
Malaria Pigment Hemozoin Impairs GM-CSF Receptor Expression and Function by 4-Hydroxynonenal

 ,
, 
Abstract
:
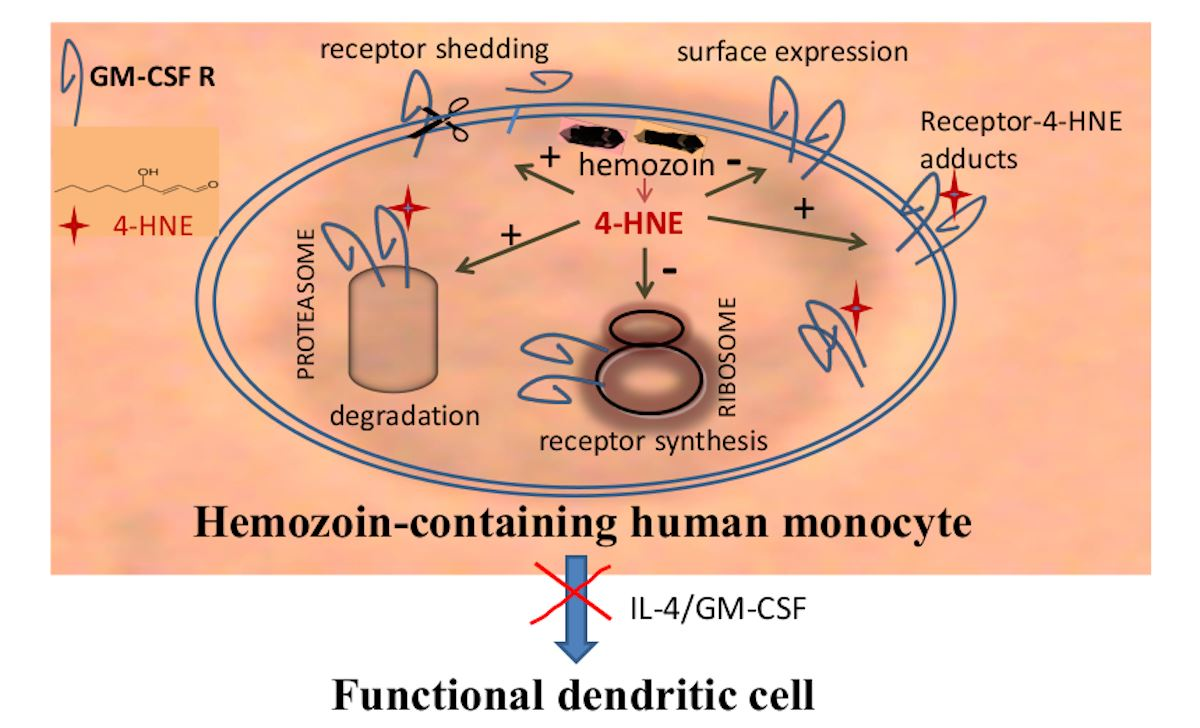{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Reagents
2.2. Culturing of Plasmodium falciparum (Pf) and Isolation of HZ
2.3. Opsonisation of HZ, Latex Beads, and RBC
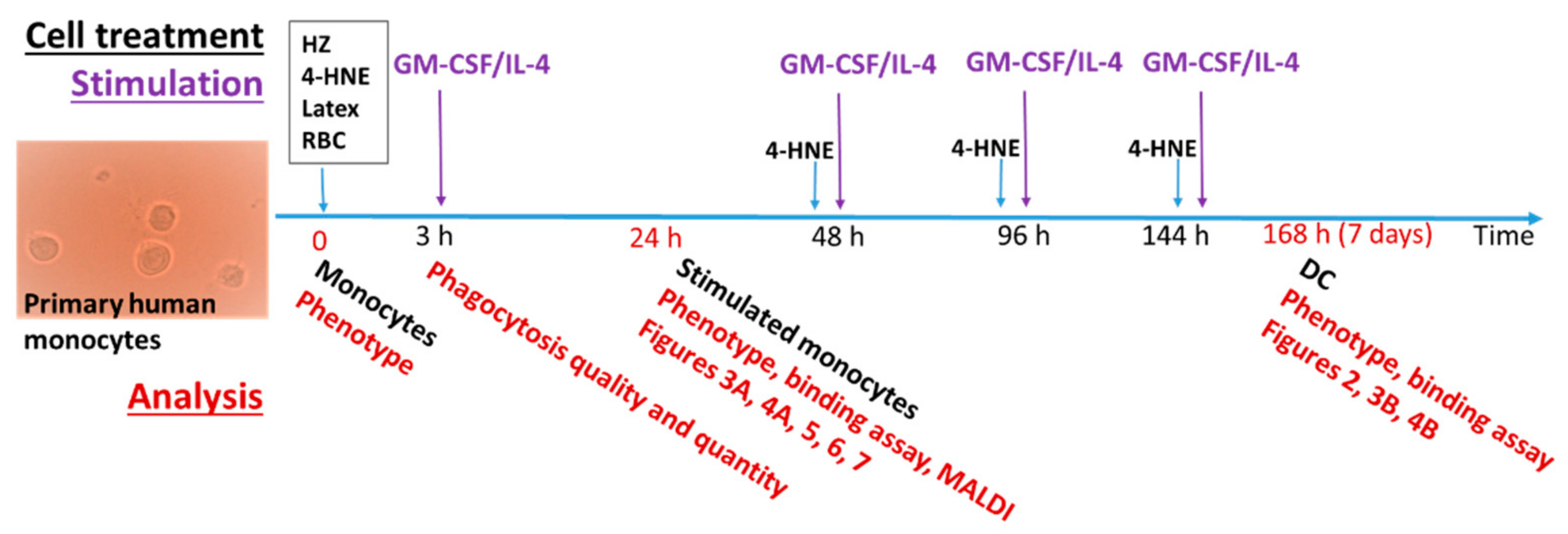
2.4. Isolation of Monocytes, Phagocytosis of Opsonised HZ, RBC, Latex Beads, and Treatment with 4-HNE
2.5. DC Differentiation
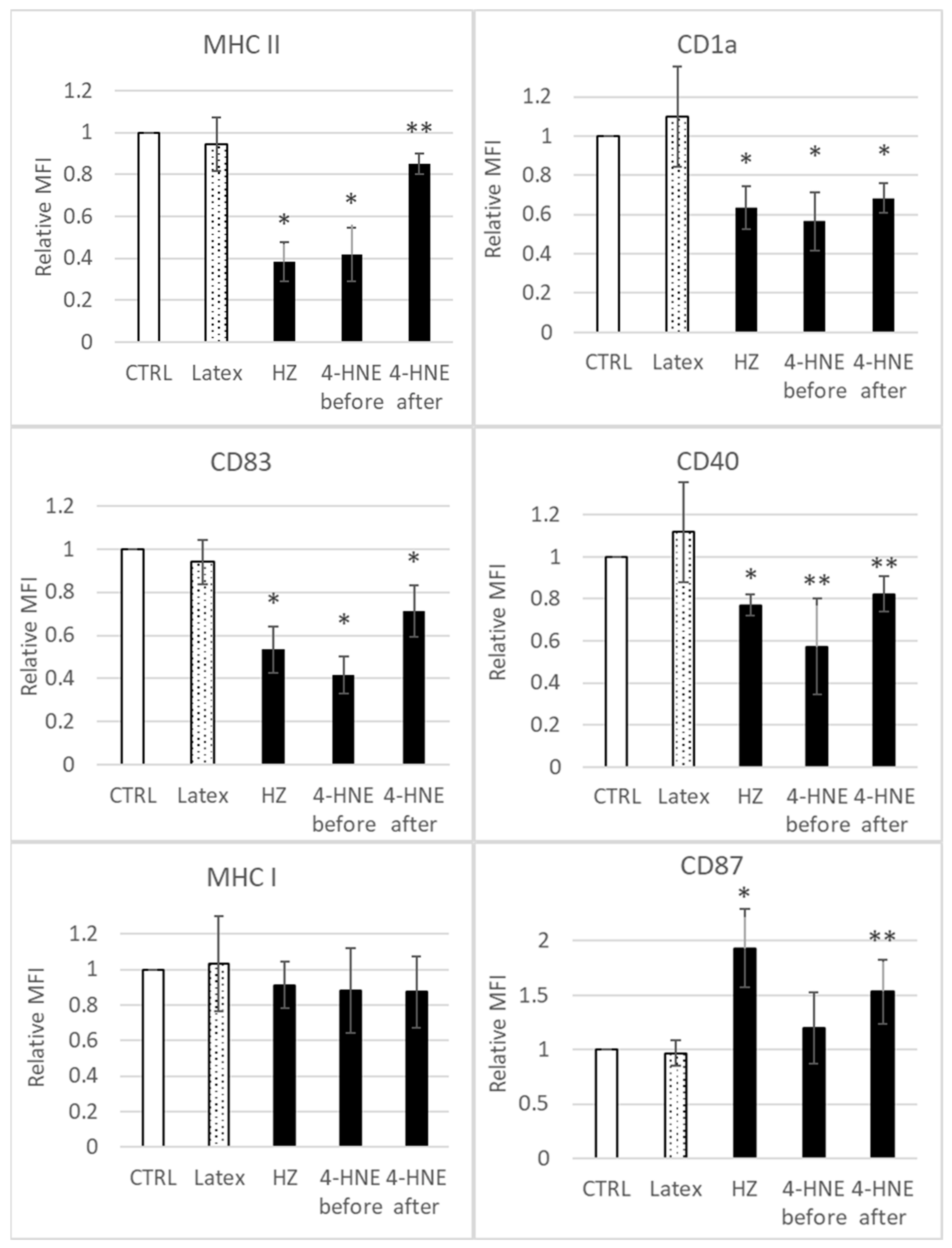
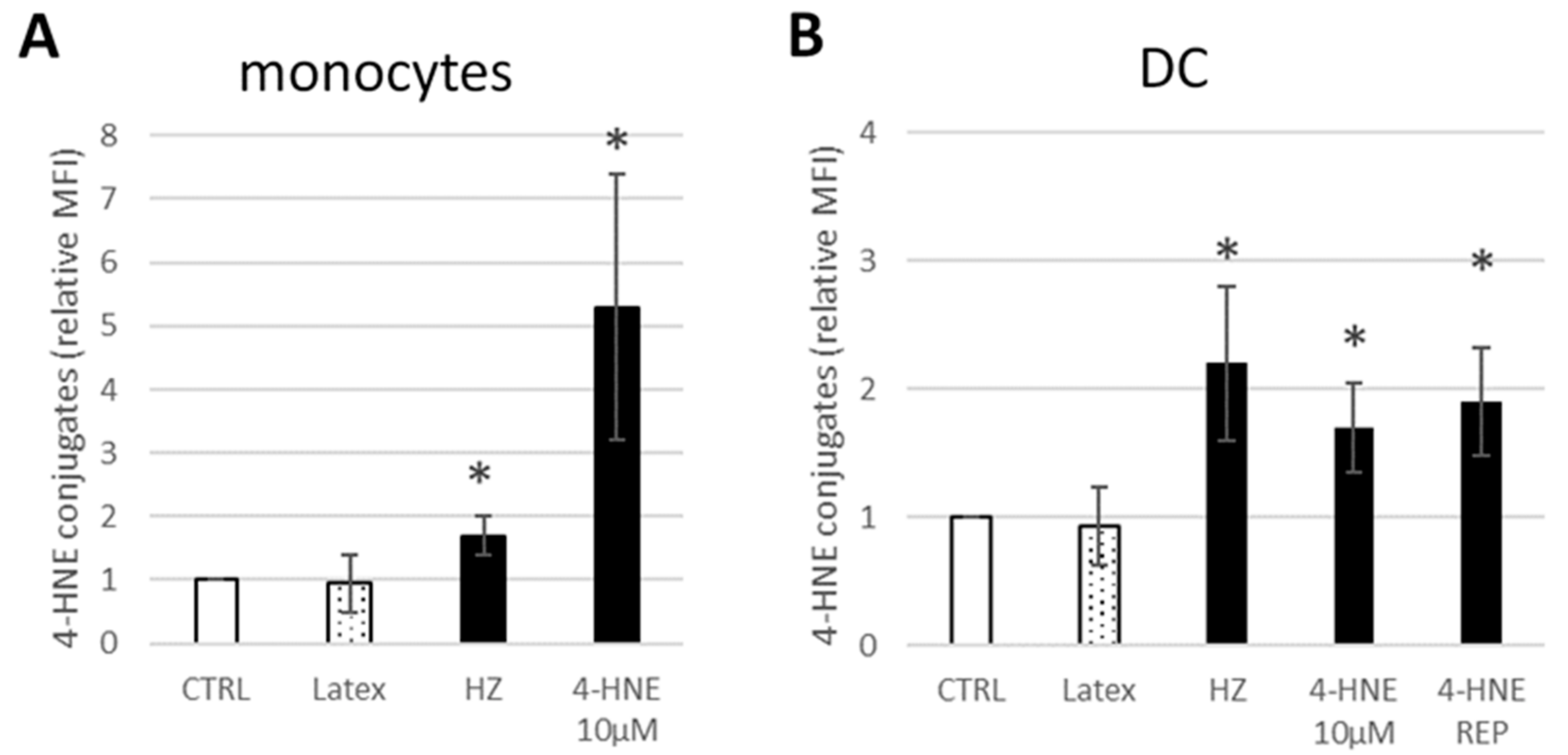
2.6. Flow Cytometry Analysis of Cell Phenotype, CD116 Expression, and Surface 4-HNE-Protein Conjugates
2.7. Fluorescent GM-CSF Binding to Cells
2.8. Whole Cell Lysate Analysis for CD116 Expression: Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blotting
2.9. Identification of 4-HNE Binding Sites in GM-CSF by Mass Spectrometry
2.10. Apoptosis Analysis
2.11. Statistical Analysis
3. Results
3.1. HZ and 4-HNE Impaired the Differentiation of Monocytes to DC
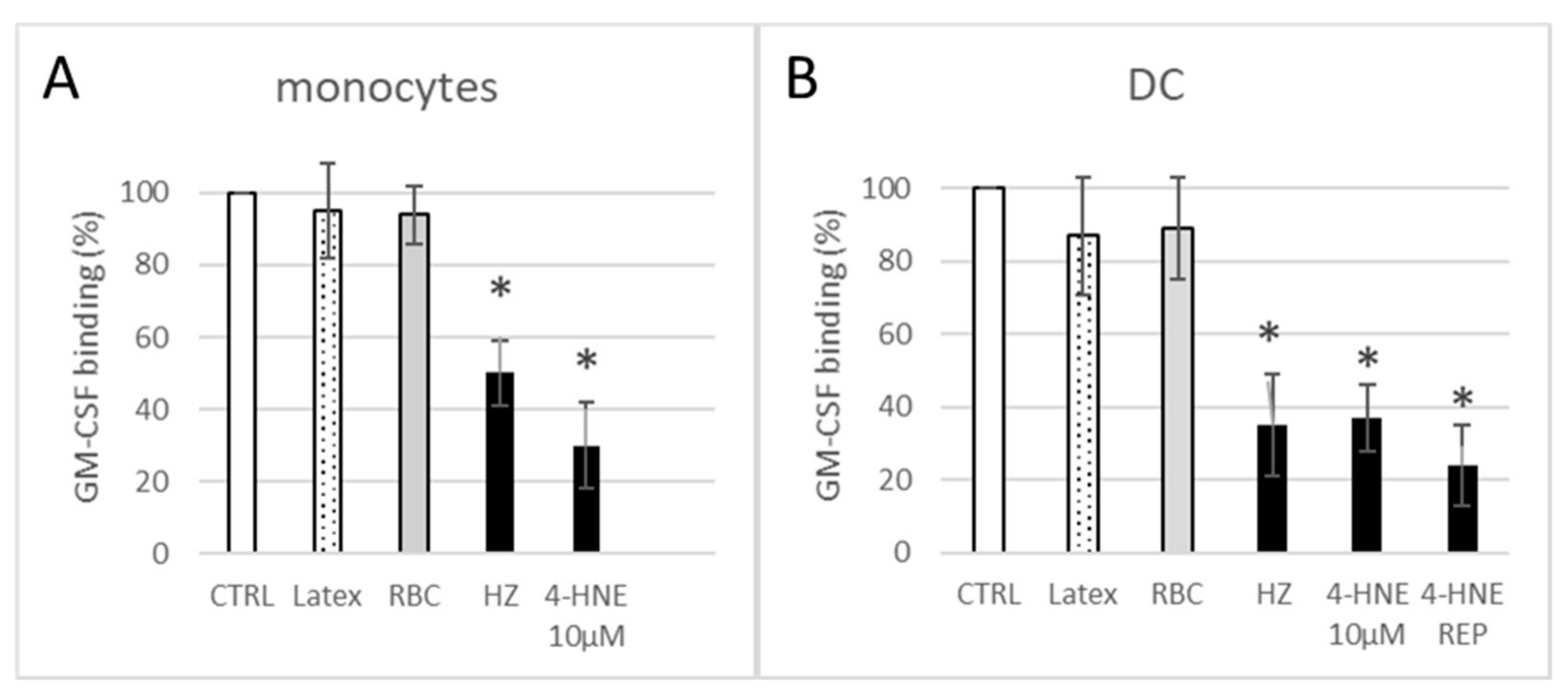
3.2. HZ and 4-HNE Impaired Functional Binding of Fluorescent GM-CSF
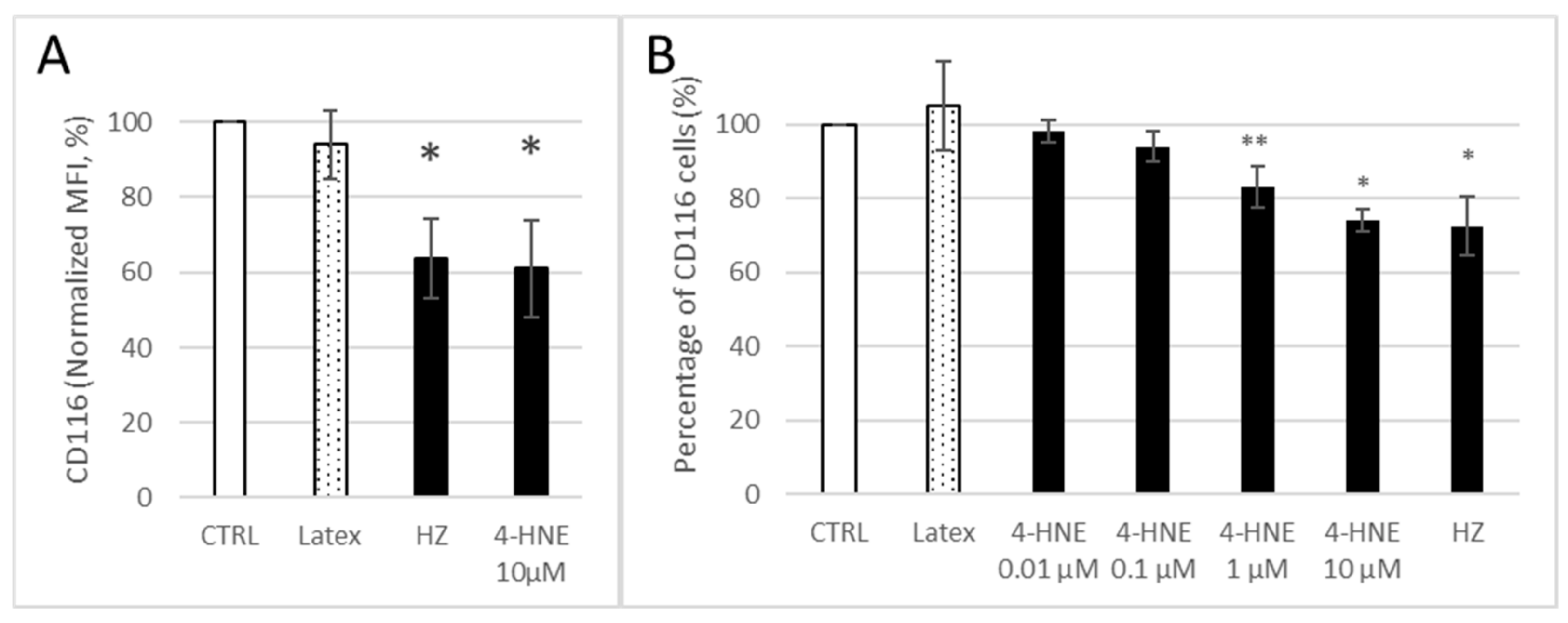
3.3. HZ and 4-HNE Dose-Dependently Decreased the Expression of GM-CSF R on Monocyte Surface
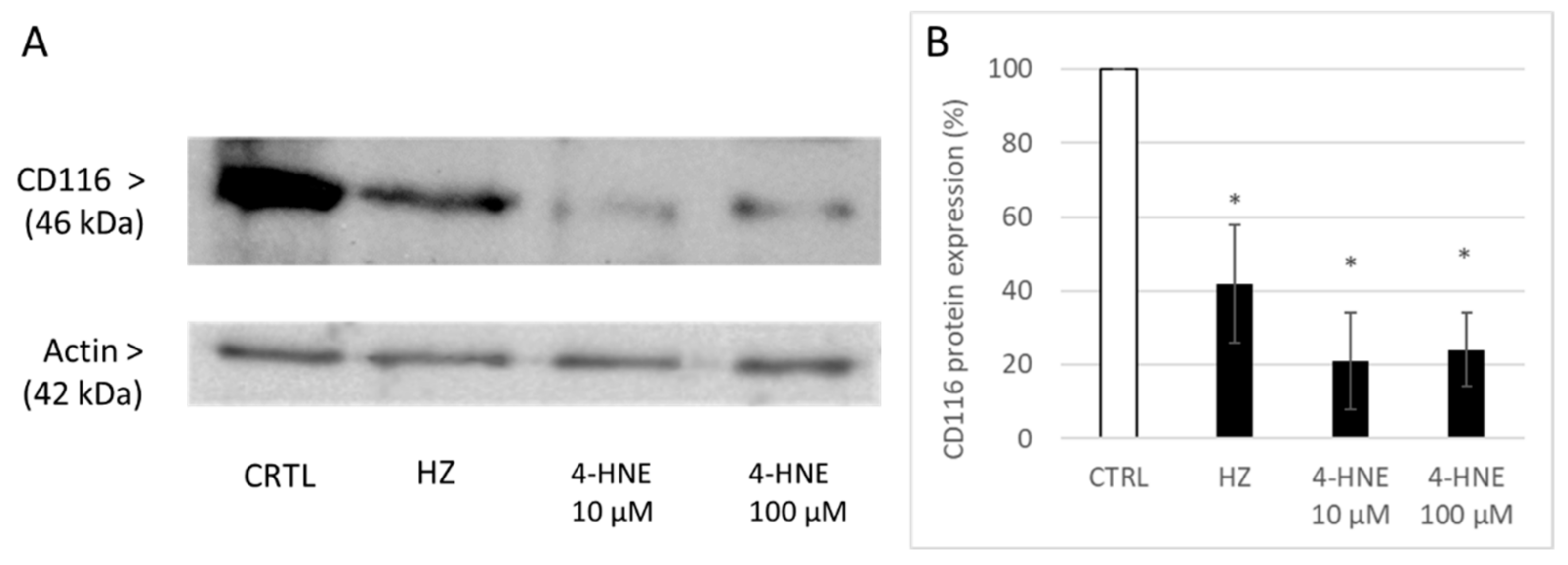
3.4. HZ and 4-HNE Decreased the Expression of GM-CSF R Detected in Cellular Protein Lysate
3.5. 4-HNE Modified Distinct Amino Acids in GM-CSF R
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ho, M.; Webster, H.K.; Looareesuwan, S.; Supanaranond, W.; Phillips, R.E.; Chanthavanich, P.; Warrell, D.A. Antigen-specific immunosuppression in human malaria due to plasmodium falciparum. J. Infect. Dis. 1986, 153, 763–771. [Google Scholar] [CrossRef]
