
Anti-Allergic Effects of Myrciaria dubia (Camu-Camu) Fruit Extract by Inhibiting Histamine H1 and H4 Receptors and Histidine Decarboxylase in RBL-2H3 Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. High-Performance Liquid Chromatography Analysis
2.3. Sample Preparation
2.4. Cell Culture, Camu-Camu Fruit Treatment, and Stimulation
2.5. Cell Viability Assay
2.6. β-Hexosaminidase Release Assay
2.7. Histamine Release Assay
2.8. Measurement of Intracellular ROS Production
2.9. Reverse Transcription-Polymerase Chain Reaction
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
3.1. Quantification of Active Components from Camu-Camu Fruit Extract
3.2. Effects of Camu-Camu Fruit Extract on Cell Viability, β-Hexosaminidase, and Histamine Release
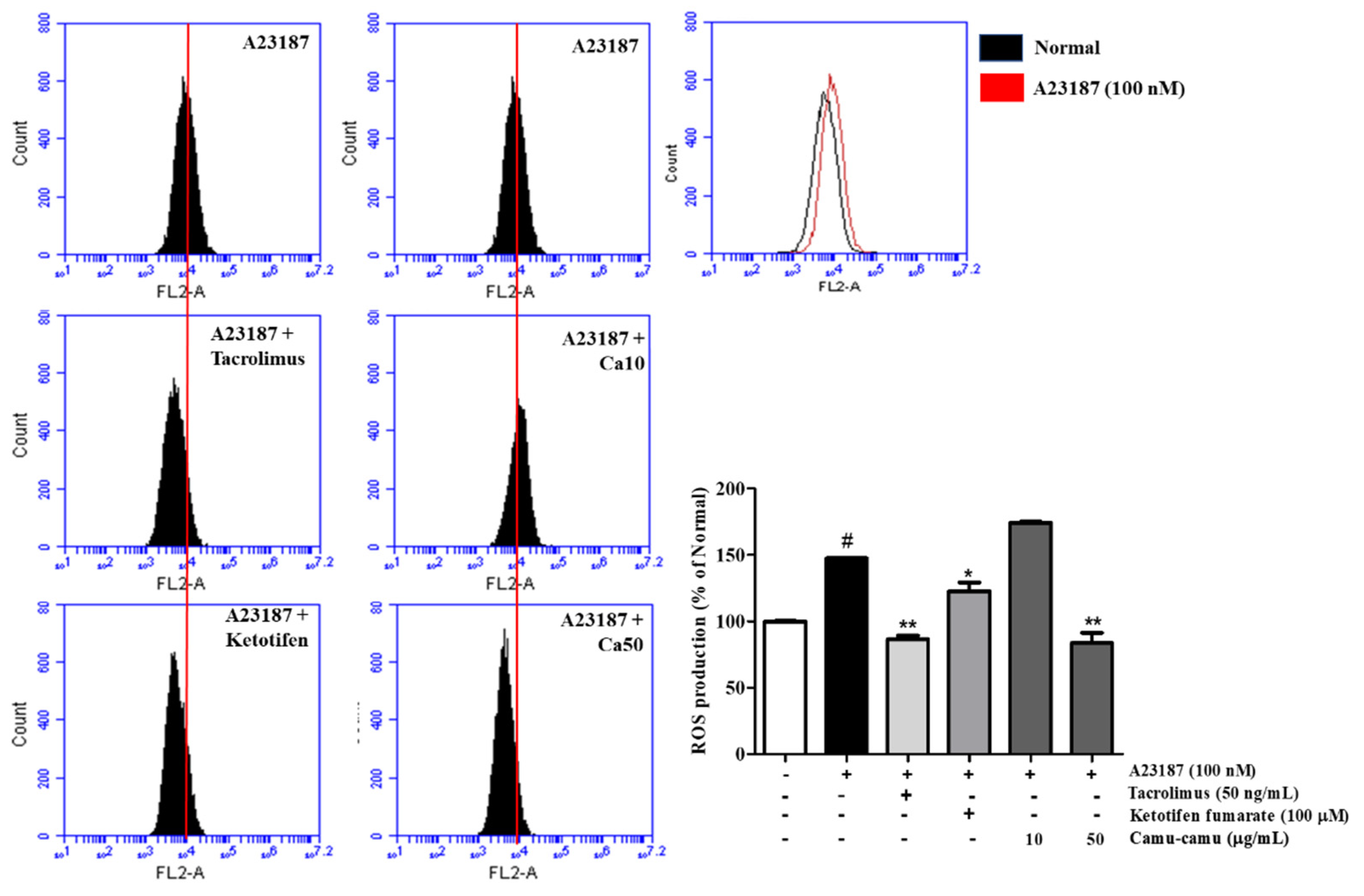
3.3. Effect of Camu-Camu Fruit Extract on Intracellular ROS Production
3.4. Effect of Camu-Camu Fruit Extract on mRNA Expression of H1R and H4R
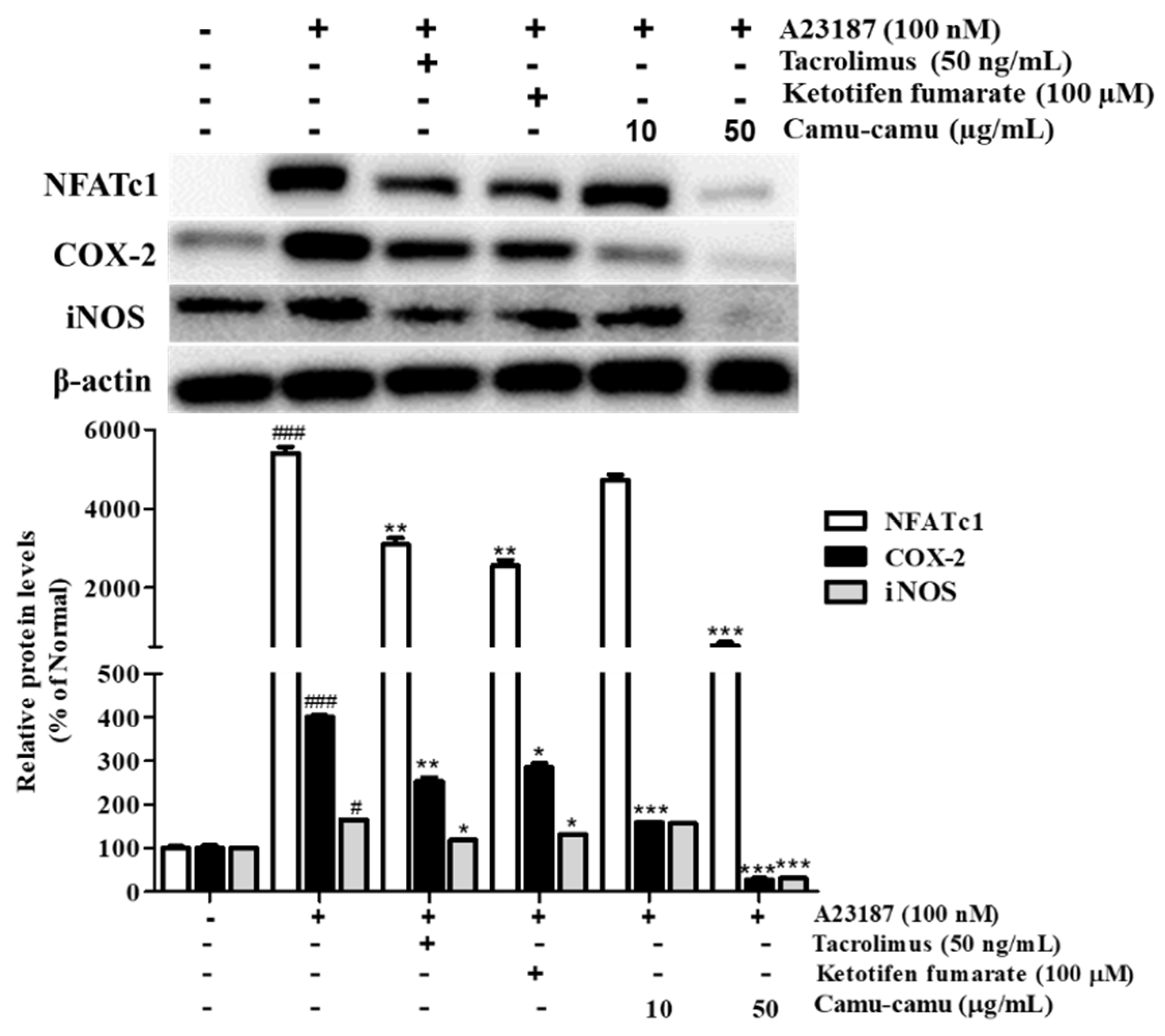
3.5. Effect of Camu-Camu Fruit Extract on mRNA and Protein Expression of HDC
3.6. Effect of Camu-Camu Fruit Extract on Calcium Channel Protein Expression
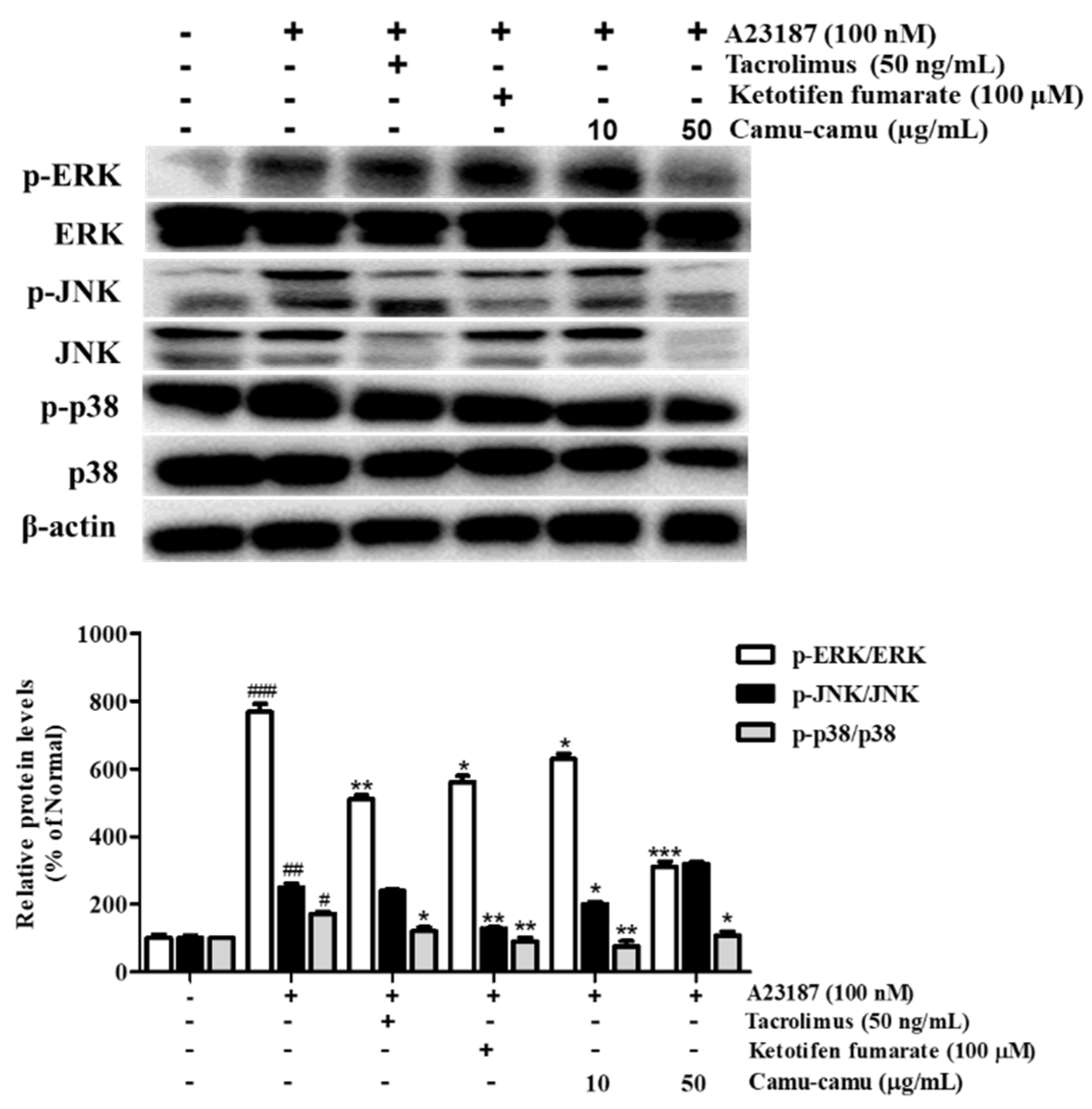
3.7. Effect of Camu-Camu Fruit Extract on MAPK Activation
3.8. Effect of Camu-Camu Fruit Extract on the Expression of KLF4/SP1 and GATA2/MITF Transcription Factors
3.9. Effect of Camu-Camu Fruit Extract on Nrf2 Activation
3.10. Effect of Camu-Camu Fruit Extract on NFAT Activation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galli, S.J.; Tsai, M. IgE and Mast Cells in Allergic Disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Parekh, A.B. Store-Operated CRAC Channels: Function in Health and Disease. Nat. Rev. Drug. Discov. 2010, 9, 399–410. [Google Scholar] [CrossRef]
