
Variability of Phenolic Compound Accumulation and Antioxidant Activity in Wild Plants of Some Rumex Species (Polygonaceae)

 ,
,  ,
, 
 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extract Preparation
2.3. Determination of Total Contents of Some Groups of Phenolic Compounds
2.3.1. Determination of Total Phenolic Content
2.3.2. Determination of Total Flavonoid Content
2.3.3. Determination of Total Content of Hydroxycinnamic Acids
2.3.4. Determination of Total Content of Proanthocyanidins
2.3.5. Determination of Total Catechin Content
2.3.6. Determination of Total Tannin Content
2.4. High-Performance Liquid Chromatography with Diode-Array Detection (HPLC-DAD) Analysis of Individual Phenolic Compounds
2.5. Determination of Antioxidant Activity
2.6. Statistical Analysis
3. Results
3.1. Variation in the Content of Some Groups of Phenolic Compounds
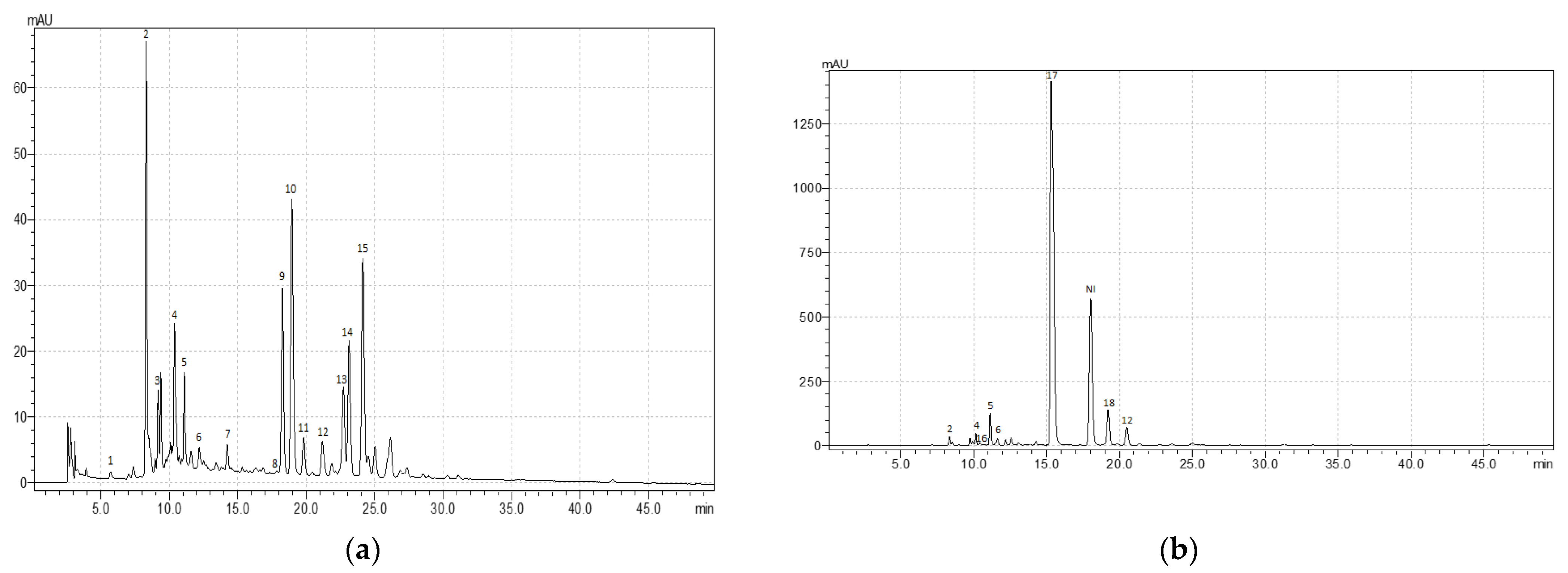
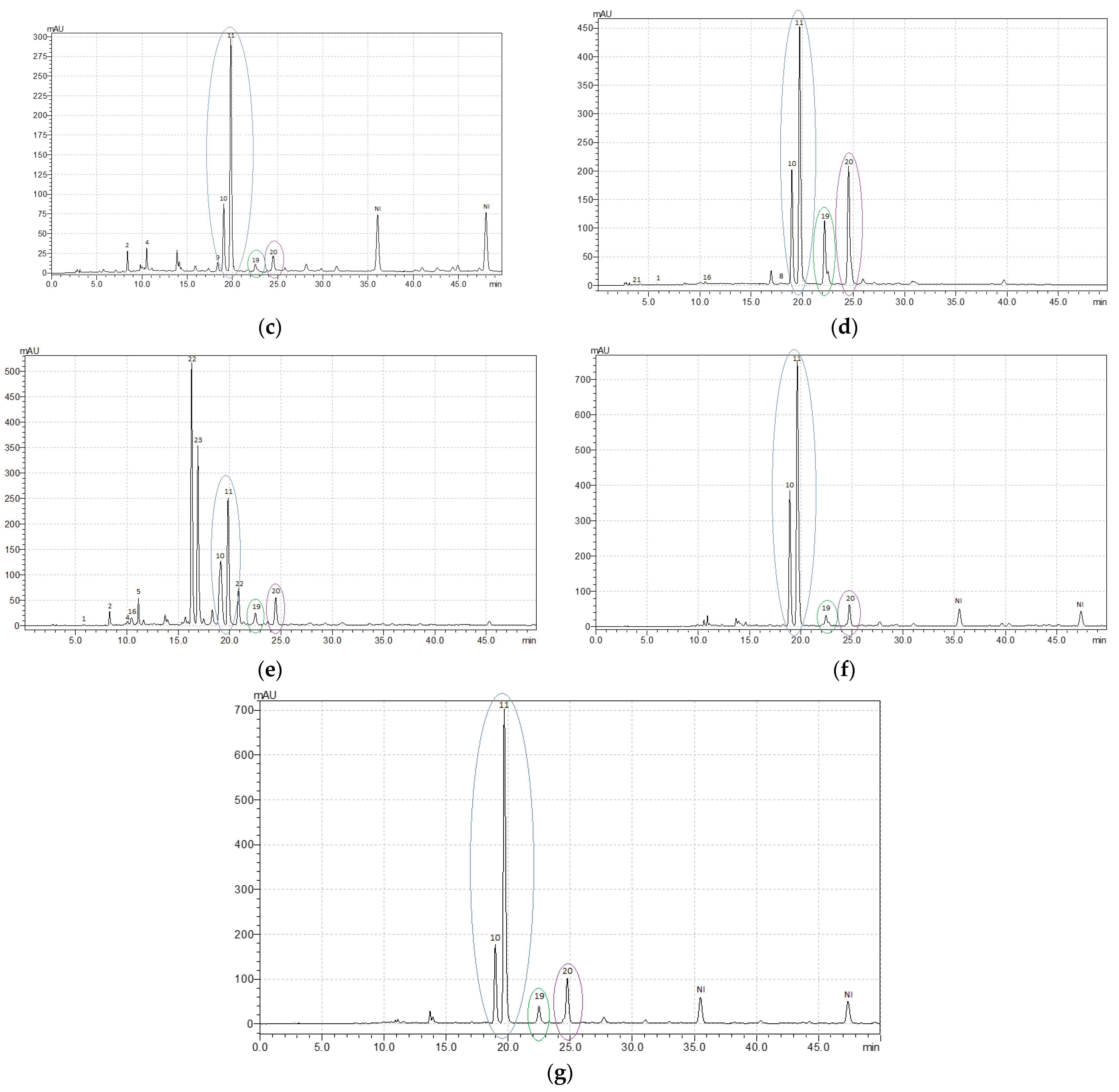
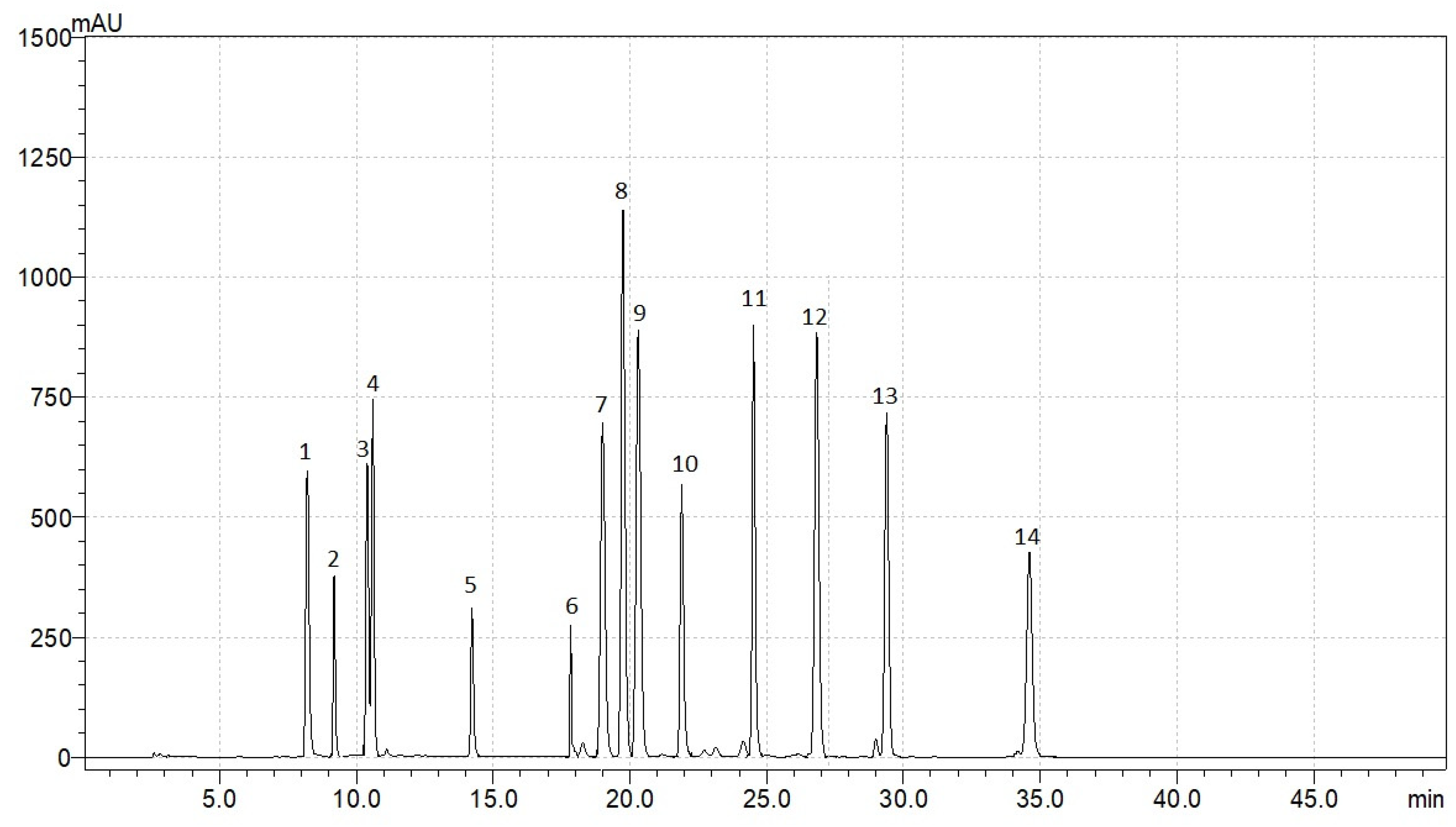
3.2. Variation in the Content of Individual Phenolic Compounds
3.3. Antioxidant Activity of the Rumex Extracts
3.4. Correlation between Phenolic Compounds Content and Antioxidant Activity
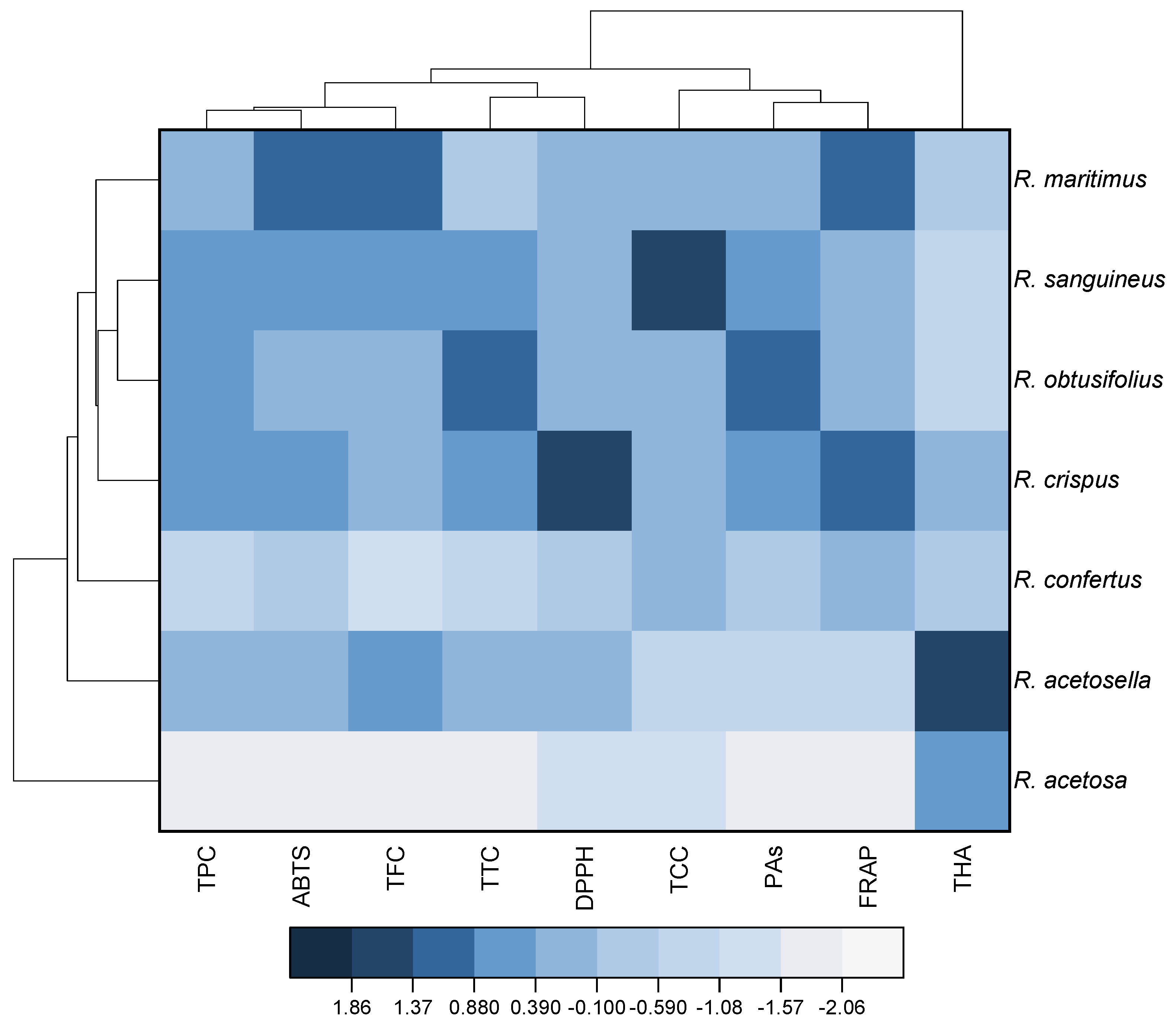
3.5. Heat Map and Cluster Analysis of Studied Rumex Species Based on the Content of Phenolic Compounds and Antioxidant Activity of Their Extracts
4. Discussion
4.1. Approaching the Problem of the Rumex Taxonomy
4.2. The Rumex Plants as a Resource Object Specificity of Use
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A



References
- Carvalho, A.M.; Barata, A.M. The consumption of wild edible plants. In Wild Plants, Mushrooms and Nuts: Functional Food Properties and Applications; Ferreira, I.C.F.R., Morales, P., Barros, L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 159–198. [Google Scholar]
- Ceccanti, C.; Landi, M.; Benvenuti, S.; Pardossi, A.; Guidi, L. Mediterranean Wild Edible Plants: Weeds or “New Functional Crops”? Molecules 2018, 23, 2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puccinelli, M.; Pezzarossa, B.; Pintimalli, L.; Malorgio, F. Selenium Biofortification of Three Wild Species, Rumex acetosa L., Plantago coronopus L., and Portulaca oleracea L., Grown as Microgreens. Agronomy 2021, 11, 1155. [Google Scholar] [CrossRef]