- Urban, B.C.; Mwangi, T.; Ross, A.; Kinyanjui, S.; Mosobo, M.; Kai, O.; Lowe, B.; Marsh, K.; Roberts, D.J. Peripheral blood dendritic cells in children with acute Plasmodium falciparum malaria. Blood 2001, 98, 2859–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, M.M.; Riley, E.M. Innate immunity to malaria. Nat. Rev. Immunol. 2004, 4, 169–180. [Google Scholar] [CrossRef]
- Long, C.A.; Zavala, F. Immune responses in malaria. Cold Spring Harb. Perspect. Med. 2017, 7, a025577. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Hu, Z.; Yu, X. Accelerator or brake: Immune regulators in malaria. Front. Cell. Infect. Microbiol. 2020, 10, 610121. [Google Scholar] [CrossRef]
- Metzger, W.G.; Mordmüller, B.; Kremsner, P.G. Malaria pigment in leucocytes. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 637–638. [Google Scholar] [CrossRef]
- Birhanu, M.; Asres, Y.; Adissu, W.; Yemane, T.; Zemene, E.; Gedefaw, L. Hematological parameters and hemozoin-containing leukocytes and their association with disease severity among malaria infected children: A cross-sectional study at pawe general hospital, Northwest Ethiopia. Interdiscip. Perspect. Infect. Dis. 2017, 2017, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, E.; Turrini, F.; Ulliers, D.; Giribaldi, G.; Ginsburg, H.; Arese, P. Impairment of macrophage functions after ingestion of Plasmodium falciparum-infected erythrocytes or isolated malarial pigment. J. Exp. Med. 1992, 176, 1033–1041. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, E.; Müller, O.; Arese, P.; Siems, W.G.; Grune, T. Increased levels of 4-hydroxynonenal in human monocytes fed with malarial pigment hemozoin A possible clue for hemozoin toxicity. FEBS Lett. 1996, 388, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Green, M.D.; Xiao, L.; Lal, A.A. Formation of hydroxyeicosatetraenoic acids from hemozoin-catalyzed oxidation of arachidonic acid. Mol. Biochem. Parasitol. 1996, 83, 183–188. [Google Scholar] [CrossRef]
