
Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4TM, a Formulation with a High Content of Type-A Proanthocyanidins



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material, Cell Lines, Reagents and Chemicals
2.2. Spectophotometric Determination of Bioactive Compounds
2.2.1. Content of Total Phenolic Compounds
2.2.2. Content of Total Flavonoid Compounds
2.2.3. Content of Total Anthocyanin Compounds
2.2.4. Determination of SP4TM Total PAC Content with the BL-DMAC Method
2.3. Characterization of SP4TM Soluble Proanthocyanidin (PAC) Distribution by Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS)
2.4. Characterization of Other SP4TM Phenolic Compounds by HPLC-DAD-ESI-MS/MS
2.5. Antioxidant Activity of SP4TM on Human HepG2 Cells Using the Light-Up Cell System (LUCS) Method
2.5.1. Neutralization of Intracellular Free Radicals by SP4TM
2.5.2. Inhibition of Plasma Membrane-Based Lipid Peroxidation by SP4TM
2.5.3. H2O2 Quenching (Catalase-like) Activity of SP4TM
2.6. Chemical Antioxidant Capacity of SP4TM by TEAC, FRAP and DPPH Assays
2.7. In Vitro Bacterial Anti-Adhesion Activity (AAA) of SP4TM
2.8. Statistical Analysis
3. Results and Discussion
3.1. SP4TM Contains a High Level of Total Proanthocyanidins and Other Phenolic Compounds
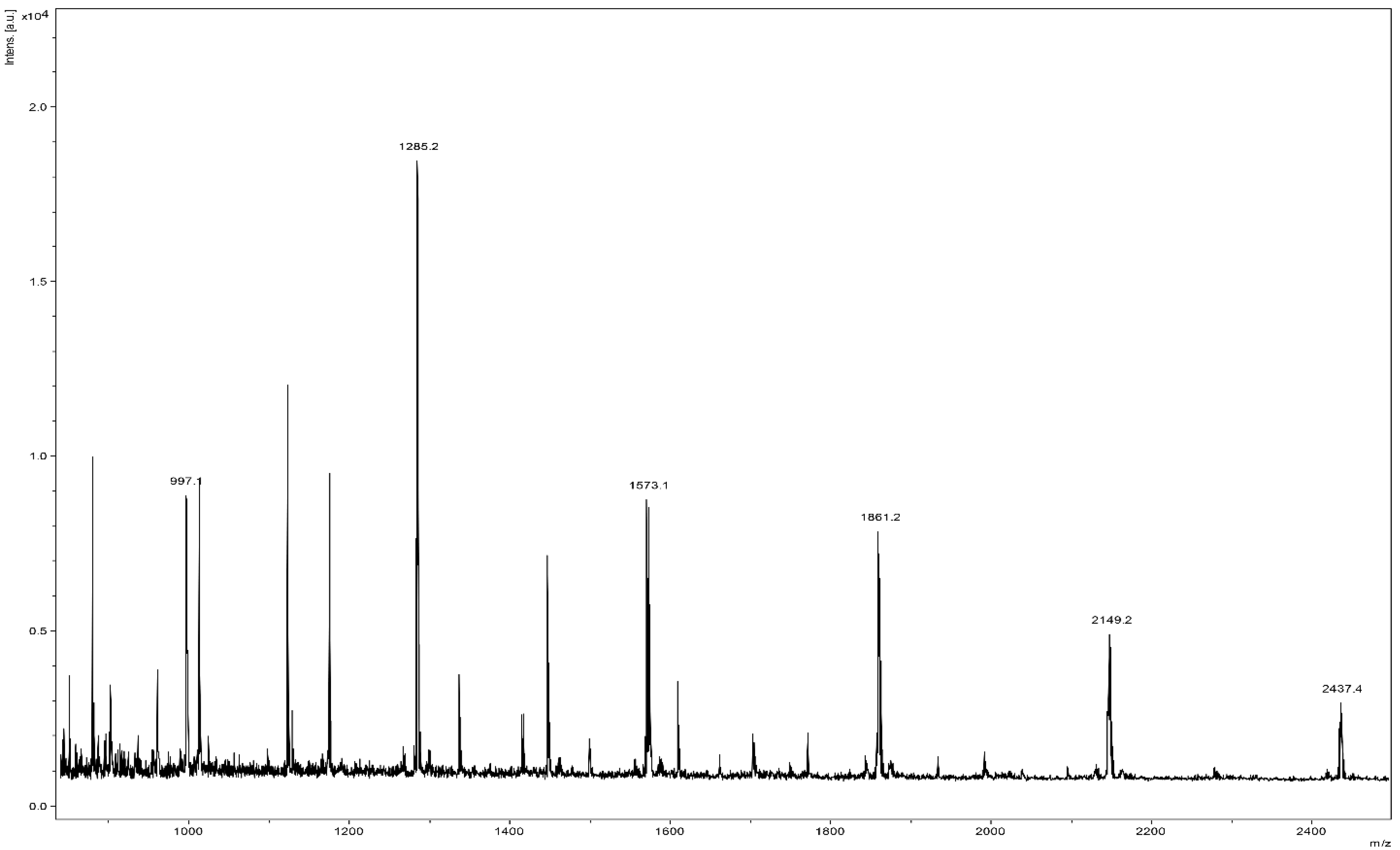
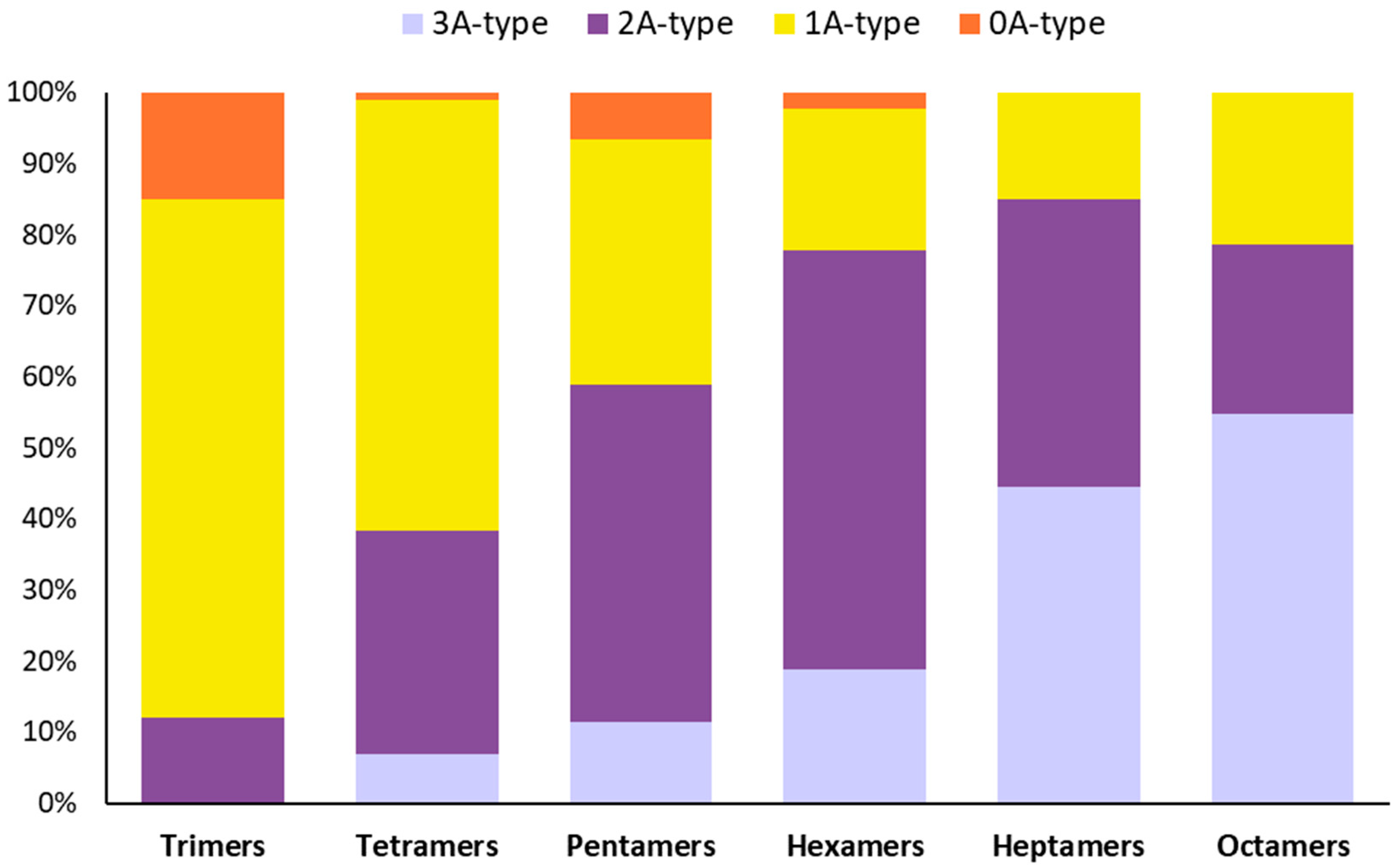
3.2. The Chemical Characterization of SP4TM by MALDI-TOF MS Shows a High Percentage of Type-A- PACs and the Presence of an Oligosaccharide
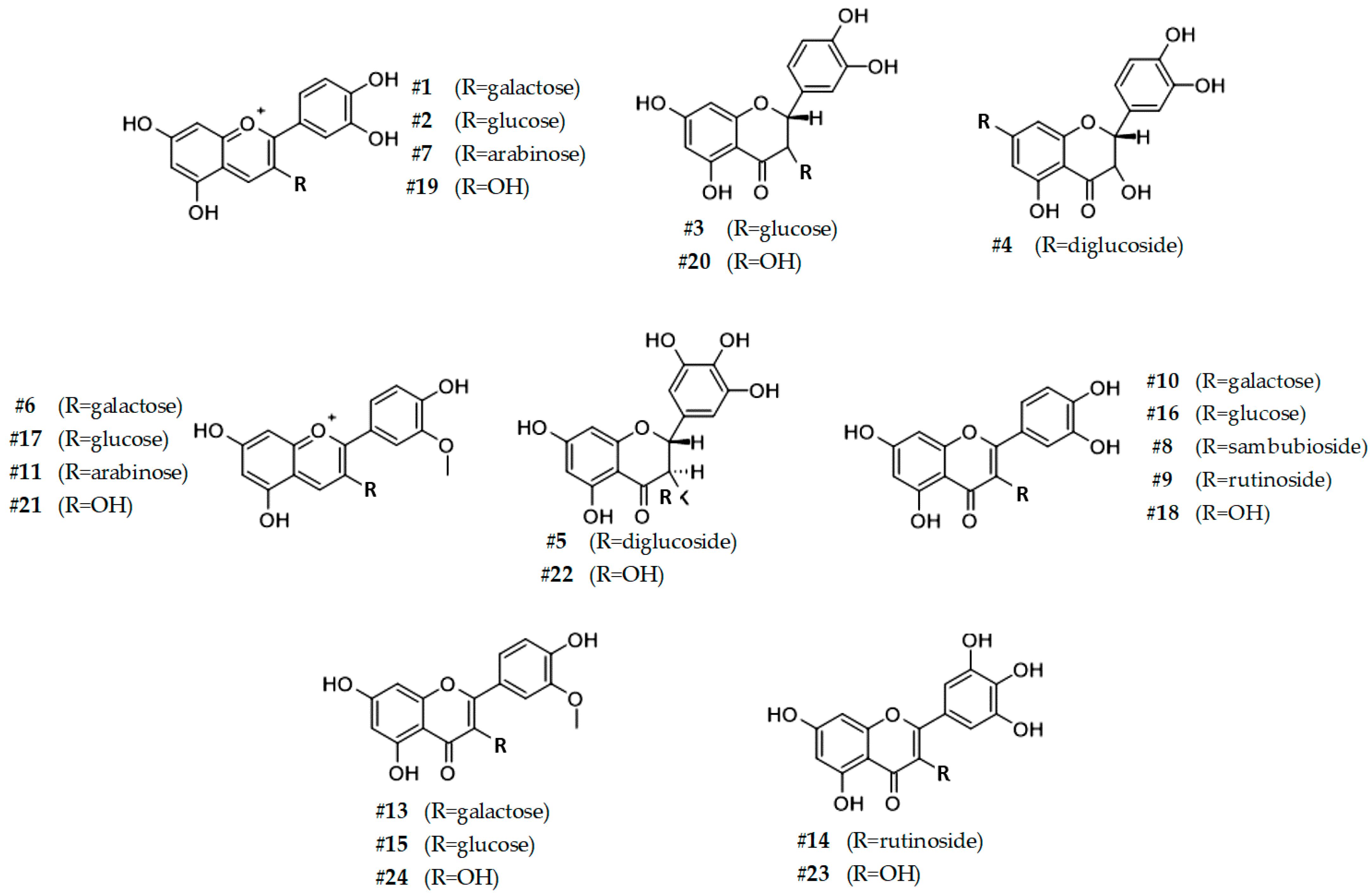
3.3. HPLC-DAD-ESI-MS/MS Analysis of SP4TM Reveals the Presence of Other Phenolic Compounds
3.4. The Antioxidant Power of SP4TM by Cell-Based Methods on HepG2 Human Liver Cell Models Reveals a Strong Intracellular Free Radical Quenching Activity
3.5. Chemical Antioxidant Analysis of SP4TM Confirm Its High Antioxidant Capacity
3.6. SP4TM Shows a Strong In Vitro Bacterial Anti-Adhesion Activity against Uropathogenic P-Fimbriated E. coli
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pennathur, S.; Hecker, L.; Thannickal, V.J. Oxidative stress and cardiovascular fibrosis. In Studies on Cardiovascular Disorders; Sauer, H., Shah, A.M., Laurindo, F.R.M., Eds.; Springer: New York, NY, USA, 2010; pp. 425–441. [Google Scholar]
- Dozor, A.J. The role of oxidative stress in the pathogenesis and treatment of asthma. In Oxidative/Nitrosative Stress and Disease; Laskin, D.L., Ed.; Wiley-Blackwell: Malden, MA, USA, 2010; Volume 1203, pp. 133–137. [Google Scholar]
- Hoeschen, R.J. Oxidative stress and cardiovascular disease. Can. J. Cardiol. 1997, 13, 1021–1025. [Google Scholar] [PubMed]
- Ivan, P.; Anna, J.; Zeno, A.; Istvan, K.; Balazs, N. Psoriasis and oxidative stress. Orv. Hetil. 2016, 157, 1781–1785. [Google Scholar]
- Ward, N.C.; Croft, K.D. Hypertension and oxidative stress. Clin. Exp. Pharmacol. Physiol. 2006, 33, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Haskins, K.; Bradley, B.; Powers, K.; Fadok, V.; Flores, S.; Ling, X.F.; Pugazhenthi, S.; Reusch, J.; Kench, J. Oxidative stress in type 1 diabetes. In Immunology of Diabetes II: Pathogenesis from Mouse to Man; Sanjeevi, C.B., Eisenbarth, G.S., Eds.; New York Acad Sciences: New York, NY, USA, 2003; Volume 1005, pp. 43–54. [Google Scholar]