- Faehnrich, B.; Franz, C.; Nemaz, P.; Kaul, H.-P. Medicinal plants and their secondary metabolites—State of the art and trends in breeding, analytics and use in feed supplementation—with special focus on German chamomile. J. Appl. Bot. Food Qual. 2021, 94, 61–74. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Myles, S. Exploiting Wild Relatives for Genomics-assisted Breeding of Perennial Crops. Front. Plant Sci. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, O.M.; Fasinu, P.S.; Bello, O.E.; Ogbesejana, A.B.; Adetunji, C.O.; Dada, A.O.; Ibitoye, O.S.; Aloko, S.; Oguntoye, O.S. Wild vegetable Rumex acetosa Linn.: Its ethnobotany, pharmacology and phytochemistry—A review. S. Afr. J. Bot. 2019, 125, 149–160. [Google Scholar] [CrossRef]
- Vasas, A.; Orbán-Gyapai, O.; Hohmann, J. The Genus Rumex: Review of traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2015, 175, 198–228. [Google Scholar] [CrossRef]
- Podgurskaya, V.V.; Luksha, E.A.; Gushchina, E.S.; Savchenko, I.A.; Korneeva, I.N.; Kalinkina, G.I. Biological activity of the genus Rumex (Polygonaceae) plants. Chem. Plant Raw Mater. 2021, 2, 59–78. [Google Scholar] [CrossRef]
- Pinela, J.; Carvalho, A.M.; Ferreira, I.C.F.R. Wild edible plants: Nutritional and toxicological characteristics, retrieval strategies and importance for today’s society. Food Chem. Toxicol. 2017, 110, 165–188. [Google Scholar] [CrossRef] [Green Version]
- Wegiera, M.; Smolarz, H.D.; Wianowska, D.; Dawidowicz, A.L. Anthracene derivatives in some species of Rumex L. genus. Acta Soc. Bot. Pol. 2007, 76, 103–108. [Google Scholar]
- Upton, R.; Graff, A.; Jolliffe, G.; Länger, R.; Williamson, E. American Herbal Pharmacopoeia: Botanical Pharmacognosy-Microscopic Characterization of Botanical Medicines; CRC Press: Boca Raton, FL, USA, 2016; ISBN 1420073281. [Google Scholar]
- Gosudarstvennaya Farmakopeya Rossiyskoy Federatsii XIV izdaniye. In [State Pharmacopoeia of the Russian Federation XIV edition]; Emshanova, S.V.; Potanina, O.G.; Budanova, E.V.; Chistyakov, V.V. (Eds.) Ministry of Health of Russian Federation: Moscow, Russia, 2018; Volume 4, p. 1844. (In Russian) [Google Scholar]