- Morakote, N.; Justus, D.E. Immunosuppression in malaria: Effect of hemozoin produced by Plasmodium berghei and Plasmodium falciparum. Int. Arch. Allergy Appl. Immunol. 1988, 86, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Scorza, T.; Magez, S.; Brys, L.; De Baetselier, P. Hemozoin is a key factor in the induction of malaria-associated immunosuppression. Parasite Immunol. 1999, 21, 545–554. [Google Scholar] [CrossRef]
- Barrera, V.; Skorokhod, O.A.; Baci, D.; Gremo, G.; Arese, P.; Schwarzer, E. Host fibrinogen stably bound to hemozoin rapidly activates monocytes via TLR-4 and CD11b/CD18-integrin: A new paradigm of hemozoin action. Blood 2011, 117, 5674–5682. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Pajares, A.; Rogerson, S.J. The rough guide to monocytes in malaria infection. Front. Immunol. 2018, 9, 2888. [Google Scholar] [CrossRef]
- Skorokhod, O.A.; Barrera, V.; Heller, R.; Carta, F.; Turrini, F.; Arese, P.; Schwarzer, E. Malarial pigment hemozoin impairs chemotactic motility and transendothelial migration of monocytes via 4-hydroxynonenal. Free. Radic. Biol. Med. 2014, 75, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Schwarzer, E.; Arese, P. Phagocytosis of malarial pigment hemozoin inhibits NADPH-oxidase activity in human monocyte-derived macrophages. Biochim. Biophys. Acta 1996, 1316, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, E.; Alessio, M.; Ulliers, D.; Arese, P. Phagocytosis of the malarial pigment, hemozoin, impairs expression of major histo-compatibility complex class II antigen, CD54, and CD11c in human monocytes. Infect. Immun. 1998, 66, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Skorokhod, O.A.; Alessio, M.; Mordmüller, B.; Arese, P.; Schwarzer, E. Hemozoin (malarial pigment) inhibits differentiation and maturation of human monocyte-derived dendritic cells: A peroxisome proliferator-activated receptor-gamma-mediated effect. J. Immunol. 2004, 173, 4066–4074. [Google Scholar] [CrossRef]
