
Impact of Ascorbate—Glutathione Cycle Components on the Effectiveness of Embryogenesis Induction in Isolated Microspore Cultures of Barley and Triticale

 ,
,  , ,
, ,  , ,
, ,
Abstract
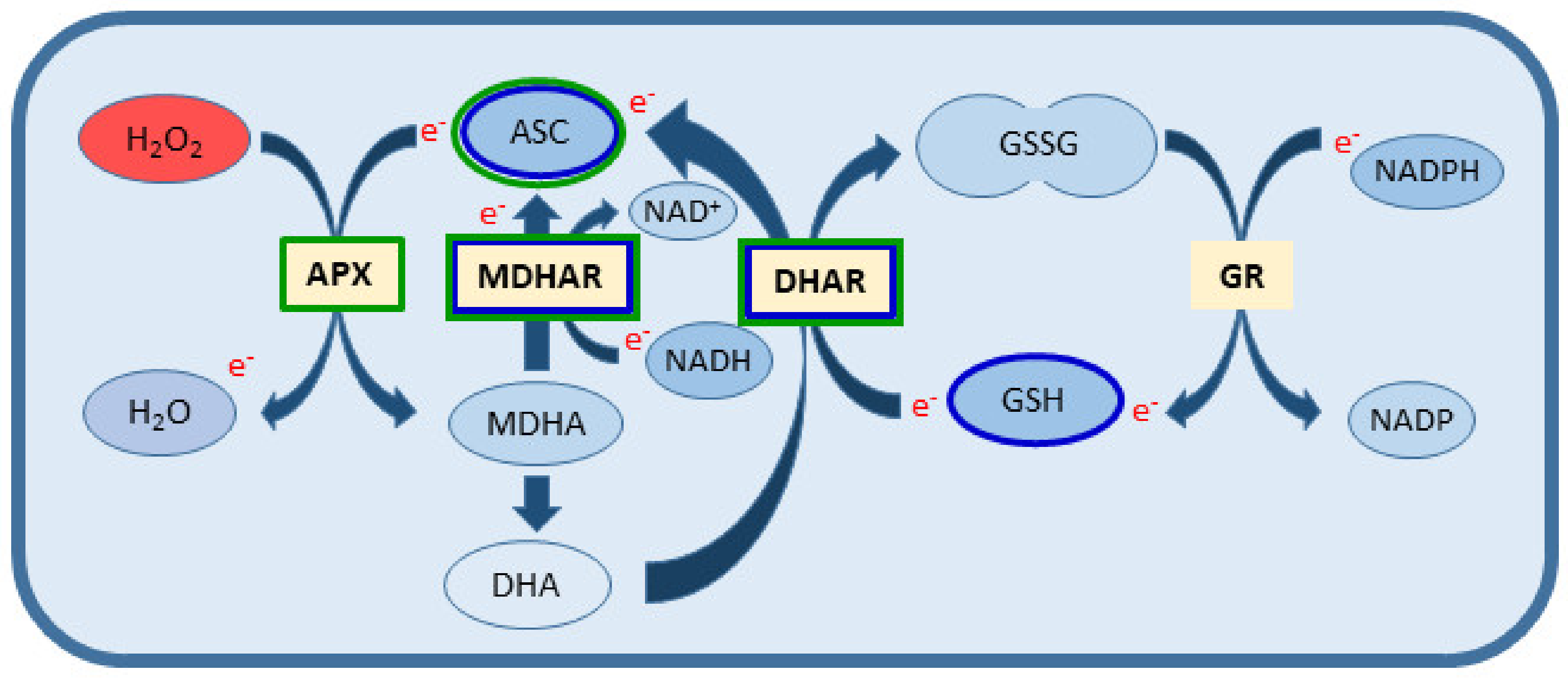
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. ME Inducing Treatment, Microspore Isolation and In Vitro Culture Conditions for Antioxidant System Analysis
2.3. Parameters Describing Productivity, Viability and Embryogenic Potential of Microspores
- (i)
- Microspore yield—mean number of isolated microspores obtained per one spike of a donor plant calculated using a Neubauer chamber.
- (ii)
- Microspore viability—percentage of fully viable microspores in the whole population of isolated cells, assessed based on its reaction with fluorescein diacetate (FDA; 0.01%; λEx = 465 nm, λEm = 515 nm, green fluorescence; according to Heslop-Harrison and Heslop-Harrison [25].
- (iii)
- Effectiveness of ME induction—the number of developed embryo-like structures (ELS) observed after 6 weeks of in vitro culture, calculated per one spike of a donor plant [ELS/spike].