- Shaikh, S.; Shriram, V.; Srivastav, A.; Barve, P.; Kumar, V. A critical review on Nepal Dock (Rumex nepalensis): A tropical herb with immense medicinal importance. Asian Pac. J. Trop. Med. 2018, 11, 405–414. [Google Scholar] [CrossRef]
- Prakash Mishra, A.; Sharifi-Rad, M.; Shariati, M.A.; Mabkhot, Y.N.; Al-Showiman, S.S.; Rauf, A.; Salehi, B.; Župunski, M.; Sharifi-Rad, M.; Gusain, P.; et al. Bioactive compounds and health benefits of edible Rumex species-A review. Cell. Mol. Biol. 2018, 64, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jiang, J.-G. Health functions and structure-activity relationships of natural anthraquinones from plants. Food Funct. 2018, 9, 6063–6080. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Muñoz, G.; Miranda, I.L.; Sartori, S.K.; de Rezende, D.C.; Diaz, M.A.N. Chapter 11—Anthraquinones: An overview. In Studies in Natural Products Chemistry; Atta-ur-Rahman, B.T.-S., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 58, pp. 313–338. ISBN 1572-5995. [Google Scholar]
- Balog, K.; Svirčev, E.; Lesjak, M.; Orcic, D.; Beara, I.; Francišković, M.; Simin, N. Phenolic profiling of Rumex L. species by means of the LC-MS/MS. Planta Med. 2012, 78, PL12. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Padhi, E.M.T.; Liu, R.; Hernandez, M.; Tsao, R.; Ramdath, D.D. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J. Funct. Foods 2017, 38, 602–611. [Google Scholar] [CrossRef]
- Sevket, A.L.P.; Ercisli, S.; Jurikova, T.; Cakir, O.; Gozlekci, S. Bioactive content of rose hips of different wildly grown Rosa dumalis genotypes. Not. Bot. Horti Agrobot. 2016, 44, 472–476. [Google Scholar]
- Štefan, M.B.; Rodríguez, J.V.; Blažeković, B.; Kindl, M.; Vladimir-Knežević, S. Total hydroxycinnamic acids assay: Prevalidation and application on Lamiaceae species. Food Anal. Methods 2014, 7, 326–336. [Google Scholar] [CrossRef]
- Chupin, L.; Motillon, C.; Charrier-El Bouhtoury, F.; Pizzi, A.; Charrier, B. Characterisation of maritime pine (Pinus pinaster) bark tannins extracted under different conditions by spectroscopic methods, FTIR and HPLC. Ind. Crops Prod. 2013, 49, 897–903. [Google Scholar] [CrossRef]
- He, Q.; Yao, K.; Jia, D.; Fan, H.; Liao, X.; Shi, B. Determination of total catechins in tea extracts by HPLC and spectrophotometry. Nat. Prod. Res. 2009, 23, 93–100. [Google Scholar] [CrossRef]
