isolation of 5-hydroxymethylfurfural biotransforming bacteria to produce 2,5-furan dicarboxylic acid in algal acid hydrolysate. J. Biosci. Bioeng. 2018, 125, 407–412. [Google Scholar] [CrossRef]
- Duangkaew, P.; Inoue, S.; Aki, T.; Nakashimada, Y.; Okamura, Y.; Tajima, T.; Matsumura, Y. Quantitative In Situ Mass Spectrometry Analysis of Mannitol Decomposition Products under Hydrothermal Conditions. Energy Fuels 2017, 31, 10866–10873. [Google Scholar] [CrossRef]
- Ou, J.; Zheng, J.; Huang, J.; Ho, C.T.; Ou, S. Interaction of Acrylamide, Acrolein, and 5-Hydroxymethylfurfural with Amino Acids and DNA. J. Agric. Food Chem. 2020, 68, 5039–5048. [Google Scholar] [CrossRef] [PubMed]
- Paz, S.; Rubio, C.; Frías, I.; Gutiérrez, A.J.; González-Weller, D.; Martín, V.; Revert, C.; Hardisson, A. Toxic metals (Al, Cd, Pb and Hg) in the most consumed edible seaweeds in Europe. Chemosphere 2019, 218, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Almela, C.; Clemente, M.J.; Vélez, D.; Montoro, R. Total arsenic, inorganic arsenic, lead and cadmium contents in edible seaweed sold in Spain. Food Chem. Toxicol. 2006, 44, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Biancarosa, I.; Belghit, I.; Bruckner, C.G.; Liland, N.S.; Waagbo, R.; Amlund, H.; Heesch, S.; Lock, E.J. Chemical characterization of 21 species of marine macroalgae common in Norwegian waters: Benefits of and limitations to their potential use in food and feed. J. Sci. Food Agric. 2018, 98, 2035–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besada, V.; Andrade, J.M.; Schultze, F.; González, J.J. Heavy metals in edible seaweeds commercialised for human consumption. J. Mar. Syst. 2009, 75, 305–313. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M.; Jeitner, C.; Gray, M.; Shukla, T.; Shukla, S.; Burke, S. Kelp as a bioindicator: Does it matter which part of 5 m long plant is used for metal analysis? Environ. Monit. Assess. 2007, 128, 311–321. [Google Scholar] [CrossRef]
- Pan, Y.; Wernberg, T.; de Bettignies, T.; Holmer, M.; Li, K.; Wu, J.; Lin, F.; Yu, Y.; Xu, J.; Zhou, C.; et al. Screening of seaweeds in the East China Sea as potential bio-monitors of heavy metals. Environ. Sci. Pollut. Res. 2018, 25, 16640–16651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maehre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Díaz, O.; Tapia, Y.; Muñoz, O.; Montoro, R.; Vélez, D.; Almela, C. Total and inorganic arsenic concentrations in different species of economically important algae harvested from coastal zones of Chile. Food Chem. Toxicol. 2012, 50, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Camurati, J.R.; Salomone, V.N. Arsenic in edible macroalgae: An integrated approach. J. Toxicol. Environ. Health Part B 2020, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.A.; Cassiano, J.; Veiga, P.; Rubal, M.; Sousa-Pinto, I. Fucus spiralis as monitoring tool of metal contamination in the northwest coast of Portugal under the European Water Framework Directives. Environ. Monit. Assess. 2014, 186, 5447–5460. [Google Scholar] [CrossRef]
- Sáez, C.A.; Lobos, M.G.; Macaya, E.C.; Oliva, D.; Quiroz, W.; Brown, M.T. Variation in patterns of metal accumulation in thallus parts of Lessonia trabeculata (Laminariales; Phaeophyceae): Implications for biomonitoring. PLoS ONE 2012, 7, e50170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Lee, J.Y.; Seo, J.E.; Jeong, J.Y.; Jung, K.K.; Yoon, H.J.; Park, K.S. Lead, cadmium and mercury levels in the 2010 Korean diet. Food Addit. Contam. Part B 2012, 5, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Nagpal, A.K. Contamination of vegetables with heavy metals across the globe: Hampering food security goal. J. Food Sci. Technol. 2020, 57, 391–403. [Google Scholar] [CrossRef] [PubMed]