2.4. In Situ Histochemical Detection of Superoxide Anions and Hydrogen Peroxide in Microspores
2.5. Low Molecular Weight Antioxidant Activity by DPPH Assay
2.6. Sample Preparation for Other Assays
2.7. Sample Preparation for Ascorbate and Glutathione Assays
2.8. Reduced and Oxidized Glutathione Measurements
2.9. Reduced and Oxidized Ascorbate Measurements
2.10. Sample Preparation and Enzyme Activity Assays
2.11. Statistical Analysis
3. Results
3.1. The Effect of Tiller Pre-Treatments on the Effectiveness of ME Induction
3.2. In Situ Histochemical Detection of Superoxide Anions and Hydrogen Peroxide in Microspores
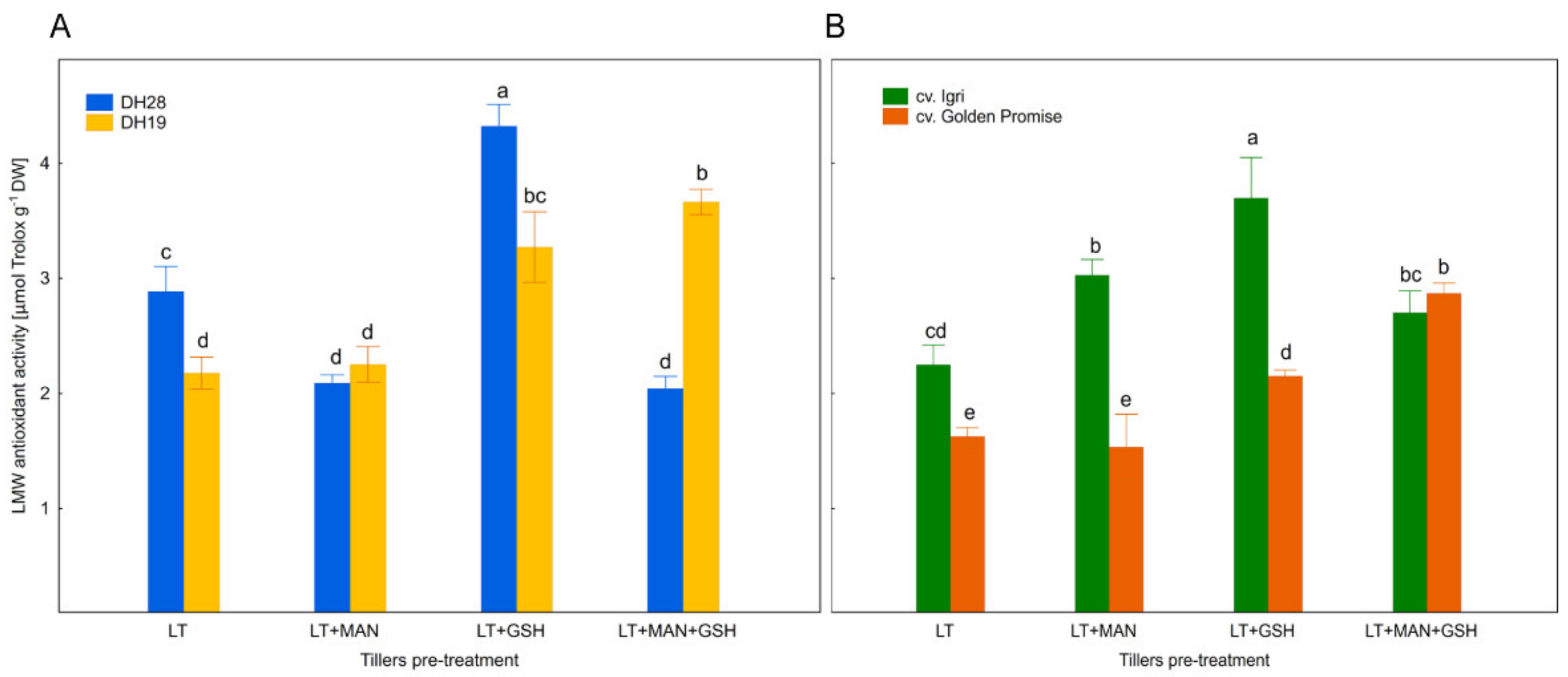
3.3. Total Activity of Low Molecular Weight Antioxidants in Microspores after Various ME-Inducing Pre-Treatments of Tillers
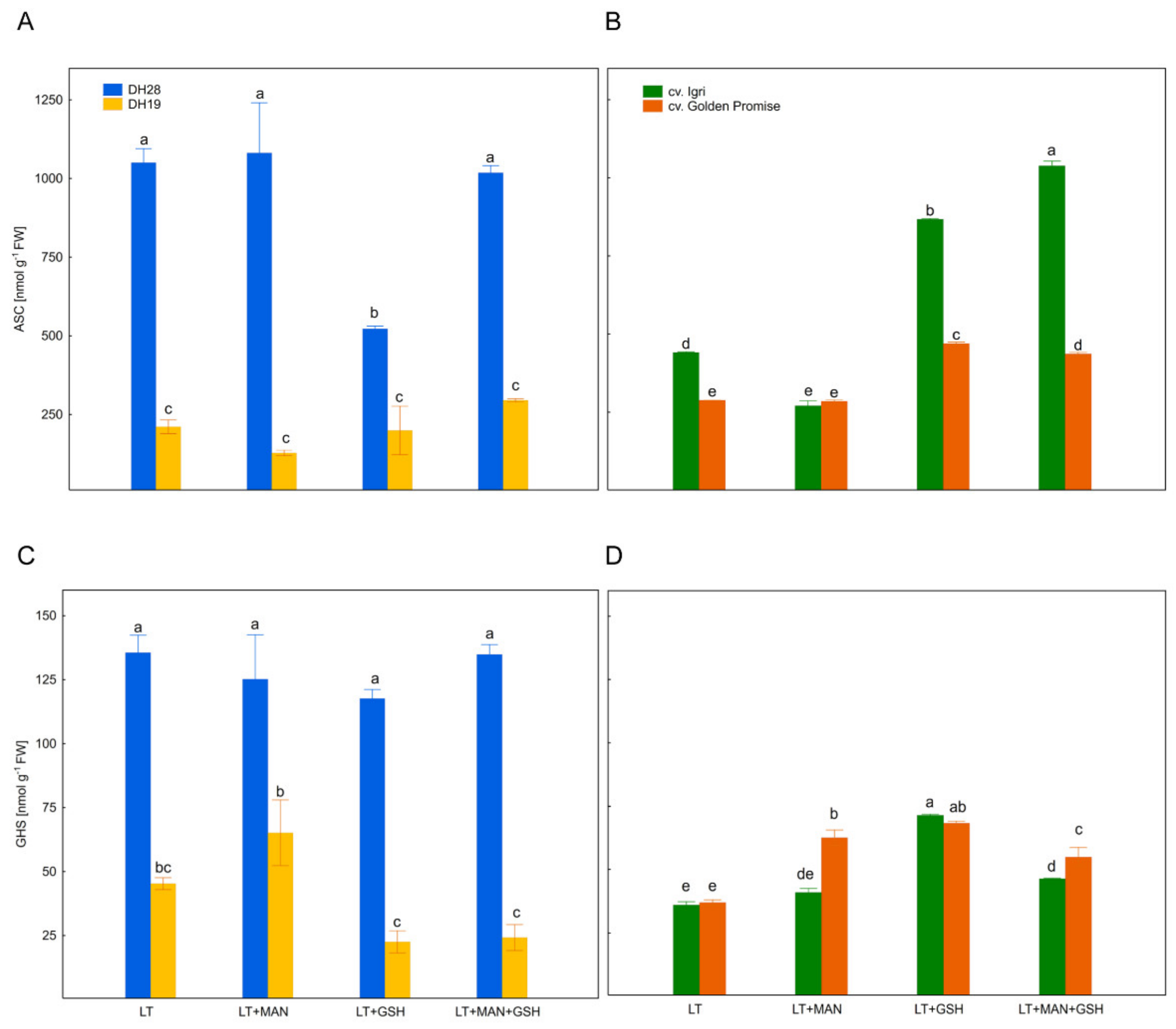
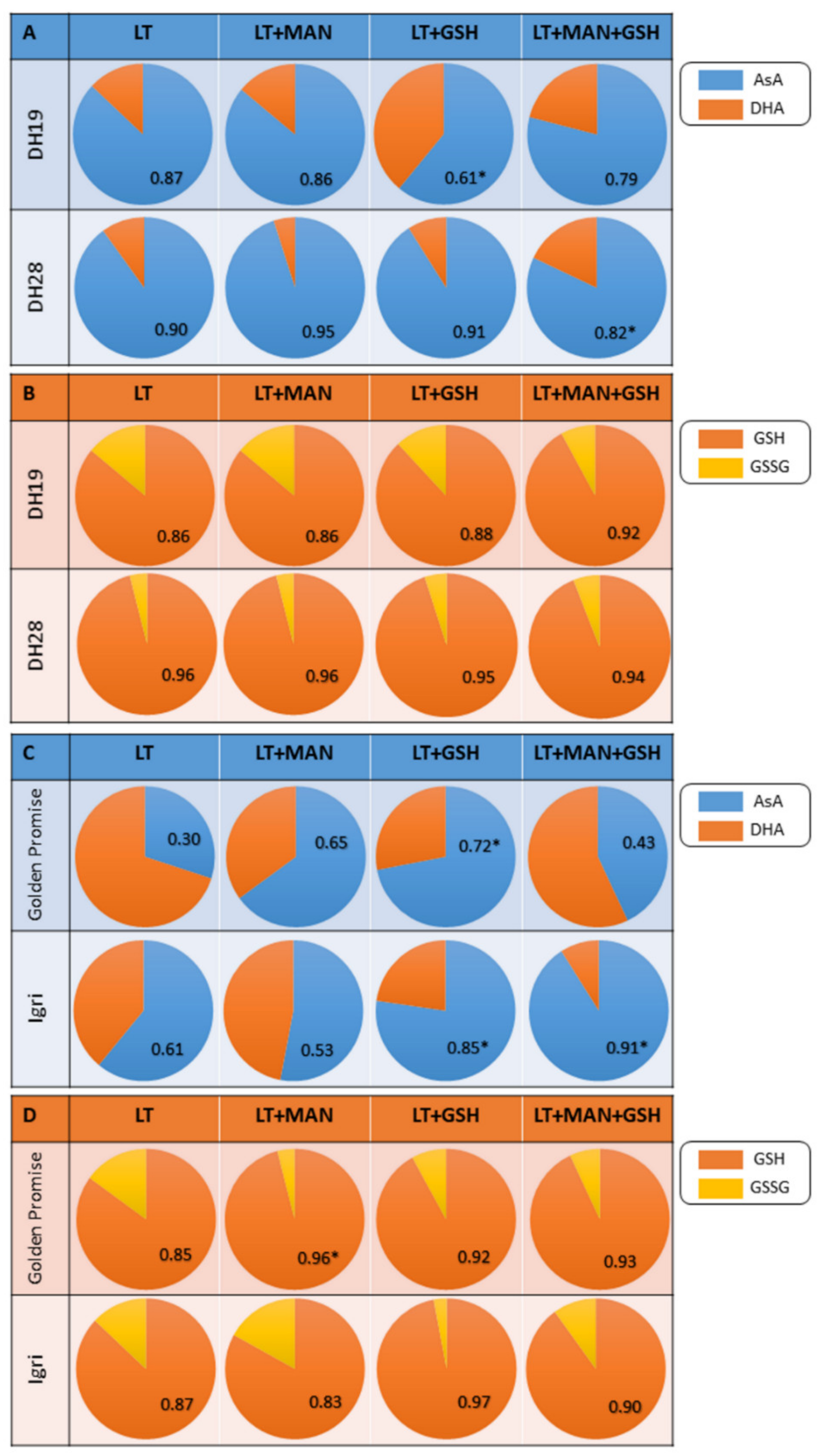
3.4. Glutathione and Ascorbate Levels and Their Reduction State in Triticale and Barley Microspores after Various ME-Inducing Pre-Treatments of Tillers
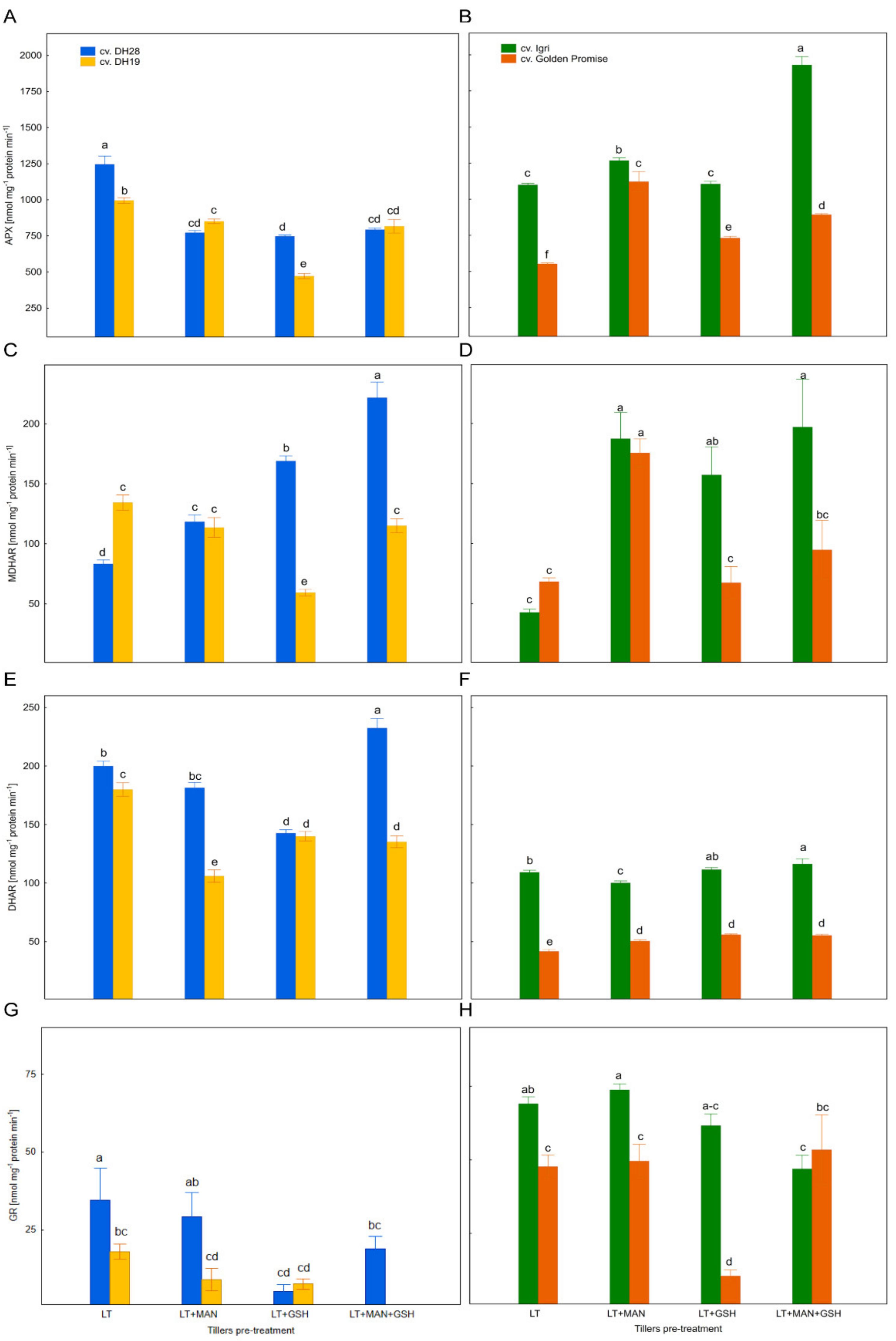
3.5. The Activity of Enzymes of the Glutathione–Ascorbate Cycle in Microspores after Various ME-Inducing Pre-Treatments of Tillers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Janků, M.; Luhová, L.; Petřivalský, M. On the Origin and Fate of Reactive Oxygen Species in Plant Cell Compartments. Antioxidants 2019, 8, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Żur, I.; Dubas, E.; Krzewska, M.; Zieliński, K.; Fodor, J.; Janowiak, F. Glutathione provides antioxidative defence and promotes microspore-derived embryo development in isolated microspore cultures of triticale (×Triticosecale Wittm.). Plant Cell Rep. 2019, 38, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Żur, I.; Dubas, E.; Krzewska, M.; Kopeć, P.; Nowicka, A.; Surówka, E.; Gawrońska, K.; Gołębiowska, G.; Juzoń, K.; Malaga, S. Triticale and barley microspore embryogenesis induction requires both reactive oxygen species generation and efficient system of antioxidative defence. Plant Cell Tissue Organ Cult. 2021, 145, 347–366. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid: A potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef]
- Zhang, Y. Biological role of ascorbate in plants. In Ascorbic Acid in Plants; SpringerBriefs in Plant Science; Springer: New York, NY, USA, 2013; pp. 7–33. [Google Scholar] [CrossRef]
- Xiao, M.; Li, Z.; Zhu, L.; Wang, J.; Zhang, B.; Zheng, F.; Zhao, B.; Zhang, H.; Wang, Y.; Zhang, Z. The multiple roles of ascorbate in the abiotic stress response of plants: Antioxidant, cofactor, and regulator. Front. Plant Sci. 2021, 12, 592. [Google Scholar] [CrossRef]
- Elhiti, M.; Yang, C.; Belmonte, M.F.; Gulden, R.H.; Stasolla, C. Transcriptional changes of antioxidant responses, hormone signalling and developmental processes evoked by the Brassica napus SHOOTMERISTEMLESS during in vitro embryogenesis. Plant Physiol. Biochem. 2012, 58, 297–311. [Google Scholar] [CrossRef]