- Skrypnik, L.; Grigorev, N.; Michailov, D.; Antipina, M.; Danilova, M.; Pungin, A. Comparative study on radical scavenging activity and phenolic compounds content in water bark extracts of alder (Alnus glutinosa (L.) Gaertn.), oak (Quercus robur L.) and pine (Pinus sylvestris L.). Eur. J. Wood Wood Prod. 2019, 77, 879–890. [Google Scholar] [CrossRef]
- Skrypnik, L.; Novikova, A. Response surface modeling and optimization of polyphenols extraction from apple pomace based on nonionic emulsifiers. Agronomy 2020, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Feduraev, P.; Chupakhina, G.; Maslennikov, P.; Tacenko, N.; Skrypnik, L. Variation in Phenolic Compounds Content and Antioxidant Activity of Different Plant Organs from Rumex crispus L. and Rumex obtusifolius L. at Different Growth Stages. Antioxidants 2019, 8, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H. Flora Europaea: Plantaginaceae to Compositae (and Rubiaceae); Cambridge University Press: Cambridge, UK, 1964; Volume 4, ISBN 0521087171. [Google Scholar]
- Löve, Á.; Kapoor, B.M. A Chromosome Atlas of the Collective Genus Rumex. Cytologia 1967, 32, 328–342. [Google Scholar] [CrossRef] [Green Version]
- Rechinger, K.H. Rumex subgen. Rumex sect. Axillares (Polygonaceae) in South America. Plant Syst. Evol. 1990, 172, 151–192. [Google Scholar] [CrossRef]
- Rechinger, K.H. Rumex (Polygonaceae) in Australia: A reconsideration. Nuytsia 1984, 5, 75–122. [Google Scholar]
- Schuster, T.M.; Reveal, J.L.; Bayly, M.J.; Kron, K.A. An updated molecular phylogeny of Polygonoideae (Polygonaceae): Relationships of Oxygonum, Pteroxygonum, and Rumex, and a new circumscription of Koenigia. TAXON 2015, 64, 1188–1208. [Google Scholar] [CrossRef]
- Grant, K.D.; Koenemann, D.; Mansaray, J.; Ahmed, A.; Khamar, H.; El Oualidi, J.; Burke, J.M. Phylogeny of docks and sorrels Rumex, Polygonaceae) reveals plasticity of reproductive systems. bioRxiv 2020. [Google Scholar] [CrossRef]
- Yang, K.; Shang, M.; Jiang, Y.; Qian, J.; Duan, B.; Yang, Y. The complete chloroplast genome of Rumex hastatus D. Don and its phylogenetic analysis. Mitochondrial DNA Part B 2020, 5, 1681–1682. [Google Scholar] [CrossRef]
- Tsvelev, N.N. Genus Aconogonon (Meissn.) Reichenb. (Polygonaceae) in Eastern Europe and Northern Asia. Novosti Sist. Vyssh. Rast. 1993, 29, 55–65. [Google Scholar]