- Ozturk, Z. Diabetes, oxidative stress and endothelial dysfunction. Bezmialem Sci. 2019, 7, 52–57. [Google Scholar] [CrossRef]
- Vital, P.; Castro, P.; Ittmann, M. Oxidative stress promotes benign prostatic hyperplasia. Prostate 2016, 76, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roumeguere, T.; Sfeir, J.; El Rassy, E.; Albisinni, S.; Van Antwerpen, P.; Boudjeltia, K.Z.; Fares, N.; Kattan, J.; Aoun, F. Oxidative stress and prostatic diseases. Mol. Clin. Oncol. 2017, 7, 723–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottkamp, C.A.; Nunomura, A.; Raina, A.K.; Sayre, L.M.; Perry, G.; Smith, M.A. Oxidative stress, antioxidants, and alzheimer disease. Alzheimer Dis. Assoc. Dis. 2000, 14, S62–S66. [Google Scholar] [CrossRef]
- Chernyak, B.V.; Popova, E.N.; Prikhodko, A.S.; Grebenchikov, O.A.; Zinovkina, L.A.; Zinovkin, R.A. COVID-19 and oxidative stress. Biochemistry 2020, 85, 1543–1553. [Google Scholar] [CrossRef]
- Vicari, E.; La Vignera, S.; Calogero, A.E. Oxidative stress and infection. In Studies on Mens Health and Fertility; Agarwal, A., Aitken, R.J., Alvarez, J.G., Eds.; Springer: New York, NY, USA, 2012; pp. 551–570. [Google Scholar]
- Grant, S.S.; Hung, D.T. Persistent bacterial infections, antibiotic tolerance, and the oxidative stress response. Virulence 2013, 4, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, D.; Mukherjee, M. Reactive oxygen species and uspa overexpession: An alternative bacterial response toward selection and maintenance of multidrug resistance in clinical isolates of uropathogenic E. Coli. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1753–1760. [Google Scholar] [CrossRef]
- Foxman, B. Epidemiology of urinary tract infections: Incidence, morbidity, and economic costs. Am. J. Med. 2002, 113, 5s–13s. [Google Scholar] [CrossRef]
- Aubron, C.; Glodt, J.; Matar, C.; Huet, O.; Borderie, D.; Dobrindt, U.; Duranteau, J.; Denamur, E.; Conti, M.; Bouvet, O. Variation in endogenous oxidative stress in Escherichia Coli natural isolates during growth in urine. BMC Microbiol. 2012, 12, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina-Quiroz, R.C.; Silva-Valenzuela, C.; Brewster, J.; Castro-Nallar, E.; Levy, S.B.; Camilli, A. Cyclic amp regulates bacterial persistence through repression of the oxidative stress response and sos-dependent DNA repair in uropathogenic escherichia coli. mBio 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, L.Y.; Wu, W.Z.; Li, S.J.; Lai, Y.C.; Chen, D.; He, Z.C.; Chang, Z.L.; Xu, P.; Huang, Y.P.; Lei, M.; et al. Escherichia coli aggravates calcium oxalate stone formation via ppk1/flagellin-mediated renal oxidative injury and inflammation. Oxidative Med. Cell. Longev. 2021, 2021, 16. [Google Scholar] [CrossRef]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. Uropathogenic Escherichia coli (upec) infections: Virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef]
- Stahl, W. Oxidative stress—Antioxidants in foods. Ernahr. Umsch. 2011, 58, 536–541. [Google Scholar]
- Urquiaga, I.; Leighton, F. Plant polyphenol antioxidants and oxidative stress. Biol. Res. 2000, 33, 55–64. [Google Scholar] [CrossRef]
- Guo, Q.P.; Li, F.N.; Duan, Y.H.; Wen, C.Y.; Wang, W.L.; Zhang, L.Y.; Huang, R.L.; Yin, Y.L. Oxidative stress, nutritional antioxidants and beyond. Sci. China-Life Sci. 2020, 63, 866–874. [Google Scholar] [CrossRef]