- Wu, T.-F.; Chan, Y.-Y.; Shi, W.-Y.; Jhong, M.-T. Uncovering the Molecular Mechanism of Anti-Allergic Activity of Silkworm Pupa-Grown Cordyceps Militaris Fruit Body. Am. J. Chin. Med. 2017, 45, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Bryce, P.J.; Mathias, C.B.; Harrison, K.L.; Watanabe, T.; Geha, R.S.; Oettgen, H.C. The H1 Histamine Receptor Regulates Allergic Lung Responses. J. Clin. Investig. 2006, 116, 1624–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jemima, E.A.; Prema, A.; Thangam, E.B. Functional Characterization of Histamine H4 Receptor on Human Mast Cells. Mol. Immunol. 2014, 62, 19–28. [Google Scholar] [CrossRef]
- Grant, S.M.; Goa, K.L.; Fitton, A.; Sorkin, E.M. Ketotifen: A Review of Its Pharmacodynamic and Pharmacokinetic Properties, and Therapeutic Use in Asthma and Allergic Disorders. Drugs 1990, 40, 412–448. [Google Scholar] [CrossRef]
- Ohsawa, Y.; Hirasawa, N. The Role of Histamine H1 and H4 Receptors in Atopic Dermatitis: From Basic Research to Clinical Study. Allergol. Int. 2014, 63, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, H.; Nitta, Y.; Ueno, H.; Higuchi, Y. Structural Study Reveals That Ser-354 Determines Substrate Specificity on Human Histidine Decarboxylase. J. Biol. Chem. 2012, 287, 29175–29183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtsu, H.; Tanaka, S.; Terui, T.; Hori, Y.; Makabe-Kobayashi, Y.; Pejler, G.; Tchougounova, E.; Hellman, L.; Gertsenstein, M.; Hirasawa, N.; et al. Mice Lacking Histidine Decarboxylase Exhibit Abnormal Mast Cells. FEBS Lett. 2001, 502, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Ai, W.; Zheng, H.; Yang, X.; Liu, Y.; Wang, T.C. Tip60 Functions as a Potential Corepressor of KLF4 in Regulation of HDC Promoter Activity. Nucleic. Acids. Res. 2007, 35, 6137–6149. [Google Scholar] [CrossRef] [Green Version]