- Urban, B.C.; Todryk, S. Malaria pigment paralyzes dendritic cells. J. Biol. 2006, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Bujila, I.; Schwarzer, E.; Skorokhod, O.; Weidner, J.M.; Troye-Blomberg, M.; Östlund Farrants, A.K. Malaria-derived hemozoin exerts early modulatory effects on the phenotype and maturation of human dendritic cells. Cell Microbiol. 2016, 18, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, E.; Arese, P.; Skorokhod, O.A. Role of the lipoperoxidation product 4-hydroxynonenal in the pathogenesis of severe malaria anemia and malaria immunodepression. Oxid. Med. Cell. Longev. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Tilley, L.; Vennerstrom, J.L.; Roberts, D.; Rogerson, S.; Ginsburg, H. Oxidative stress in malaria parasite-infected erythrocytes: Host-parasite interactions. Int. J. Parasitol. 2004, 34, 163–189. [Google Scholar] [CrossRef]
- Percário, S.; Moreira, D.R.; Gomes, B.A.; Ferreira, M.E.; Gonçalves, A.C.; Laurindo, P.S.; Vilhena, T.C.; Dolabela, M.F.; Green, M.D. Oxidative stress in malaria. Int. J. Mol. Sci. 2012, 13, 16346–16372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acquah, S.; Boampong, J.N.; Eghan Jnr, B.A.E. Increased oxidative stress and inflammation independent of body adiposity in diabetic and nondiabetic controls in falciparum malaria. BioMed Res. Int. 2016, 2016, 5216913. [Google Scholar] [CrossRef] [Green Version]
- Kain, H.S.; Glennon, E.K.K.; Vijayan, K.; Arang, N.; Douglass, A.N.; Fortin, C.L.; Zuck, M.; Lewis, A.J.; Whiteside, S.L.; Dudgeon, D.R.; et al. Liver stage malaria infection is controlled by host regulators of lipid peroxidation. Cell Death Differ. 2020, 27, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Nanda, N.K.; Das, B.S. Presence of pro-oxidants in plasma of patients suffering from falciparum malaria. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 684–688. [Google Scholar] [CrossRef]