- Asif, M.; Eudes, F.; Goyal, A.; Amundsen, E.; Randhawa, H.; Spaner, D. Organelle antioxidants improve microspore embryogenesis in wheat and triticale. Vitro Cell. Dev. Biol. 2013, 49, 489–497. [Google Scholar] [CrossRef]
- Hoseini, M.; Ghadimzadeh, M.; Ahmadi, B.; da Silva, J.A.T. Effects of ascorbic acid, alpha-tocopherol, and glutathione on microspore embryogenesis in Brassica napus L. Vitro Cell. Dev. Biol. 2014, 50, 26–35. [Google Scholar] [CrossRef]
- Yerzhebayeva, R.S.; Abekova, A.M.; Ainebekova, B.A.; Urazaliyev, K.R.; Bazylova, T.A.; Daniyarova, A.K.; Bersimbayeva, G.K. Influence of different concentrations of ascorbic and gibberellic acids and pH of medium on embryogenesis and regeneration in anther culture of spring triticale. Cytol. Genet. 2017, 51, 448–454. [Google Scholar] [CrossRef]
- Cistue, L.; Ziauddin, A.; Simion, E.; Kasha, K.J. Effects of culture conditions on isolated microspore response of barley cultivar Igri. Plant Cell Tissue Organ Cult. 1995, 42, 163–169. [Google Scholar] [CrossRef]
- Coronado, M.J.; Hensel, G.; Broeders, S.; Otto, I.; Kumlehn, J. Immature pollen-derived doubled haploid formation in barley cv. Golden Promise as a tool for transgene recombination. Acta Physiol. Plant 2005, 27, 591–599. [Google Scholar] [CrossRef]
- Lippmann, R.; Friedel, S.; Mock, H.-P.; Kumlehn, J. The low molecular weight fraction of compounds released from immature embryogenesis. Front. Plant Sci. 2015, 6, 498. [Google Scholar] [CrossRef] [Green Version]
- Żur, I.; Dubas, E.; Krzewska, M.; Waligórski, P.; Dziurka, M.; Janowiak, F. Hormonal requirements for effective induction of microspore embryogenesis in triticale (×Triticosecale Wittm.) anther cultures. Plant Cell Rep. 2015, 34, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Krzewska, M.; Gołebiowska-Pikania, G.; Dubas, E.; Gawin, M.; Żur, I. Identification of proteins related to microspore embryogenesis responsiveness in anther cultures of winter triticale (×Triticosecale Wittm.). Euphytica 2017, 213, 192. [Google Scholar] [CrossRef] [Green Version]
- Wędzony, M. Protocol for doubled haploid production in hexaploid Triticale (×Triticosecale Wittm.) by crosses with maize. In Doubled Haploid Production in Crop Plants. A Manual; Maluszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 135–140. [Google Scholar] [CrossRef]
- Zhuang, J.; Xu, J. Increasing differentiation frequencies in wheat pollen callus. In Cell and Tissue Culture Techniques for Cereal Crop Improvement; Hu, H., Vega, M., Eds.; Science Press: Beijing, China, 1983; p. 431. [Google Scholar]
- Pauk, J.; Mihály, R.; Monostori, T.; Puolimatka, M. Protocol of triticale (×Triticosecale Wittmack) microspore culture. In Doubled Haploid Production in Crop Plants. A Manual; Maluszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 129–134. [Google Scholar] [CrossRef]