- EC. Commission Regulation No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs; European Comission: Brussels, Belgium, 2006; pp. 6–9. [Google Scholar]
- Squadrone, S.; Brizio, P.; Battuello, M.; Nurra, N.; Sartor, R.M.; Riva, A.; Staiti, M.; Benedetto, A.; Pessani, D.; Abete, M.C. Trace metal occurrence in Mediterranean seaweeds. Environ. Sci. Pollut. Res. 2018, 25, 9708–9721. [Google Scholar] [CrossRef]
- Filippini, T.; Tancredi, S.; Malagoli, C.; Cilloni, S.; Malavolti, M.; Violi, F.; Vescovi, L.; Bargellini, A.; Vinceti, M. Aluminum and tin: Food contamination and dietary intake in an Italian population. J. Trace Elem. Med. Biol. 2019, 52, 293–301. [Google Scholar] [CrossRef]
- Corrias, F.; Atzei, A.; Addis, P.; Secci, M.; Russo, M.; Angioni, A. Integrated environmental evaluation of heavy metals and metalloids bioaccumulation in invertebrates and seaweeds from different marine coastal areas of sardinia, mediterranean sea. Environ. Pollut. 2020, 266, 115048. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.; Cardoso, C.; Ripol, A.; Varela, J.; Quental-Ferreira, H.; Pousao-Ferreira, P.; Ventura, M.S.; Delgado, I.M.; Coelho, I.; Castanheira, I.; et al. Composition and bioaccessibility of elements in green seaweeds from fish pond aquaculture. Food Res. Int. 2018, 105, 271–277. [Google Scholar] [CrossRef]
- Desideri, D.; Cantaluppi, C.; Ceccotto, F.; Meli, M.A.; Roselli, C.; Feduzi, L. Essential and toxic elements in seaweeds for human consumption. J. Toxicol. Environ. Health Part A 2016, 79, 112–122. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Area | Alga Part | Group ID |
|---|---|---|---|
| D. incurvata | Las Cruces | Holdfast | DLH |
| Stipe | DLS | ||
| Frond | DLF | ||
| Niebla | Holdfast | DNH | |
| Stipe | DNS | ||
| Frond | DNF | ||
| L. spicata | Las Cruces | Holdfast | LLH |
| Stipe | LLS | ||
| Frond | LLF | ||
| Niebla | Holdfast | LNH | |
| Stipe | LNS | ||
| Frond | LNF |
| Compound | Concentration Range (mg/L) | Slope | Interception | Coefficient of Determination (R2) | LOD (mg/L) | LOQ (mg/L) | RSD |
|---|---|---|---|---|---|---|---|
| Mannitol | 100–1000 | 0.0031 | −0.0399 | 0.9986 | 42 | 126 | 0.039 |
| HMF | 0.025–0.250 | 0.0042 | −0.0332 | 0.9983 | 0.012 | 0.036 | 0.015 |
| Group ID | Extraction Yield | TPC | DPPH RSA | ORAC | |
|---|---|---|---|---|---|
| (% Dry Seaweed) | (mg PE/g Dry Extract) | (g PE/100 g Dry Seaweed) | (μmol TE/g Dry Extract) | (μmol TE/g Dry Extract) | |
| DLH | 36.8 ± 6.6 a | 75 ± 14 a,b | 2.7 ± 0.6 a | 238 ± 50 a,b | 416 ± 73 a |
| DLS | 34.7 ± 5.0 a,b | 17.9 ± 3.4 d | 0.6 ± 0.2 d | 39 ± 14 d,e | 135 ± 32 b |
| DLF | 33.6 ± 6.7 a,b | 21.1 ± 9.2 c,d | 0.7 ± 0.2 c,d | 47 ± 23 d,e | 126 ± 36 b |
| DNH | 34.7 ± 4.8 a,b | 90 ± 14 a | 3.1 ± 0.1 a | 392 ± 71 a,b | 530 ± 100 a |
| DNS | 31 ± 11 a,b | 20 ± 11 c,d | 0.5 ± 0.2 d,e | 49 ± 25 c-e | 144 ± 63 b |
| DNF | 31.5 ± 7.1 a,b | 41 ± 21 b,c | 1.2 ± 0.5 b,c | 160 ± 120 b,c | 220 ± 110 b |
| LLH | 23.8 ± 5.2 a,b | 85 ± 12 a | 2.0 ± 0.5 a,b | 377 ± 92 a,b | 559 ± 96 a |
| LLS | 34.4 ± 7.0 a,b | 24 ± 10 c,d | 0.8 ± 0.3 c,d | 82 ± 45 c,d | 152 ± 45 b |
| LLF | 30.3 ± 3.0 a,b | 16.3 ± 5.4 d | 0.5 ± 0.1 d,e | 35 ± 24 d,e | 181 ± 62 b |
| LNH | 24.9 ± 2.6 a,b | 129 ± 30 a | 3.2 ± 0.5 a | 580 ± 180 a | 740 ± 190 a |