- Mao, X.B.; Gu, C.S.; Chen, D.W.; Yu, B.; He, J. Oxidative stress-induced diseases and tea polyphenols. Oncotarget 2017, 8, 81649–81661. [Google Scholar] [CrossRef] [Green Version]
- Duan, G.L.; Wang, C.N.; Liu, Y.J.; Yu, Q.; Tang, X.L.; Ni, X.; Zhu, X.Y. Resveratrol alleviates endotoxemia-associated adrenal insufficiency by suppressing oxidative/nitrative stress. Endocr. J. 2016, 63, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Elejalde, E.; Villaran, M.C.; Alonso, R.M. Grape polyphenols supplementation for exercise-induced oxidative stress. J. Int. Soc. Sport Nutr. 2021, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Tejada, S.; Nabavi, S.M.; Capo, X.; Martorell, M.; Bibiloni, M.D.; Tur, J.A.; Pons, A.; Sureda, A. Quercetin effects on exercise induced oxidative stress and inflammation. Curr. Org. Chem. 2017, 21, 348–356. [Google Scholar] [CrossRef]
- Hor, S.L.; Teoh, S.L.; Lim, W.L. Plant polyphenols as neuroprotective agents in Parkinson’s disease targeting oxidative stress. Curr. Drug Targets 2020, 21, 458–476. [Google Scholar] [CrossRef]
- Baranowska, M.; Bartoszek, A. Antioxidant and antimicrobial properties of bioactive phytochemicals from cranberry. Postep. Hig. Med. Dosw. 2016, 70, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Yokozawa, T. Modulation of oxidative stress and melanogenesis by proanthocyanidins. Biol. Pharm. Bull. 2009, 32, 1155–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.J.; Weng, Q.N.; Liu, Z.Q.; Shen, M.; Zhang, J.Q.; Wu, W.J.; Liu, H.L. Selection of antioxidants against ovarian oxidative stress in mouse model. J. Biochem. Mol. Toxicol. 2017, 31, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Hu, P.P. Proanthocyanidins prevent ethanol-induced cognitive impairment by suppressing oxidative and inflammatory stress in adult rat brain. Neuroreport 2017, 28, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Jin, W.G.; Shi, H.C. Oligomeric proanthocyanidins protects a549 cells against h2o2-induced oxidative stress via the nrf2-are pathway. Int. J. Mol. Med. 2017, 39, 1548–1554. [Google Scholar] [CrossRef] [Green Version]
- Attia, S.M.; Bakheet, S.A.; Al-Rasheed, N.M. Proanthocyanidins produce significant attenuation of doxorubicin-induced mutagenicity via suppression of oxidative stress. Oxidative Med. Cell. Longev. 2010, 3, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.L.; Geng, X.; Tian, L.L.; Wang, D.B.; Wang, Q. Grape seed proanthocyanidins protect retinal ganglion cells by inhibiting oxidative stress and mitochondrial alteration. Arch. Pharm. Res. 2020, 43, 1056–1066. [Google Scholar] [CrossRef]
- Rajput, S.A.; Zhang, C.; Feng, Y.; Wei, X.T.; Khalil, M.M.; Rajput, I.R.; Baloch, D.M.; Shaukat, A.; Rajput, N.; Qamar, H.; et al. Proanthocyanidins alleviates aflatoxinb(1)-induced oxidative stress and apoptosis through mitochondrial pathway in the bursa of fabricius of broilers. Toxins 2019, 11, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.F.; Chen, L.; Chen, W.B.; Zhao, L.; Lu, Q.; Liu, R. Protective effect of procyanidin a-type dimers against h2o2-induced oxidative stress in prostate du145 cells through the mapks signaling pathway. Life Sci. 2021, 266, 11. [Google Scholar] [CrossRef]