- Kumlehn, J.; Serazetdinova, L.; Hensel, G.; Becker, D.; Loerz, H. Genetic transformation of barley (Hordeum vulgare L.) via infection of androgenetic pollen cultures with Agrobacterium tumefaciens. Plant Biotechnol. J. 2006, 4, 251–261. [Google Scholar] [CrossRef]
- Heslopharrison, J.; Heslopha, Y. Evaluation of pollen viability by enzymatically induced fluorescence—Intracellular hydrolysis of fluorescein diacetate. Stain Technol. 1970, 45, 115–120. [Google Scholar] [CrossRef]
- Żur, I.; Dubas, E.; Krzewska, M.; Janowiak, F.; Hura, K.; Pociecha, E.; Bączek-Kwinta, R.; Płażek, A. Antioxidant activity and ROS tolerance in triticale (×Triticosecale Wittm.) anthers affect the efficiency of microspore embryogenesis. Plant Cell Tissue Organ Cult. 2014, 119, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Wohlgemuth, H.; Mittelstrass, K.; Kschieschan, S.; Bender, J.; Weigel, H.-J.; Overmyer, K.; Kangasjärvi, J.; Sandermann, H.; Langebartels, C. Activation of an oxidative burst is a general feature of sensitive plants exposed to the air pollutant ozone. Plant Cell Environ. 2002, 25, 717–726. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free-radical method to evaluate antioxidant activity. Food Sci. Technol.-Lebensm.-Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Laskoś, K.; Pisulewska, E.; Waligórski, P.; Janowiak, F.; Janeczko, A.; Sadura, I.; Polaszczyk, S.; Czyczyło-Mysza, I.M. Herbal additives substantially modify antioxidant properties and tocopherol content of cold-pressed oils. Antioxidants 2021, 10, 781. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.; Rowell, J.; Walker, D. Measurements of the ascorbate content of spinach leaf protoplasts and chloroplasts during illumination. Planta 1983, 157, 239–244. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Klapheck, S.; Zimmer, I.; Cosse, H. Scavenging of hydrogen peroxide in the endosperm of Ricinus communis by ascorbate peroxidase. Plant Cell Physiol. 1990, 31, 1005–1013. [Google Scholar] [CrossRef]
- Hossain, M.A.; Nakano, Y.; Asada, K. Monodehydroascorbate reductase in spinach chloroplasts and its participation in regeneration of ascorbate for scavenging hydrogen peroxide. Plant Cell Physiol. 1984, 25, 385–395. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Thomas, W.; Forster, B.; Gertsson, B. Doubled haploids in breeding. In Doubled Haploid Production in Crop Plants. A Manual; Maluszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 337–349. [Google Scholar] [CrossRef]
- Forster, B.; Thomas, W. Doubled haploids in genetic mapping and genomics. In Doubled Haploid Production in Crop Plants. A Manual; Maluszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 367–390. [Google Scholar] [CrossRef]
- Szarejko, I.; Forster, B.P. Doubled haploidy and induced mutation. Euphytica 2007, 158, 359–370. [Google Scholar] [CrossRef]