- Deaton, R.A.; Gan, Q.; Owens, G.K. Sp1-Dependent Activation of KLF4 Is Required for PDGF-BB-Induced Phenotypic Modulation of Smooth Muscle. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1027–H1037. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, B.; Harmacek, L.; Long, Z.; Liang, J.; Lukin, K.; Leach, S.M.; O’Connor, B.; Gerber, A.N.; Hagman, J.; et al. The Transcription Factors GATA2 and Microphthalmia-Associated Transcription Factor Regulate Hdc Gene Expression in Mast Cells and Are Required for IgE/Mast Cell–Mediated Anaphylaxis. J. Allergy Clin. Immunol. 2018, 142, 1173–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushima, M.; Takagi, K.; Ogawa, M.; Hirose, E.; Ota, Y.; Abe, F.; Baba, K.; Hasegawa, T.; Hasegawa, Y.; Kawabe, T. Heme Oxygenase-1 Mediates the Anti-Allergic Actions of Quercetin in Rodent Mast Cells. Inflamm. Res. 2009, 58, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the Master Regulator of Anti-Oxidative Responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley, P.C.; Pergolizzi, J.V.; Taylor, R.; Ridgway, C. Antioxidant and Associated Capacities of Camu Camu (Myrciaria Dubia): A Systematic Review. J. Altern. Complement. Med. 2015, 21, 8–14. [Google Scholar] [CrossRef]
- Hermann, M. The Impact of the European Novel Food Regulation on Trade and Food Innovation Based on Traditional Plant Foods from Developing Countries. Food Policy 2009, 34, 499–507. [Google Scholar] [CrossRef]
- Conceição, N.; Albuquerque, B.R.; Pereira, C.; Corrêa, R.C.G.; Lopes, C.B.; Calhelha, R.C.; Alves, M.J.; Barros, L.; CFR Ferreira, I. By-Products of Camu-Camu [Myrciaria Dubia (Kunth) McVaugh] as Promising Sources of Bioactive High Added-Value Food Ingredients: Functionalization of Yogurts. Molecules 2019, 25, 70. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, O.V.; Boleti, A.P.A.; Yuyama, L.K.O.; Lima, E.S. Effects of Diet Supplementation with Camu-Camu (Myrciaria Dubia HBK McVaugh) Fruit in a Rat Model of Diet-Induced Obesity. An. Acad. Bras. Cienc. 2013, 85, 355–363. [Google Scholar] [CrossRef]