- Mathison, B.D.; Kimble, L.L.; Kaspar, K.L.; Khoo, C.; Chew, B.P. Consumption of cranberry beverage improved endogenous antioxidant status and protected against bacteria adhesion in healthy humans: A randomized controlled trial. Nutr. Res. 2014, 34, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Sureda, A.; Daglia, M.; Izadi, M.; Nabavi, S.M. Cranberry for urinary tract infection: From bench to bedside. Curr. Top. Med. Chem. 2017, 17, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.J.; Wei, Y.; Chen, J.N.; Chen, X.Y.; Zhang, Y. Anti-aging and redox state regulation effects of a-type proanthocyanidins-rich cranberry concentrate and its comparison with grape seed extract in mice. J. Funct. Food. 2017, 30, 63–73. [Google Scholar] [CrossRef]
- Maldonado, P.D.; Rivero-Cruz, I.; Mata, R.; Pedraza-Chaverri, J. Antioxidant activity of a-type proanthocyanidins from Geranium niveum (Geraniaceae). J. Agric. Food Chem. 2005, 53, 1996–2001. [Google Scholar] [CrossRef] [PubMed]
- Martins, G.R.; do Amaral, F.R.L.; Brum, F.L.; Mohana-Borges, R.; de Moura, S.S.T.; Ferreira, F.A.; Sangenito, L.S.; Santos, A.L.S.; Figueiredo, N.G.; da Silva, A.S. Chemical characterization, antioxidant and antimicrobial activities of acai seed (euterpe oleracea mart.) extracts containing a- and b-type procyanidins. LWT-Food Sci. Technol. 2020, 132, 11. [Google Scholar] [CrossRef]
- Huang, B.; Wang, L.; Liu, M.; Wu, X.; Lu, Q.; Liu, R. The underlying mechanism of a-type procyanidins from peanut skin on dss-induced ulcerative colitis mice by regulating gut microbiota and metabolism. J. Food Biochem. 2022, e14103. [Google Scholar] [CrossRef]
- Ishihara, T.; Kaidzu, S.; Kimura, H.; Koyama, Y.; Matsuoka, Y.; Ohira, A. Protective effect of highly polymeric a-type proanthocyanidins from seed shells of japanese horse chestnut (Aesculus turbinata blume) against light-induced oxidative damage in rat retina. Nutrients 2018, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Method Enzym. 1999, 299, 152–178. [Google Scholar]
- Ochieng, B.O.; Anyango, J.O.; Nduko, J.M.; Cheseto, X.; Mudalungu, C.M.; Khamis, F.M.; Ghemoh, C.J.; Egonyu, P.J.; Subramanian, S.; Nakimbugwe, D.; et al. Dynamics in nutrients, sterols and total flavonoid content during processing of the edible long-horned grasshopper (Ruspolia differens serville) for food. Food Chem. 2022, 383, 132397. [Google Scholar] [CrossRef] [PubMed]
- Elisia, I.; Hu, C.; Popovich, D.G.; Kitts, D.D. Antioxidant assessment of an anthocyanin-enriched blackberry extract. Food Chem. 2007, 101, 1052–1058. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Birmingham, A.D.; Esquivel-Alvarado, D.; Maranan, M.; Krueger, C.G.; Reed, J.D. Inter-laboratory validation of 4-(dimethylamino) cinnamaldehyde (dmac) assay using cranberry proanthocyanidin standard for quantification of soluble proanthocyanidins in cranberry foods and dietary supplements, first action official methodsm: 2019.06. J. AOAC Int. 2021, 104, 216–222. [Google Scholar] [CrossRef]
- Feliciano, R.P.; Shea, M.P.; Shanmuganayagam, D.; Krueger, C.G.; Howell, A.B.; Reed, J.D. Comparison of isolated cranberry (Vaccinium macrocarpon ait.) proanthocyanidins to catechin and procyanidins a2 and b2 for use as standards in the 4-(dimethylamino)cinnamaldehyde assay. J. Agric. Food Chem. 2012, 60, 4578–4585. [Google Scholar] [CrossRef]