- Humphreys, D.G.; Knox, R.E. Doubled haploid breeding in cereals. In Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland; Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2015; Volume 1, pp. 241–290. [Google Scholar]
- Touraev, A.; Vicente, O.; HeberleBors, E. Initiation of microspore embryogenesis by stress. Trends Plant Sci. 1997, 2, 297–302. [Google Scholar] [CrossRef]
- Shariatpanahi, M.E.; Belogradova, K.; Hessamvaziri, L.; Heberle-Bors, E.; Touraev, A. Efficient embryogenesis and regeneration in freshly isolated and cultured wheat (Triticum aestivum L.) microspores without stress pretreatment. Plant Cell Rep. 2006, 25, 1294–1299. [Google Scholar] [CrossRef]
- Zoriniants, S.; Tashpulatov, A.S.; Heberle-Bors, E.; Touraev, A. The role of stress in the induction of haploid microspore embryogenesis. In Haploids in Crop Improvement II; Don Palmer, C., Keller, W.A., Kasha, K.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 35–52. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Casalongue, C.A.; Simontacchi, M.; Marquez-Garcia, B.; Foyer, C.H. Interactions between hormone and redox signalling pathways in the control of growth and cross tolerance to stress. Environ. Exp. Bot. 2013, 94, 73–88. [Google Scholar] [CrossRef]
- Hosp, J.; de Faria Maraschin, S.; Touraev, A.; Boutilier, K. Functional genomics of microspore embryogenesis. Euphytica 2007, 158, 275–285. [Google Scholar] [CrossRef]
- Jacquard, C.; Mazeyrat-Gourbeyre, F.; Devaux, P.; Boutilier, K.; Baillieul, F.; Clement, C. Microspore embryogenesis in barley: Anther pre-treatment stimulates plant defence gene expression. Planta 2009, 229, 393–402. [Google Scholar] [CrossRef]
- Zieliński, K.; Krzewska, M.; Żur, I.; Juzoń, K.; Kopeć, P.; Nowicka, A.; Moravcikova, J.; Skrzypek, E.; Dubas, E. The effect of glutathione and mannitol on androgenesis in anther and isolated microspore cultures of rye (Secale cereale L.). Plant Cell Tissue Organ Cult. 2020, 140, 577–592. [Google Scholar] [CrossRef] [Green Version]
- Mignolet-Spruyt, L.; Xu, E.; Idänheimo, N.; Hoeberichts, F.A.; Mühlenbock, P.; Brosché, M.; Van Breusegem, F.; Kangasjärvi, J. Spreading the news: Subcellular and organellar reactive oxygen species production and signalling. J. Exp. Bot. 2016, 67, 3831–3844. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Van Aken, O.; Schwarzländer, M.; Belt, K.; Millar, A.H. The Roles of Mitochondrial Reactive Oxygen Species in Cellular Signaling and Stress Response in Plants. Plant Physiol. 2016, 171, 1551–1559. [Google Scholar] [CrossRef] [Green Version]
- Lennicke, C.; Rahn, J.; Lichtenfels, R.; Wessjohann, L.A.; Seliger, B. Hydrogen peroxide—Production, fate and role in redox signaling of tumor cells. Cell Commun. Signal. 2015, 13, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Potters, G.; De Gara, L.; Asard, H.; Horemans, N. Ascorbate and glutathione: Guardians of the cell cycle, partners in crime? Plant Physiol. Biochem. 2002, 40, 537–548. [Google Scholar] [CrossRef]