- Vysochina, G.I. Phenolic compounds in systematics and phylogeny of the family Polygonaceae Juss. IV. Genus Rumex L. Turczaninowia 2011, 14, 120. [Google Scholar]
- Navajas-Pérez, R.; de la Herrán, R.; López González, G.; Jamilena, M.; Lozano, R.; Ruiz Rejón, C.; Ruiz Rejón, M.; Garrido-Ramos, M.A. The Evolution of Reproductive Systems and Sex-Determining Mechanisms Within Rumex (Polygonaceae) Inferred from Nuclear and Chloroplastidial Sequence Data. Mol. Biol. Evol. 2005, 22, 1929–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapin, F.S., III; Schulze, E.D.; Mooney, H.A. The ecology and economics of storage in plants. Annu. Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Zaller, J.G. Ecology and non-chemical control of Rumex crispus and R. obtusifolius (Polygonaceae): A review. Weed Res. 2004, 44, 414–432. [Google Scholar] [CrossRef]
- Kucekova, Z.; Mlcek, J.; Humpolicek, P.; Rop, O.; Valasek, P.; Saha, P. Phenolic Compounds from Allium schoenoprasum, Tragopogon pratensis and Rumex acetosa and Their Antiproliferative Effects. Molecules 2011, 16, 9207–9217. [Google Scholar] [CrossRef]
- Aritomi, M.; Kiyota, I.; Mazaki, T. Flavonoid constituents in leaves of Rumex acetosa LINNAEUS and R. japonicus HOUTTUYN. Chem. Pharm. Bull. 1965, 13, 1470–1471. [Google Scholar] [CrossRef] [Green Version]
- Saleh, N.A.M.; El-Hadidi, M.N.; Arafa, R.F.M. Flavonoids and anthraquinones of some Egyptian Rumex species (Polygonaceae). Biochem. Syst. Ecol. 1993, 21, 301–303. [Google Scholar] [CrossRef]
- Choe, S.-G.; Hwang, B.-Y.; Kim, M.-S.; Oh, G.-J.; Lee, K.-S.; Ro, J.-S. Chemical Components of Rumex acetosella L. Korean J. Pharmacogn. 1998, 29, 209–216. [Google Scholar]
- Yıldırım, A.; Mavi, A.; Kara, A.A. Determination of Antioxidant and Antimicrobial Activities of Rumex crispus L. Extracts. J. Agric. Food Chem. 2001, 49, 4083–4089. [Google Scholar] [CrossRef]
- Jeong, G.-T.; Lee, K.-M.; Park, D.-H. Study of antimicrobial and antioxidant activities of Rumex crispus extract. Korean Chem. Eng. Res. 2006, 44, 81–86. [Google Scholar]
- Shagjjav, O.; Bhattarai, H.D.; Han Yim, J.; Dondog, P. Antioxidant Activity of Some Mongolian Plants. Mong. J. Biol. Sci. 2014, 12, 27–32. [Google Scholar]