- Zhang, G.; Skorokhod, O.A.; Khoo, S.-K. Plasma advanced oxidative protein products are associated with anti-oxidative stress pathway genes and malaria in a longitudinal cohort. Malar. J. 2014, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Das, B.S.; Nanda, N.K. Evidence for erythrocyte lipid peroxidation in acute falciparum malaria. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 58–62. [Google Scholar] [CrossRef]
- Aguilar, R.; Marrocco, T.; Skorokhod, O.A.; Barbosa, A.; Nhabomba, A.; Manaca, M.N.; Guinovart, C.; Quintó, L.; Arese, P.; Alonso, P.L.; et al. Blood oxidative stress markers and Plasmodium falciparum malaria in non-immune African children. Br. J. Haematol. 2014, 164, 438–450. [Google Scholar] [CrossRef]
- Poli, G.; Schaur, R.J.; Siems, W.G.; Leonarduzzi, G.M. 4-hydroxynonenal: A membrane lipid oxidation product of medicinal interest. Med. Res. Rev. 2008, 28, 569–631. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Pizzimenti, S.; Ciamporcero, E.S.; Daga, M.; Ullio, C.; Arcaro, A.; Cetrangolo, G.P.; Ferretti, C.; Dianzani, C.; Lepore, A.; et al. Role of 4-hydroxynonenal-protein adducts in human diseases. Antioxid. Redox Signal. 2015, 22, 1681–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorokhod, O.A.; Schwarzer, E.; Grune, T.; Arese, P. Role of 4-hydroxynonenal in the hemozoin-mediated inhibition of differentiation of human monocytes to dendritic cells induced by GM-CSF/IL-4. Biofactors 2005, 24, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Parola, M.; Bellomo, G.; Robino, G.; Barrera, G.; Dianzani, M.U. 4-Hydroxynonenal as a biological signal: Molecular basis and patho-physiological implications. Antioxid. Redox Signal. 