- Noctor, G. Metabolic signalling in defence and stress: The central roles of soluble redox couples. Plant Cell Environ. 2006, 29, 409–425. [Google Scholar] [CrossRef] [PubMed]
- Bilska, K.; Wojciechowska, N.; Alipour, S.; Kalemba, E.M. Ascorbic acid: The little-known antioxidant in woody plants. Antioxidants 2019, 8, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Yu, J.; Gomez, F.; Fernandez, L.; McIntosh, L.; Foyer, C.H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot. 2006, 57, 1621–1631. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Tambussi, E.A.; Diego, F.; Foyer, C.H. Control of ascorbic acid synthesis and accumulation and glutathione by the incident light red/far red ratio in Phaseolus vulgaris leaves. Febs Lett. 2009, 583, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Colombo, G.; Giustarini, D.; Milzani, A. Protein S-glutathionylation: A regulatory device from bacteria to humans. Trends Biochem. Sci. 2009, 34, 85–96. [Google Scholar] [CrossRef]
- Vrinten, P.L.; Nakamura, T.; Kasha, K.J. Characterization of cDNAs expressed in the early stages of microspore embryogenesis in barley (Hordeum vulgare) L. Plant Mol. Biol. 1999, 41, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Maraschin, S.d.F.; Caspers, M.; Potokina, E.; Wuelfert, F.; Graner, A.; Spaink, H.P.; Wang, M. CDNA array analysis of stress-induced gene expression in barley androgenesis. Physiol. Plant. 2006, 127, 535–550. [Google Scholar] [CrossRef]
- Munoz-Amatriain, M.; Svensson, J.T.; Castillo, A.-M.; Cistue, L.; Close, T.J.; Valles, M.-P. Transcriptome analysis of barley anthers: Effect of mannitol treatment on microspore embryogenesis. Physiol. Plant. 2006, 127, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Belanger, S.; Marchand, S.; Jacques, P.-E.; Meyers, B.; Belzile, F. Differential expression profiling of microspores during the early stages of isolated microspore culture using the responsive barley cultivar Gobernadora. G3-Genes Genom. Genet. 2018, 8, 1603–1614. [Google Scholar] [CrossRef] [Green Version]
- Żur, I.; Dubas, E.; Krzewska, M.; Sanchez-Diaz, R.A.; Castillo, A.M.; Valles, M.P. Changes in gene expression patterns associated with microspore embryogenesis in hexaploid triticale (×Triticosecale Wittm.). Plant Cell Tissue Organ Cult. 2014, 116, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Schnaubelt, D.; Queval, G.; Dong, Y.; Diaz-Vivancos, P.; Makgopa, M.E.; Howell, G.; De Simone, A.; Bai, J.; Hannah, M.A.; Foyer, C.H. Low glutathione regulates gene expression and the redox potentials of the nucleus and cytosol in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 266–279. [Google Scholar] [CrossRef]
- Stasolla, C.; Yeung, E.C. Recent advances in conifer somatic embryogenesis: Improving somatic embryo quality. Plant Cell Tissue Organ Cult. 2003, 74, 15–35. [Google Scholar] [CrossRef]