| LNS | 27.0 ± 8.0 a,b | 13.6 ± 3.9 d | 0.3 ± 0.1 e,f | 22 ± 8 e | 212 ± 69 b |
| LNF | 22.9 ± 3.7 b | 11.7 ± 1.8 d | 0.3 ± 0.1 f | <LOQ | 174 ± 51 b |
| Group ID | IC50 (μg/mL) α-Glucosidase |
|---|---|
| DLH | 87.1 ± 0.8 c |
| DNH | 45.2 ± 1.6a |
| DNS | 2700 ± 100g |
| DNF | 245.1 ± 5.3d |
| LLH | 81.2 ± 0.9c |
| LLS | 324.1 ± 6.5e |
| LNH | 62.6 ± 2.4b |
| Acarbose | 659.5 ± 36.7f |
| Group ID | Mannitol | |
|---|---|---|
| (% Dry Seaweed) | (mg/g Dry Extract) | |
| DLH | 15.3 ± 2.7 | 423 ± 85 |
| DLS | 16.4 ± 2.9 | 473 ± 59 |
| DLF | 10.0 ± 1.8 | 310 ± 120 |
| DNH | 10.4 ± 0.9 | 303 ± 46 |
| DNS | 9.3 ± 2.6 | 360 ± 240 |
| DNF | 7.7 ± 1.4 | 248 ± 39 |
| LLH | 5.9 ± 1.5 | 260 ± 110 |
| LLS | 7.2 ± 1.4 | 213 ± 43 |
| LLF | 5.0 ± 1.4 | 168 ± 46 |
| LNH | 3.6 ± 0.8 | 147 ± 42 |
| LNS | 6.6 ± 1.0 | 257 ± 56 |
| LNF | 5.4 ± 1.5 | 237 ± 46 |
| Group ID | Total As | Cd | Pb | Hg | Sn | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| S | E | S | E | S | E | S | E | S | E | |
| DLH | 23.06 | 49.09 | 1.38 | 0.27 | <1.00 | 0.55 | ND | n.a. | <2.00 | ND |
| DLS | 10.83 | 20.11 | 1.70 | 0.22 | <1.00 | 0.43 | ND | n.a. | <2.00 | ND |
| DLF | 17.80 | 27.96 | 0.84 | 0.22 | <1.00 | 0.44 | ND | n.a. | <2.00 | ND |
| DNH | 48.62 | 90.88 | 2.15 | 3.71 | <1.00 | 0.28 | ND | n.a. | ND | n.a. |
| DNS | 8.29 | 14.88 | 1.07 | 0.20 | <1.00 | 0.41 | ND | n.a. | ND | n.a. |
| DNF | 11.30 | 26.58 | 0.86 | 0.23 | <1.00 | 0.47 | ND | n.a. | ND | n.a. |
| LLH | 43.09 | 202.19 | 3.13 | 6.13 | <1.00 | 0.46 | ND | n.a. | <2.00 | ND |
| LLS | 17.81 | 39.53 | 2.33 | 0.28 | <1.00 | 0.56 | ND | n.a. | <2.00 | ND |
| LLF | 26.28 | 55.40 | 4.51 | 0.31 | <1.00 | 0.62 | ND | n.a. | ND | n.a. |
| LNH | 84.08 | 258.47 | 4.35 | 5.38 | <1.00 | 0.81 | ND | n.a. | <2.00 | ND |
| LNS | 29.78 | 61.81 | 1.87 | 0.34 | <1.00 | 0.69 | ND | n.a. | ND | n.a. |
| LNF | 29.27 | 35.68 | 1.57 | 0.38 | <1.00 | 0.75 | ND | n.a. | <2.00 | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erpel, F.; Mariotti-Celis, M.S.; Parada, J.; Pedreschi, F.; Pérez-Correa, J.R. Pressurized Hot Liquid Extraction with 15% v/v Glycerol-Water as An Effective Environment-Friendly Process to Obtain Durvillaea incurvata and Lessonia spicata Phlorotannin Extracts with Antioxidant and Antihyperglycemic Potential. Antioxidants 2021, 10, 1105. https://doi.org/10.3390/antiox10071105
Erpel F, Mariotti-Celis MS, Parada J, Pedreschi F, Pérez-Correa JR. Pressurized Hot Liquid Extraction with 15% v/v Glycerol-Water as An Effective Environment-Friendly Process to Obtain Durvillaea incurvata and Lessonia spicata Phlorotannin Extracts with Antioxidant and Antihyperglycemic Potential. Antioxidants. 2021; 10(7):1105. https://doi.org/10.3390/antiox10071105
Chicago/Turabian StyleErpel, Fernanda, María Salomé Mariotti-Celis, Javier Parada, Franco Pedreschi, and José Ricardo Pérez-Correa. 2021. "Pressurized Hot Liquid Extraction with 15% v/v Glycerol-Water as An Effective Environment-Friendly Process to Obtain Durvillaea incurvata and Lessonia spicata Phlorotannin Extracts with Antioxidant and Antihyperglycemic Potential" Antioxidants 10, no. 7: 1105. https://doi.org/10.3390/antiox10071105
APA StyleErpel, F., Mariotti-Celis, M. S., Parada, J., Pedreschi, F., & Pérez-Correa, J. R. (2021). Pressurized Hot Liquid Extraction with 15% v/v Glycerol-Water as An Effective Environment-Friendly Process to Obtain Durvillaea incurvata and Lessonia spicata Phlorotannin Extracts with Antioxidant and Antihyperglycemic Potential. Antioxidants, 10(7), 1105. https://doi.org/10.3390/antiox10071105