1999, 1, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Pizzimenti, S.; Toaldo, C.; Pettazzoni, P.; Dianzani, M.U.; Barrera, G. The “two-faced” effects of reactive oxygen species and the lipid pe-roxidation product 4-hydroxynonenal in the hallmarks of cancer. Cancers 2010, 2, 338–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorokhod, O.A.; Caione, L.; Marrocco, T.; Migliardi, G.; Barrera, V.; Arese, P.; Piacibello, W.; Schwarzer, E. Inhibition of erythropoiesis in malaria anemia: Role of hemozoin and hemozoin-generated 4-hydroxynonenal. Blood 2010, 116, 4328–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.A.; White, S.A.; Miller, J.J.; Toner, C.; Withington, S.; Newland, A.C.; Kelsey, S.M. Granulocyte-macrophage colony-stimulating factor induces activation and restores respiratory burst activity in monocytes from septic patients. J. Infect. Dis. 1998, 177, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Leino, L.; Nuutila, J.; Pelliniemi, T.T.; Rajamäki, A. Human recombinant GM-CSF selectively primes receptor mediated respiratory burst of neutrophils in vitro. Immunol. Lett. 1993, 38, 26–31. [Google Scholar] [CrossRef]
- Becher, B.; Tugues, S.; Greter, M. GM-CSF: From growth factor to central mediator of tissue inflammation. Immunity 2016, 45, 963–973. [Google Scholar] [CrossRef] [Green Version]
- Guthridge, M.A.; Stomski, F.C.; Thomas, D.; Woodcock, J.M.; Bagley, C.J.; Berndt, M.C.; Lopez, A.F. Mechanism of activation of the GM-CSF, IL-3, and IL-5 family of receptors. Stem Cells 1998, 16, 301–313. [Google Scholar] [CrossRef]
- Woodcock, J.M.; McClure, B.J.; Stomski, F.C.; Elliott, M.J.; Bagley, C.J.; Lopez, A.F. The human granulocyte-macrophage colony-stimulating factor (GM-CSF) receptor exists as a preformed receptor complex that can be activated by GM-CSF, interleukin-3, or interleukin-5. Blood 1997, 90, 3005–3017. [Google Scholar] [CrossRef]