- Bicker, J.; Petereit, F.; Hensel, A. Proanthocyanidins and a phloroglucinol derivative from Rumex acetosa L. Fitoterapia 2009, 80, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Jund, R.; Mondigler, M.; Steindl, H.; Stammer, H.; Stierna, P.; Bachert, C. Clinical efficacy of a dry extract of five herbal drugs in acute viral rhinosinusitis. Rhinology 2012, 50, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Karkhile, K.V.; Bhutani, K.K.; Jachak, S.M. Anti-inflammatory, cyclooxygenase (COX)-2, COX-1 inhibitory, and free radical scavenging effects of Rumex nepalensis. Planta Med. 2010, 76, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Harshaw, D.; Nahar, L.; Vadla, B.; Sarker, S.D. Bioactivity of Rumex obtusifolius (Polygonaceae). Arch. Biol. Sci. 2010, 62, 387–392. [Google Scholar] [CrossRef]
- Korpelainen, H.; Pietiläinen, M. Sorrel (Rumex acetosa L.): Not Only a Weed but a Promising Vegetable and Medicinal Plant. Bot. Rev. 2020, 86, 234–246. [Google Scholar] [CrossRef]
- Taulavuori, K.; Julkunen-Tiitto, R.; Hyöky, V.; Taulavuori, E. Blue Mood for Superfood. Nat. Prod. Commun. 2013, 8, 1934578X1300800627. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | TPC 1, mg GAE g–1 | TFC, mg RE g–1 | THA, mg CAE g–1 | TCC, mg CE g–1 | PAs, mg CyE g–1 | TTC, mg GAE g–1 |
|---|---|---|---|---|---|---|
| R. acetosa | 23 ± 2 | 18 ± 1 | 12.7 ± 0.6 | 0.90 ± 0.05 | 0.24 ± 0.02 | 0.46 ± 0.05 |
| R. acetosella | 117 ± 7 | 106 ± 4 | 18 ± 1 | 1.3 ± 0.1 | 2.2 ± 0.2 | 11 ± 1 |
| R. confertus | 76 ± 7 | 38± 2 | 4.8 ± 0.3 | 5.0 ± 0.3 | 4.0 ± 0.3 | 6.4 ± 0.3 |
| R. crispus | 131 ± 10 | 92± 5 | 8.9 ± 0.6 | 5.2 ± 0.3 | 6.4 ± 0.3 | 14 ± 1 |
| R. maritimus | 111 ± 6 | 120 ± 9 | 5.8 ± 0.6 | 4.8 ± 0.3 | 5.0 ± 0.4 | 7.1 ± 0.6 |
| R. obtusifolius | 129 ± 9 | 92 ± 4 | 1.9 ± 0.1 | 6.0 ± 0.4 | 7.2 ± 0.5 | 17 ± 1 |
| R. sanguineus | 126 ± 5 | 99 ± 6 | 1.9 ± 0.1 | 10.9 ± 0.6 | 6.6 ± 0.4 | 12.9 ± 0.7 |
| Compounds (Retention Time, Min) | Content of Individual Phenolic Compounds, mg g–1 | ||||||
|---|---|---|---|---|---|---|---|
| R. acetosa | R. acetosella | R. confertus | R. crispus | R. maritimus | R. obtusifolius | R. sanguineus | |
| Flavonoids | |||||||
| Catechin (9.7) | – | – | – | 1.08 ± 0.07 | 0.17 ± 0.01 | 1.32 ± 0.07 | 12.0 ± 0.8 |