- Esquivel-Alvarado, D.; Alfaro-Viquez, E.; Krueger, C.G.; Vestling, M.M.; Reed, J.D. Identification of a-type proanthocyanidins in cranberry-based foods and dietary supplements by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry, first action method: 2019.05. J. AOAC Int. 2021, 104, 223–231. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Maffei, M.E. Chemical partitioning and DNA fingerprinting of some pistachio (pistacia vera l.) varieties of different geographical origin. Phytochemistry 2019, 160, 40–47. [Google Scholar] [CrossRef]
- Derick, S.; Gironde, C.; Perio, P.; Reybier, K.; Nepveu, F.; Jauneau, A.; Furger, C. Lucs (light-up cell system), a universal high throughput assay for homeostasis evaluation in live cells. Sci. Rep. 2017, 7, 18069. [Google Scholar] [CrossRef]
- Gliszczyńska-Świgło, A. Antioxidant activity of water soluble vitamins in the teac (trolox equivalent antioxidant capacity) and the frap (ferric reducing antioxidant power) assays. Food Chem. 2006, 96, 131–136. [Google Scholar] [CrossRef]
- Tyrakowska, B.; Soffers, A.E.; Szymusiak, H.; Boeren, S.; Boersma, M.G.; Lemańska, K.; Vervoort, J.; Rietjens, I.M. Teac antioxidant activity of 4-hydroxybenzoates. Free. Radic. Biol. Med. 1999, 27, 1427–1436. [Google Scholar] [CrossRef]
- Kiselova-Kaneva, Y.; Galunska, B.; Nikolova, M.; Dincheva, I.; Badjakov, I. High resolution lc-ms/ms characterization of polyphenolic composition and evaluation of antioxidant activity of sambucus ebulus fruit tea traditionally used in Bulgaria as a functional food. Food Chem. 2022, 367, 130759. [Google Scholar] [CrossRef] [PubMed]
- Foo, L.Y.; Lu, Y.R.; Howell, A.B.; Vorsa, N. The structure of cranberry proanthocyanidins which inhibit adherence of uropathogenic p-fimbriated escherichia coli in vitro. Phytochemistry 2000, 54, 173–181. [Google Scholar] [CrossRef]
- Evans, D.G.; Evans, D.J., Jr.; Tjoa, W. Hemagglutination of human group a erythrocytes by enterotoxigenic escherichia coli isolated from adults with diarrhea: Correlation with colonization factor. Infect. Immun. 1977, 18, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Man, P.; Cedergren, B.; Enerbäck, S.; Larsson, A.C.; Leffler, H.; Lundell, A.L.; Nilsson, B.; Svanborg-Edén, C. Receptor-specific agglutination tests for detection of bacteria that bind globoseries glycolipids. J. Clin. Microbiol. 1987, 25, 401–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Jiménez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the phenol-explorer database. Eur. J. Clin. Nutr. 2010, 64, S112–S120. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Trandafir, I.; Nour, V. Phenolic acids and flavonoids profiles of extracts from edible wild fruits and their antioxidant properties. Int. J. Food Prop. 2017, 20, 3124–3134. [Google Scholar] [CrossRef] [Green Version]
- Shraim, A.M.; Ahmed, T.A.; Rahman, M.M.; Hijji, Y.M. Determination of total flavonoid content by aluminum chloride assay: A critical evaluation. LWT 2021, 150, 111932. [Google Scholar] [CrossRef]
- Taghavi, T.; Patel, H.; Rafie, R. Comparing ph differential and methanol-based methods for anthocyanin assessments of strawberries. Food Sci. Nutr. 2021, 1–9. [Google Scholar] [CrossRef]