- Stasolla, C.; Belmonte, M.F.; van Zyl, L.; Craig, D.L.; Liu, W.B.; Yeung, E.C.; Sederoff, R.R. The effect of reduced glutathione on morphology and gene expression of white spruce (Picea glauca) somatic embryos. J. Exp. Bot. 2004, 55, 695–709. [Google Scholar] [CrossRef]
- Belmonte, M.F.; Ambrose, S.J.; Ross, A.R.S.; Abrams, S.R.; Stasolla, C. Improved development of microspore-derived embryo cultures of Brassica napus cv Topaz following changes in glutathione metabolism. Physiol. Plant. 2006, 127, 690–700. [Google Scholar] [CrossRef]
- Cistue, L.; Romagosa, I.; Batlle, F.; Echavarri, B. Improvements in the production of doubled haploids in durum wheat (Triticum turgidum L.) through isolated microspore culture. Plant Cell Rep. 2009, 28, 727–735. [Google Scholar] [CrossRef] [Green Version]
- De Gara, L.; de Pinto, M.C.; Moliterni, V.M.C.; D’Egidio, M.G. Redox regulation and storage processes during maturation in kernels of Triticum durum. J. Exp. Bot. 2003, 54, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, M.E.; Macey, J.; Yeung, E.C.; Stasolla, C. The effect of osmoticum on ascorbate and glutathione metabolism during white spruce (Picea glauca) somatic embryo development. Plant Physiol. Biochem. 2005, 43, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Stasolla, C. Glutathione redox regulation of in vitro embryogenesis. Plant Physiol. Biochem. 2010, 48, 319–327. [Google Scholar] [CrossRef]
- Gémes, K.; Kim, Y.J.; Park, K.Y.; Moschou, P.N.; Andronis, E.; Valassaki, C.; Roussis, A.; Roubelakis-Angelakis, K.A. An NADPH-oxidase/polyamine oxidase feedback loop controls oxidative burst under salinity. Plant Physiol. 2016, 172, 1418–1431. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Shang, Z.; Wu, J.; Jiang, X.; Moschou, P.N.; Sun, W.; Roubelakis-Angelakis, K.A.; Zhang, S. Spermidine oxidase-derived H2O2 regulates pollen plasma membrane hyperpolarization-activated Ca2+-permeable channels and pollen tube growth. Plant J. 2010, 63, 1042–1053. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Material | Treatment | Microspore Yield | Viability | ME Induction |
|---|---|---|---|---|
| DH19 | LT (control) | 111.0 ± 11 ns | 14.3 ±1 cd | 0 |
| LT+MAN | 103.5 ± 11 | 6.9 ± 1 d | 0 | |
| LT+GSH | 88.5 ± 6 | 17.8 ± 2 bc | 0 | |
| LT+MAN+GSH | 116.5 ± 6 | 5.9 ± 1 d | 0.7 ± 0.7 | |
| mean | 104.8 ± 5 | 11.7 ± 1 | 0.2 ± 0.2 | |
| DH28 | LT (control) | 99.2 ± 6 ns | 30.3 ± 3 a | 17 ± 6 ns |
| LT+MAN | 87.1 ± 10 | 17.4 ± 5 bc | 32 ± 11 | |
| LT+GSH | 86.3 ± 67 | 29.4 ± 2 a | 12 ± 5 | |
| LT+MAN+GSH | 81.2 ± 5 | 24.0 ± 3 ab | 31 ± 18 | |
| mean | 88.4 ± 3 | 26.1 ±2 | 22± 5 | |
| cv. Golden Promise | LT (control) | 29 ± 4 ns | 51 ± 3 b | 2 ± 0.9 c |
| LT+MAN | 27 ± 5 | 57 ± 3 ab | 15 ± 7 c | |
| LT+GSH | 26 ± 5 | 53 ± 3 b | 2 ± 1 c | |