- Do, N.Q.; Zheng, S.; Park, B.; Nguyen, Q.T.N.; Choi, B.-R.; Fang, M.; Kim, M.; Jeong, J.; Choi, J.; Yang, S.-J.; et al. Camu-Camu Fruit Extract Inhibits Oxidative Stress and Inflammatory Responses by Regulating NFAT and Nrf2 Signaling Pathways in High Glucose-Induced Human Keratinocytes. Molecules 2021, 26, 3174. [Google Scholar] [CrossRef]
- Ngo, H.T.T.; Hwang, E.; Kang, H.; Park, B.; Seo, S.A.; Yi, T.-H. Anti-Inflammatory Effects of Achillea Millefolium on Atopic Dermatitis-Like Skin Lesions in NC/Nga Mice. Am. J. Chin. Med. 2020, 48, 1121–1140. [Google Scholar] [CrossRef]
- Nishimura, K.; Okamoto, M.; Shibue, R.; Mizuta, T.; Shibayama, T.; Yoshino, T.; Murakami, T.; Yamaguchi, M.; Tanaka, S.; Toida, T.; et al. KLF4 Is Required for Suppression of Histamine Synthesis by Polyamines during Bone Marrow-Derived Mast Cell Differentiation. PLoS ONE 2020, 15, e0229744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q. Role of Nrf2 in Oxidative Stress and Toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, N. Expression of Histidine Decarboxylase and Its Roles in Inflammation. Int. J. Mol. Sci. 2019, 20, 376. [Google Scholar] [CrossRef] [Green Version]
- Holowka, D.; Calloway, N.; Cohen, R.; Gadi, D.; Lee, J.; Smith, N.L.; Baird, B. Roles for Ca2+ Mobilization and Its Regulation in Mast Cell Functions. Front. Immun. 2012, 3, 104. [Google Scholar] [CrossRef] [Green Version]
- Monticelli, S.; Solymar, D.C.; Rao, A. Role of NFAT Proteins in IL13 Gene Transcription in Mast Cells. J. Biol. Chem. 2004, 279, 36210–36218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.K.; Kim, H.S.; Kim, A.-R.; Kim, J.H.; Kim, B.; Noh, G.; Kim, H.S.; Beaven, M.A.; Kim, Y.M.; Choi, W.S. DJ-1 Regulates Mast Cell Activation and IgE-Mediated Allergic Responses. J. Allergy. Clin. Immunol. 