| Quercetin 3-O-rutinoside (rutin) (19.3) | 3.4 ± 0.2 | – | 4.3 ± 0.2 | 10.2 ± 0.7 | 9.4 ± 0.6 | 19.0 ± 1.1 | 8.6 ± 0.5 |
| Quercetin 3-β-D-glucoside (isoquercitrin) (19.9) | 0.56 ± 0.03 | – | 20.2 ± 1.3 | 31.9 ± 1.8 | 22.6 ± 1.5 | 54.8 ± 3.5 | 49.5 ± 0.3 |
| Quercetin derivative (16.3)1 | – | – | – | – | 39.6 ± 2.9 | – | – |
| Quercetin derivative (16.9) | – | – | – | – | 27.1 ± 1.5 | – | – |
| Quercetin derivative (18.3) | 2.4 ± 0.2 | – | 0.94 ± 0.06 | – | – | – | – |
| Quercetin derivative (22.73) | 1.31 ± 0.07 | – | – | – | – | – | – |
| Quercetin derivative (23.1) | 2.0 ± 0.1 | – | – | – | – | – | – |
| Quercetin derivative (24.1) | 3.4 ± 0.2 | – | – | – | – | – | – |
| Kaempferol 3-O-glucoside (astragalin) (24.7) | – | – | 1.82 ± 0.09 | 24.4 ± 1.6 | 4.4 ± 0.3 | 5.1 ± 0.3 | 8.6 ± 0.6 |
| Kaempferolderivative (22.8) | – | – | 0.75 ± 0.04 | 12.9 ± 1.0 | 1.4 ± 0.1 | 2.6 ± 0.2 | 3.2 ± 0.2 |
| Kaempferolderivative (20.9) | – | – | – | – | 4.9 ± 0.3 | – | – |
| Luteolin 7-O-glucoside (cynaroside) (20.7) | 0.51 ± 0.03 | 4.3 ± 0.3 | – | – | – | – | – |
| Luteolinderivative (15.5) | – | 89.5 ± 4.7 | – | – | – | – | – |
| Apigeninderivative (19.4) | – | 5.1 ± 0.3 | |||||
| Phenolic acids | |||||||
| Gallic acid (3.8) | – | – | – | 5.3 ± 0.3 | – | 0.34 ± 0.02 | 0.33 ± 0.02 |
| 3,4-Dihydroxybenzoic acid (protocatechuic acid) (5.8) | 0.12 ± 0.01 | 0.58 ± 0.03 | – | 0.56 ± 0.03 | 0.21 ± 0.01 | – | 0.21 ± 0.01 |
| Sinapic acid (8.2) | 4.9 ± 0.4 | 1.22 ± 0.08 | 1.5 ±0.1 | – | 1.8 ± 0.1 | – | – |
| Caftaric acid (9.2) | 1.7 ± 0.1 | – | – | – | – | – | – |
| Chlorogenic acid (10.2) | 1.21 ± 0.09 | 3.04 ± 0.17 | 1.8 ± 0.1 | – | 0.19 ± 0.01 | – | – |
| Caffeic acid (10.5) | – | 0.93 ± 0.05 | – | 0.10 ± 0.01 | 0.29 ± 0.03 | – | – |
| p-Coumaric acid (14.2) | 0.15 ± 0.02 | – | – | – | – | – | – |
| Ellagic acid (17.9) | 0.28 ± 0.02 | – | – | 0.83 ± 0.05 | – | – | – |
| Hydroxybenzoic acid derivative (11.2) | 0.97 ± 0.05 | 4.0 ± 0.2 | – | – | 2.9 ± 0.2 | – | – |
| Hydroxybenzoic acidderivative (12.5) | 0.54 ± 0.05 | 3.0 ± 0.2 | – | – | – | – | – |
| Species | AOA (DPPH) 1, mg TE g–1 | AOA (ABTS), mg TE g–1 | AOA (FRAP), mg TE g–1 |
|---|---|---|---|
| R. acetosa | 3.1 ± 0.3 | 5.1 ± 0.5 | 3.9 ± 0.3 |
| R. acetosella | 31 ± 3 | 48 ± 3 | 27 ± 3 |
| R. confertus | 22 ± 1.3 | 37 ± 3 | 39 ± 4 |