- MC Lee, K.; Achuthan, A.A.; Hamilton, J.A. GM-CSF: A Promising Target in Inflammation and Autoimmunity. ImmunoTargets Ther. 2020, 9, 225–240. [Google Scholar] [CrossRef]
- Isom, A.L.; Barnes, S.; Wilson, L.; Kirk, M.; Coward, L.; Darley-Usmar, V. Modification of cytochrome c by 4-hydroxy-2-nonenal: Evidence for histidine, lysine, and arginine-aldehyde adducts. J. Am. Soc. Mass Spectrom. 2004, 15, 1136–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzer, E.; Turrini, F.; Arese, P. A luminescence method for the quantitative determination of phagocytosis of erythrocytes, of malaria-parasitized erythrocytes and of malarial pigment. Br. J. Haematol. 1994, 88, 740–745. [Google Scholar] [CrossRef]
- Lanza, F.; Castagnari, B.; Rigolin, G.; Moretti, S.; Latorraca, A.; Ferrari, L.; Bardi, A.; Castoldi, G. Flow cytometry measurement of GM-CSF receptors in acute leukemic blasts, and normal hemopoietic cells. Leukemia 1997, 11, 1700–1710. [Google Scholar] [CrossRef] [Green Version]
- Skorokhod, O.A.; Davalos-Schafler, D.; Gallo, V.; Valente, E.; Ulliers, D.; Notarpietro, A.; Mandili, G.; Novelli, F.; Persico, M.; Taglialatela-Scafati, O.; et al. Oxidative stress-mediated antimalarial activity of plakortin, a natural endoperoxide from the tropical sponge Plakortis simplex. Free. Radic. Biol. Med. 2015, 89, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Revelli, A.; Canosa, S.; Bergandi, L.; Skorokhod, O.A.; Biasoni, V.; Carosso, A.; Bertagna, A.; Maule, M.; Aldieri, E.; D’Eufemia, M.D.; et al. Oocyte polarized light microscopy, assay of specific follicular fluid metabolites, and gene expression in cumulus cells as different approaches to predict fertilization efficiency after ICSI. Reprod. Biol. Endocrinol. 2017, 15, 1–12. [Google Scholar] [CrossRef]
- UniProt. Available online: https://www.uniprot.org/uniprot/P15509 (accessed on 29 July 2021).