| LT+MAN+GSH | 25 ± 4 | 58 ± 3 ab | 12 ± 8 c | |
| mean | 26.1 ± 2 | 54.7 ± 1 | 8 ± 2 | |
| cv. Igri | LT (control) | 21 ± 6 ns | 65 ± 2 a | 414 ± 121 bc |
| LT+MAN | 25 ± 7 | 64 ± 2 a | 1069 ± 407 b | |
| LT+GSH | 28 ± 8 | 57 ± 3 ab | 543 ± 296 bc | |
| LT+MAN+GSH | 32 ± 9 | 59 ± 2 ab | 2047 ± 622 a | |
| mean | 30.4 ± 5 | 61.6 ± 1 | 1073 ± 277 |
| Parameter | Variable | MS | F | p |
|---|---|---|---|---|
| ASC | (1) Plant genotype | 118 E4 | 179.5 | *** |
| (2) Tillers pre-treatment | 147 E3 | 22.4 | *** | |
| (1) × (2) | 164 E3 | 24.9 | *** | |
| GSH | (1) Plant genotype | 199 E2 | 172.0 | *** |
| (2) Tillers pre-treatment | 252 | 2.2 | ns | |
| (1) × (2) | 826 | 7.1 | *** | |
| LMW antioxidants | (1) Plant genotype | 3.564 | 22.13 | *** |
| (2) Tillers pre-treatment | 5.635 | 34.99 | *** | |
| (1) × (2) | 2.002 | 12.43 | *** | |
| APX | (1) Plant genotype | 887 E3 | 216.6 | *** |
| (2) Tillers pre-treatment | 294 E3 | 70.2 | *** | |
| (1) × (2) | 279 E3 | 66.4 | *** | |
| MDHAR | (1) Plant genotype | 107 E2 | 61.9 | ** |
| (2) Tillers pre-treatment | 216 E2 | 125.2 | ** | |
| (1) × (2) | 9349 | 54.3 | ** | |
| DHAR | (1) Plant genotype | 468 E2 | 576.4 | *** |
| (2) Tillers pre-treatment | 2113 | 26.0 | *** | |
| (1) × (2) | 2181 | 26.8 | *** | |
| GR | (1) Plant genotype | 7468 | 91.21 | *** |
| (2) Tillers pre-treatment | 1298 | 15.85 | *** | |
| (1) × (2) | 398 | 4.86 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żur, I.; Kopeć, P.; Surówka, E.; Dubas, E.; Krzewska, M.; Nowicka, A.; Janowiak, F.; Juzoń, K.; Janas, A.; Barna, B.; et al. Impact of Ascorbate—Glutathione Cycle Components on the Effectiveness of Embryogenesis Induction in Isolated Microspore Cultures of Barley and Triticale. Antioxidants 2021, 10, 1254. https://doi.org/10.3390/antiox10081254
Żur I, Kopeć P, Surówka E, Dubas E, Krzewska M, Nowicka A, Janowiak F, Juzoń K, Janas A, Barna B, et al. Impact of Ascorbate—Glutathione Cycle Components on the Effectiveness of Embryogenesis Induction in Isolated Microspore Cultures of Barley and Triticale. Antioxidants. 2021; 10(8):1254. https://doi.org/10.3390/antiox10081254
Chicago/Turabian StyleŻur, Iwona, Przemysław Kopeć, Ewa Surówka, Ewa Dubas, Monika Krzewska, Anna Nowicka, Franciszek Janowiak, Katarzyna Juzoń, Agnieszka Janas, Balázs Barna, and et al. 2021. "Impact of Ascorbate—Glutathione Cycle Components on the Effectiveness of Embryogenesis Induction in Isolated Microspore Cultures of Barley and Triticale" Antioxidants 10, no. 8: 1254. https://doi.org/10.3390/antiox10081254
APA StyleŻur, I., Kopeć, P., Surówka, E., Dubas, E., Krzewska, M., Nowicka, A., Janowiak, F., Juzoń, K., Janas, A., Barna, B., & Fodor, J. (2021). Impact of Ascorbate—Glutathione Cycle Components on the Effectiveness of Embryogenesis Induction in Isolated Microspore Cultures of Barley and Triticale. Antioxidants, 10(8), 1254. https://doi.org/10.3390/antiox10081254