| R. crispus | 69 ± 4 | 56 ± 4 | 57 ± 3 |
| R. maritimus | 31 ± 2 | 63 ± 4 | 61 ± 4 |
| R. obtusifolius | 37 ± 2 | 48 ± 4 | 43 ± 2 |
| R. sanguineus | 35 ± 3 | 52 ± 4 | 47 ± 4 |
| Variables | TPC 1 | TFC | THA | TCC | PAs | TTC | DPPH | ABTS | FRAP |
|---|---|---|---|---|---|---|---|---|---|
| TPC | 1 | 0.881 ** | –0.317 ns | 0.567 * | 0.822 ** | 0.915 ** | 0.806 ** | 0.921 ** | 0.785 ** |
| TFC | 1 | –0.114 ns | 0.368 ns | 0.586 * | 0.664 * | 0.602 * | 0.918 ** | 0.714 ** | |
| THA | 1 | –0.820 ** | –0.768 ** | –0.354 ns | –0.174 ns | –0.322 ns | –0.563 * | ||
| TCC | 1 | 0.809 ** | 0.537 * | 0.389 ns | 0.513 * | 0.614 * | |||
| PAs | 1 | 0.826 ** | 0.721 ** | 0.751 ** | 0.842 ** | ||||
| TTC | 1 | 0.776 ** | 0.701 ** | 0.591 * | |||||
| DPPH | 1 | 0.728 ** | 0.742 ** | ||||||
| ABTS | 1 | 0.909 ** | |||||||
| FRAP | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feduraev, P.; Skrypnik, L.; Nebreeva, S.; Dzhobadze, G.; Vatagina, A.; Kalinina, E.; Pungin, A.; Maslennikov, P.; Riabova, A.; Krol, O.; et al. Variability of Phenolic Compound Accumulation and Antioxidant Activity in Wild Plants of Some Rumex Species (Polygonaceae). Antioxidants 2022, 11, 311. https://doi.org/10.3390/antiox11020311
Feduraev P, Skrypnik L, Nebreeva S, Dzhobadze G, Vatagina A, Kalinina E, Pungin A, Maslennikov P, Riabova A, Krol O, et al. Variability of Phenolic Compound Accumulation and Antioxidant Activity in Wild Plants of Some Rumex Species (Polygonaceae). Antioxidants. 2022; 11(2):311. https://doi.org/10.3390/antiox11020311
Chicago/Turabian StyleFeduraev, Pavel, Liubov Skrypnik, Sofia Nebreeva, Georgii Dzhobadze, Anna Vatagina, Evgeniia Kalinina, Artem Pungin, Pavel Maslennikov, Anastasiia Riabova, Olesya Krol, and et al. 2022. "Variability of Phenolic Compound Accumulation and Antioxidant Activity in Wild Plants of Some Rumex Species (Polygonaceae)" Antioxidants 11, no. 2: 311. https://doi.org/10.3390/antiox11020311
APA StyleFeduraev, P., Skrypnik, L., Nebreeva, S., Dzhobadze, G., Vatagina, A., Kalinina, E., Pungin, A., Maslennikov, P., Riabova, A., Krol, O., & Chupakhina, G. (2022). Variability of Phenolic Compound Accumulation and Antioxidant Activity in Wild Plants of Some Rumex Species (Polygonaceae). Antioxidants, 11(2), 311. https://doi.org/10.3390/antiox11020311