- Hansen, G.; Hercus, T.R.; McClure, B.J.; Stomski, F.C.; Dottore, M.; Powell, J.; Ramshaw, H.; Woodcock, J.; Xu, Y.; Guthridge, M.; et al. The structure of the GM-CSF receptor complex reveals a distinct mode of cytokine receptor activation. Cell 2008, 134, 496–507. [Google Scholar] [CrossRef]
- Pizzimenti, S.; Laurora, S.; Briatore, F.; Ferretti, C.; Dianzani, M.U.; Barrera, G. Synergistic effect of 4-hydroxynonenal and PPAR ligands in controlling human leukemic cell growth and differentiation. Free. Radic. Biol. Med. 2002, 32, 233–245. [Google Scholar] [CrossRef]
- Pott, S.; Kamrani, N.K.; Bourque, G.; Pettersson, S.; Liu, E.T. PPARG binding landscapes in macrophages suggest a genome-wide con-tribution of PU.1 to divergent PPARG binding in human and mouse. PLoS ONE 2012, 7, e48102. [Google Scholar] [CrossRef] [Green Version]
- Prevost, J.M.; Pelley, J.L.; Zhu, W.; D’Egidio, G.E.; Beaudry, P.P.; Pihl, C.; Neely, G.G.; Claret, E.; Wijdenes, J.; Bown, C.B. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and inflammatory stimuli up-regulate secretion of the soluble GM-CSF receptor in human monocytes: Evidence for ectodomain shedding of the cell surface GM-CSF receptor alpha subunit. J. Immunol. 2002, 169, 5679–5688. [Google Scholar] [CrossRef] [Green Version]
- Prato, M.; Gallo, V.; Giribaldi, G.; Arese, P. Phagocytosis of haemozoin (malarial pigment) enhances metalloproteinase-9 activity in human adherent monocytes: Role of IL-1beta and 15-HETE. Malar. J. 2008, 7, 157. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.C.; Applewhite, L.; Mitchell, P.O.; Fernainy, K.; Roman, J.; Eaton, D.C.; Guidot, D.M. GM-CSF receptor expression and signaling is decreased in lungs of ethanol-fed rats. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2006, 291, L1150–L1158. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, J.I.; Kominsky, D.J.; Jacobson, N.; Bowers, B.; Regalia, K.; Austin, G.L.; Yousefi, M.; Falta, M.T.; Fontenot, A.P.; Gerich, M.E.; et al. Defective leukocyte GM-CSF receptor (CD116) expression and function in inflammatory bowel disease. Gastroenterology 2011, 141, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, D.W.; Hamilton, R.F., Jr.; Holian, A. 4-Hydroxynonenal inhibits interleukin-1 beta converting enzyme. J. Interferon Cytokine Res. 1997, 17, 205–210. [Google Scholar] [CrossRef]
- Ringwald, P.; Peyron, F.; Vuillez, J.P.; Touze, J.E.; Le Bras, J.; Deloron, P. Levels of cytokines in plasma during Plasmodium falciparum malaria attacks. J. Clin. Microbiol. 1991, 29, 2076–2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, S.S.; Chorro, L.; Chan, J.; Lauvau, G. Splenic innate B1 B cell plasmablasts produce sustained granulocyte-macrophage colo-ny-stimulating factor and interleukin-3 cytokines during murine malaria infections. Infect. Immun. 2019, 87, e00482-19. [Google Scholar] [CrossRef] [PubMed]
- Donahue, R.E.; Clark, S.C. Granulocyte colony-stimulating factors as therapeutic agents. Immunol. Ser. 1992, 57, 637–649. [Google Scholar] [PubMed]
- Kumaratilake, L.M.; Ferrante, A.; Jaeger, T.; Rzepczyk, C. GM-CSF-induced priming of human neutrophils for enhanced phagocytosis and killing of asexual blood stages of Plasmodium falciparum: Synergistic effects of GM-CSF and TNF. Parasite Immunol. 1996, 18, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Richie, T.L.; Charoenvit, Y.; Wang, R.; Epstein, J.E.; Hedstrom, R.C.; Kumar, S.; Luke, T.C.; Freilich, D.A.; Aguiar, J.C.; Sacci, J.B., Jr.; et al. Clinical trial in healthy malaria-naïve adults to evaluate the safety, tolerability, immunogenicity and efficacy of MuStDO5, a five-gene, sporozoite/hepatic stage Plasmodium falciparum DNA vaccine combined with escalating dose human GM-CSF DNA. Hum. Vaccin. Immunother. 2012, 8, 1564–1584. [Google Scholar] [CrossRef] [Green Version]
- Ananth, N.; Shetty, B.V.; Vasudevan, D.M. Possible role of granulocyte macrophage colony stimulating factor receptor (GM-CSF R) in malaria. Indian J. Exp. Biol. 2003, 41, 357–359. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skorokhod, O.; Barrera, V.; Mandili, G.; Costanza, F.; Valente, E.; Ulliers, D.; Schwarzer, E. Malaria Pigment Hemozoin Impairs GM-CSF Receptor Expression and Function by 4-Hydroxynonenal. Antioxidants 2021, 10, 1259. https://doi.org/10.3390/antiox10081259
Skorokhod O, Barrera V, Mandili G, Costanza F, Valente E, Ulliers D, Schwarzer E. Malaria Pigment Hemozoin Impairs GM-CSF Receptor Expression and Function by 4-Hydroxynonenal. Antioxidants. 2021; 10(8):1259. https://doi.org/10.3390/antiox10081259
Chicago/Turabian StyleSkorokhod, Oleksii, Valentina Barrera, Giorgia Mandili, Federica Costanza, Elena Valente, Daniela Ulliers, and Evelin Schwarzer. 2021. "Malaria Pigment Hemozoin Impairs GM-CSF Receptor Expression and Function by 4-Hydroxynonenal" Antioxidants 10, no. 8: 1259. https://doi.org/10.3390/antiox10081259
APA StyleSkorokhod, O., Barrera, V., Mandili, G., Costanza, F., Valente, E., Ulliers, D., & Schwarzer, E. (2021). Malaria Pigment Hemozoin Impairs GM-CSF Receptor Expression and Function by 4-Hydroxynonenal. Antioxidants, 10(8), 1259. https://doi.org/10.3390/antiox10081259