- Mannino, G.; Chinigò, G.; Serio, G.; Genova, T.; Gentile, C.; Munaron, L.; Bertea, C.M. Proanthocyanidins and where to find them: A meta-analytic approach to investigate their chemistry, biosynthesis, distribution, and effect on human health. Antioxidants 2021, 10, 1229. [Google Scholar] [CrossRef]
- Krueger, C.G.; Reed, J.D.; Feliciano, R.P.; Howell, A.B. Quantifying and characterizing proanthocyanidins in cranberries in relation to urinary tract health. Anal. Bioanal. Chem. 2013, 405, 4385–4395. [Google Scholar] [CrossRef]
- Reed, J.D.; Krueger, C.G.; Vestling, M.M. Maldi-tof mass spectrometry of oligomeric food polyphenols. Phytochemistry 2005, 66, 2248–2263. [Google Scholar] [CrossRef]
- Rue, E.A.; Glinski, J.A.; Glinski, V.B.; van Breemen, R.B. Ion mobility-mass spectrometry for the separation and analysis of procyanidins. J. Mass Spectrom. 2020, 55, e4377. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhou, K.; Jiang, H.Q.; Wen, L.K.; He, Y.; Lu, S.Y.; Wang, B.X.; Li, J. Current advances in anthocyanins: Structure, bioactivity and human health. J. Food Nutr. Res. 2021, 60, 203–216. [Google Scholar]
- Das, A.; Baidya, R.; Chakraborty, T.; Samanta, A.K.; Roy, S. Pharmacological basis and new insights of taxifolin: A comprehensive review. Biomed. Pharmacother. 2021, 142, 112004. [Google Scholar] [CrossRef]
- Ribeiro, D.; Fernandes, E.; Freitas, M. Chapter 20—Flavonoids as modulators of neutrophils’ oxidative burst: Structure-activity relationship. In Polyphenols: Mechanisms of Action in Human Health and Disease, 2nd ed.; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: London, UK, 2018; pp. 261–276. [Google Scholar]
- Sato, M.; Murakami, K.; Uno, M.; Nakagawa, Y.; Katayama, S.; Akagi, K.-I.; Masuda, Y.; Takegoshi, K.; Irie, K. Site-specific inhibitory mechanism for amyloid β42 aggregation by catechol-type flavonoids targeting the Lys residues. J. Biol. Chem. 2013, 288, 23212–23224. [Google Scholar] [CrossRef] [Green Version]
- Furger, C. Live cell assays for the assessment of antioxidant activities of plant extracts. Antioxidants 2021, 10, 944. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (caa) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef]
- Gironde, C.; Rigal, M.; Dufour, C.; Furger, C. Aop1, a new live cell assay for the direct and quantitative measure of intracellular antioxidant effects. Antioxidants 2020, 9, 471. [Google Scholar] [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of methods to determine antioxidant capacities. Food Anal. Methods 2009, 2, 41–60. [Google Scholar] [CrossRef]
- Bolanos de la Torre, A.A.S.; Henderson, T.; Nigam, P.S.; Owusu-Apenten, R.K. A universally calibrated microplate ferric reducing antioxidant power (frap) assay for foods and applications to manuka honey. Food Chem. 2015, 174, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of abts, dpph, frap, and orac assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Mantzorou, M.; Giaginis, C. Cranberry consumption against urinary tract infections: Clinical stateof- the-art and future perspectives. Curr. Pharm. Biotechnol. 2018, 19, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Howell, A.B.; Zhang, D.J.; Khoo, C. A randomized, double-blind, placebo-controlled pilot study to assess bacterial anti-adhesive activity in human urine following consumption of a cranberry supplement. Food Funct. 2019, 10, 7645–7652. [Google Scholar] [CrossRef] [Green Version]