2013, 131, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Gan, X.; Su, G.; Wanling, G.; Li, S.; Hei, Z.; Yang, C.; Wang, H. The Interaction between Oxidative Stress and Mast Cell Activation Plays a Role in Acute Lung Injuries Induced by Intestinal Ischemia–Reperfusion. J. Surg. Res. 2014, 187, 542–552. [Google Scholar] [CrossRef]
- Calabrese, V.; Copani, A.; Testa, D.; Ravagna, A.; Spadaro, F.; Tendi, E.; Nicoletti, V.G.; Giuffrida Stella, A.M. Nitric Oxide Synthase Induction in Astroglial Cell Cultures: Effect on Heat Shock Protein 70 Synthesis and Oxidant/Antioxidant Balance. J. Neurosci. Res. 2000, 60, 613–622. [Google Scholar] [CrossRef]
- Calabrese, V.; Cornelius, C.; Dinkova-Kostova, A.T.; Calabrese, E.J.; Mattson, M.P. Cellular Stress Responses, the Hormesis Paradigm, and Vitagenes: Novel Targets for Therapeutic Intervention in Neurodegenerative Disorders. Antioxid. Redox. Signal. 2010, 13, 1763–1811. [Google Scholar] [CrossRef]
- Dattilo, S.; Mancuso, C.; Koverech, G.; Di Mauro, P.; Ontario, M.L.; Petralia, C.C.; Petralia, A.; Maiolino, L.; Serra, A.; Calabrese, E.J.; et al. Heat Shock Proteins and Hormesis in the Diagnosis and Treatment of Neurodegenerative Diseases. Immun. Ageing. 2015, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Hirasawa, N.; Torigoe, M.; Ohgawara, R.; Murakami, A.; Ohuchi, K. Involvement of MAP Kinases in Lipopolysaccharide-Induced Histamine Production in RAW 264 Cells. Life Sci. 2006, 80, 36–42. [Google Scholar] [CrossRef]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef] [Green Version]
- Andoh, T.; Zhang, Q.; Yamamoto, T.; Tayama, M.; Hattori, M.; Tanaka, K.; Kuraishi, Y. Inhibitory Effects of the Methanol Extract of Ganoderma Lucidum on Mosquito Allergy-Induced Itch-Associated Responses in Mice. J. Pharmacol. Sci. 2010, 114, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollbracht, C.; Raithel, M.; Krick, B.; Kraft, K.; Hagel, A.F. Intravenous Vitamin C in the Treatment of Allergies: An Interim Subgroup Analysis of a Long-Term Observational Study. J. Int. Med. Res. 2018, 46, 3640–3655. [Google Scholar] [CrossRef] [PubMed]