- Howell, A.; Souza, D.; Roller, M.; Fromentin, E. Comparison of the anti-adhesion activity of three different cranberry extracts on uropathogenic p-fimbriated Escherichia coli: A randomized, double-blind, placebo controlled, ex vivo, acute study. Nat. Prod. Commun. 2015, 10, 1215–1218. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, K.L.; Howell, A.B.; Khoo, C. A randomized, double-blind, placebo-controlled trial to assess the bacterial anti-adhesion effects of cranberry extract beverages. Food Funct. 2015, 6, 1212–1217. [Google Scholar] [CrossRef]
- Hendrich, A.B.; Strugala, P.; Dudra, A.; Kucharska, A.Z.; Sokol-Letowska, A.; Wojnicz, D.; Cisowska, A.; Sroka, Z.; Gabrielska, J. Microbiological, antioxidant and lipoxygenase-1 inhibitory activities of fruit extracts of chosen rosaceae family species. Adv. Clin. Exp. Med. 2020, 29, 215–224. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Phenolics | Method Used | Content |
|---|---|---|
| TPC | Folin–Ciocalteu assay | 885.51 ± 14.19 mg GAE g−1 d.wt |
| TFC | Aluminum Chloride assay | 135.52 ± 8.98 mg RE g−1 d.wt |
| TAC | pH differential method | 54.84 ± 2.97 mg CE g−1 d.wt |
| TPAC | DMAC assay | 379.43 ± 12.44 mg PAC-A2 type g−1 d.wt |
| 737.54 ± 8.12 mg cPAC g−1 d.wt |
| Degree of Polymerization | Type-A to Type-B Interflavan Bonds | Relative Percentage (%) |
|---|---|---|
| Trimers | 2Type-A:0Type-B | 12.10 |
| 1Type-A:1Type-B | 72.83 | |
| 0Type-A:2Type-B | 15.07 | |
| Tetramers | 3Type-A:0Type-B | 6.91 |
| 2Type-A:1Type-B | 31.37 | |
| 1Type-A:2Type-B | 60.70 | |
| 0Type-A:3Type-B | 1.02 | |
| Pentamers | 4Type-A:0Type-B | 0.00 |
| 3Type-A:1Type-B | 11.38 | |
| 2Type-A:2Type-B | 47.41 | |
| 1Type-A:3Type-B | 34.59 | |
| 0Type-A:4Type-B | 6.62 | |
| Hexamers | 4Type-A: 1Type-B | 0.00 |
| 3Type-A: 2Type-B | 18.88 | |
| 2Type-A: 3Type-B | 58.81 | |
| 1Type-A: 4Type-B | 19.98 | |
| 0Type-A: 5Type-B | 2.33 | |
| Heptamers | 4Type-A: 2Type-B | 0.00 |
| 3Type-A: 3Type-B | 45.93 | |
| 2Type-A: 4Type-B | 41.86 | |
| 1Type-A: 5Type-B | 15.50 | |
| 0Type-A: 6Type-B | 0.00 | |
| Octamers | 4Type-A: 2Type-B | 0.00 |
| 3Type-A: 3Type-B | 57.29 | |
| 2Type-A: 4Type-B | 25.05 | |
| 1Type-A: 5Type-B | 22.33 | |
| 0Type-A: 6Type-B | 0.00 |
| # | RT | m/z | MS/MS | λ | Compound | |||
|---|---|---|---|---|---|---|---|---|
| Common Name | Chemical Name | CAS-ID | mg g−1 d.wt | |||||
| 1 | 22.6 | 449 | 286.9 | 520 | Idaein | Cyanidin 3-O-galactoside | 27661-36-5 | 0.52 ± 0.01 m |
| 2 | 24.6 | 449 | 286.9 | 520 | Chrysanthemin | Cyanidin 3-O-glucoside | 7084-24-4 | 4.14 ± 0.12 f |
| 3 | 24.9 | 465 | 436.7; 302.7 | 260, 280 | Taxifolin-glucoside | Dihydroquercetin-3-O-glucoside | 27297-45-6 | 0.12 ± 0.01 op |
| 4 | 25.0 | 611 | 574.7; 422.7 | 260, 280 | Dihydroneohesperidin | Dihydroquercetin-7-O-diglucoside | 13241-33-3 | 2.06 ± 0.06 h |
| 5 | 25.1 | 643 | 596.6; 490.6; 318.7 | 260, 280 | Dihydromyricetin 3,4’-O-diglucoside | 0.28 ± 0.01 n | ||
| 6 | 26.4 | 463 | 301 | 520 | Peonidin 3-O-galactoside | 28148-89-2 | 1.95 ± 0.06 h | |
| 7 | 26.8 | 419 | 286.8 | 520 | Cyanidin 3-O-arabinoside | 57186-11-5 | 0.09 ± 0.01 op | |
| 8 | 26.9 | 597 | 548.9; 301.0 | 260, 280 | Quercetin 3-O-sambubioside | 83048-35-5 | 0.01 ± 0.01 p | |
| 9 | 27.5 | 609 | 342.7; 300.7 | 260, 280 | Rutin | Quercetin 3-O-rutinoside | 153-18-4 | 0.96 ± 0.02 l |
| 10 | 28.7 | 463 | 416.7; 300.7 | 260, 280 | Hyperoside | Quercetin 3-O-galactoside | 482-36-0 | 0.18 ± 0.01 no |
| 11 | 29.1 | 473 | 455.1, 427.0, 300.9 | 520 | Peonidin 3-O-arabinoside | 524943-91-7 | 0.55 ± 0.02 m | |
| 12 | 29.2 | 477 | 300.7 | 260, 280 | Miquelianin | Quercetin 3-O-glucuronide | 22688-79-5 | 0.08 ± 0.01 op |
| 13 | 30.2 | 623 | 314.7; 299.7; 270.7 | 260, 280 | Isorhamnetin-galactoside | 3′-Methylquercetin 3-O-galactoside | 6743-92-6 | 0.43 ± 0.01 m |
| 14 | 30.2 | 625 | 317.1 | 260, 280 | Myricetin 3-O-rutinoside | 41093-68-9 | 0.15 ± 0.01 op | |
| 15 | 31.0 | 477 | 356.7; 314.7 | 260, 280 | Isorhamnetin-glucoside | 3′-Methylquercetin 3-O-glucoside | 5041-82-7 | 0.15 ± 0.01 op |
| 16 | 31.9 | 463 | 416.7; 300.7 | 260, 280 | Isoquercitrin | Quercetin 3-O-glucoside | 482-35-9 | 1.47 ± 0.04 i |
| 17 | 32.5 | 463 | 300.9 | 520 | Oxycoccicyanin | Peonidin 3-O-glucoside | 68795-37-9 | 0.29 ± 0.0 1n |
| 18 | 35.5 | 301 | 256.8 | 260, 280 | Quercetin | 117-39-5 | 11.29 ± 0.43 e | |
| 19 | 36.1 | 287 | 258.9; 230.9 | 520 | Cyanidin | 13306-05-3 | 37.21 ± 1.29 b | |
| 20 | 38.5 | 303 | 284.7 | 260.28 | Taxifolin | Dihydroquercetin | 480-18-2 | 24.08 ± 0.88 d |
| 21 | 48.0 | 301 | 240.1 | 520 | Peonidin | 134-01-0 | 2.21 ± 0.07 g | |
| 22 | 48.4 | 318 | 295.4 | 260, 280 | Ampelopsin | Dihydromyricetin | 27200-12-0 | 74.23 ± 2.94 a |
| 23 | 51.9 | 317 | 297.9; 270.7 | 260, 280 | Myricetin | 529-44-2 | 30.36 ± 0.93 c | |
| 24 | 52.1 | 315 | 297 | 260, 280 | Isorhamnetin | 3′-Methylquercetin | 480-19-3 | 11.55 ± 0.35 e |
| Antioxidant Assay | EC10 | EC50 | EC90 |
|---|---|---|---|
| Neutralization of intracellular free radicals | 7.18 a μg/ mL (95% CI: 3.36–12.13 μg/mL) | 30.11 a μg/ mL (95% CI: 23.23–39.08 μg/mL) | 126.30 a μg/ mL (95% CI: 74.62–267.7 μg/mL) |
| Inhibition of plasma membrane-based lipid peroxidation | 2.26 b mg/ mL (95% CI: 0.021–238.3) | 7.81 b mg/ mL (95% CI: 1.105–55.2) | 27.02 b mg/ mL (95% CI: 0.058–12548) |
| H2O2 quenching (catalase-like) activity | 7.17 a μg/ mL (95% CI: ND) | 596.00 c μg/ mL (95% CI: ND) | 49.53 c mg/ mL (95% CI: ND) |
| Assay | µmol TE/g | IC50 |
|---|---|---|
| ABTS | 139.92 ± 6.16 | 2.85 ± 0.03 |
| DPPH | 89.51 ± 3.91 | 4.46 ± 0.12 |
| FRAP | 2643.28 ± 39.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, G.; Maffei, M.E. Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4TM, a Formulation with a High Content of Type-A Proanthocyanidins. Antioxidants 2022, 11, 1234. https://doi.org/10.3390/antiox11071234
Mannino G, Maffei ME. Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4TM, a Formulation with a High Content of Type-A Proanthocyanidins. Antioxidants. 2022; 11(7):1234. https://doi.org/10.3390/antiox11071234
Chicago/Turabian StyleMannino, Giuseppe, and Massimo E. Maffei. 2022. "Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4TM, a Formulation with a High Content of Type-A Proanthocyanidins" Antioxidants 11, no. 7: 1234. https://doi.org/10.3390/antiox11071234