- Pyo, M.Y.; Yoon, S.J.; Yu, Y.; Park, S.; Jin, M. Cyanidin-3-Glucoside Suppresses Th2 Cytokines and GATA-3 Transcription Factor in EL-4 T Cells. Biosci. Biotechnol. Biochem. 2014, 78, 1037–1043. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, N.Q.; Zheng, S.; Oh, S.; Nguyen, Q.T.N.; Fang, M.; Kim, M.; Choi, J.; Kim, M.-J.; Jeong, J.; Yi, T.-H. Anti-Allergic Effects of Myrciaria dubia (Camu-Camu) Fruit Extract by Inhibiting Histamine H1 and H4 Receptors and Histidine Decarboxylase in RBL-2H3 Cells. Antioxidants 2022, 11, 104. https://doi.org/10.3390/antiox11010104
Do NQ, Zheng S, Oh S, Nguyen QTN, Fang M, Kim M, Choi J, Kim M-J, Jeong J, Yi T-H. Anti-Allergic Effects of Myrciaria dubia (Camu-Camu) Fruit Extract by Inhibiting Histamine H1 and H4 Receptors and Histidine Decarboxylase in RBL-2H3 Cells. Antioxidants. 2022; 11(1):104. https://doi.org/10.3390/antiox11010104
Chicago/Turabian StyleDo, Nhung Quynh, Shengdao Zheng, Sarang Oh, Quynh T. N. Nguyen, Minzhe Fang, Minseon Kim, Junhui Choi, Myeong-Ju Kim, Jeehaeng Jeong, and Tae-Hoo Yi. 2022. "Anti-Allergic Effects of Myrciaria dubia (Camu-Camu) Fruit Extract by Inhibiting Histamine H1 and H4 Receptors and Histidine Decarboxylase in RBL-2H3 Cells" Antioxidants 11, no. 1: 104. https://doi.org/10.3390/antiox11010104
APA StyleDo, N. Q., Zheng, S., Oh, S., Nguyen, Q. T. N., Fang, M., Kim, M., Choi, J., Kim, M.-J., Jeong, J., & Yi, T.-H. (2022). Anti-Allergic Effects of Myrciaria dubia (Camu-Camu) Fruit Extract by Inhibiting Histamine H1 and H4 Receptors and Histidine Decarboxylase in RBL-2H3 Cells. Antioxidants, 11(1), 104. https://doi.org/10.3390/antiox11010104