
TRPV4 Role in Neuropathic Pain Mechanisms in Rodents


Abstract
:1. Neuropathic Pain
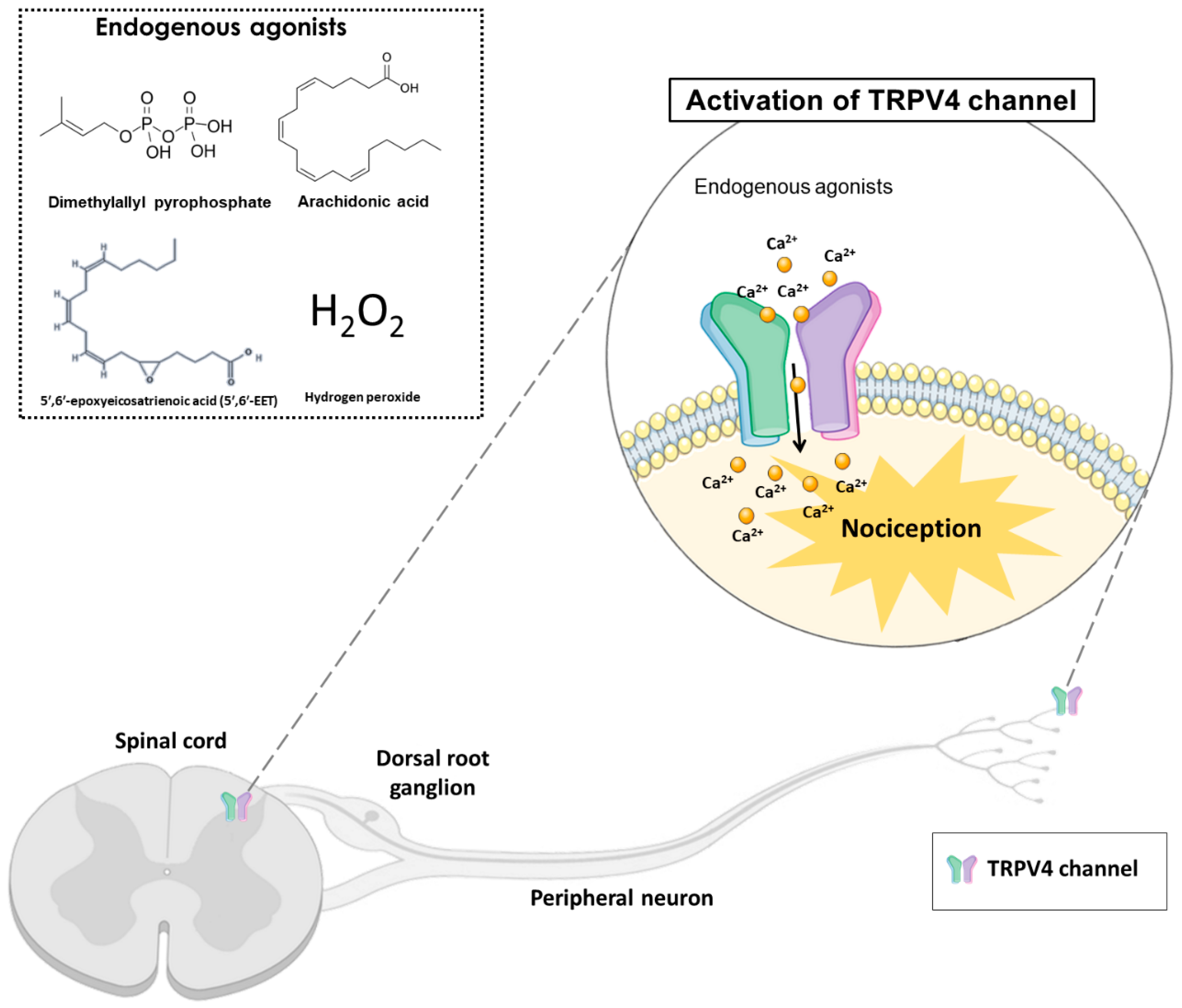
2. Transient Receptor Potential (TRP)
Transient Receptor Potential Vanilloid 4 (TRPV4)
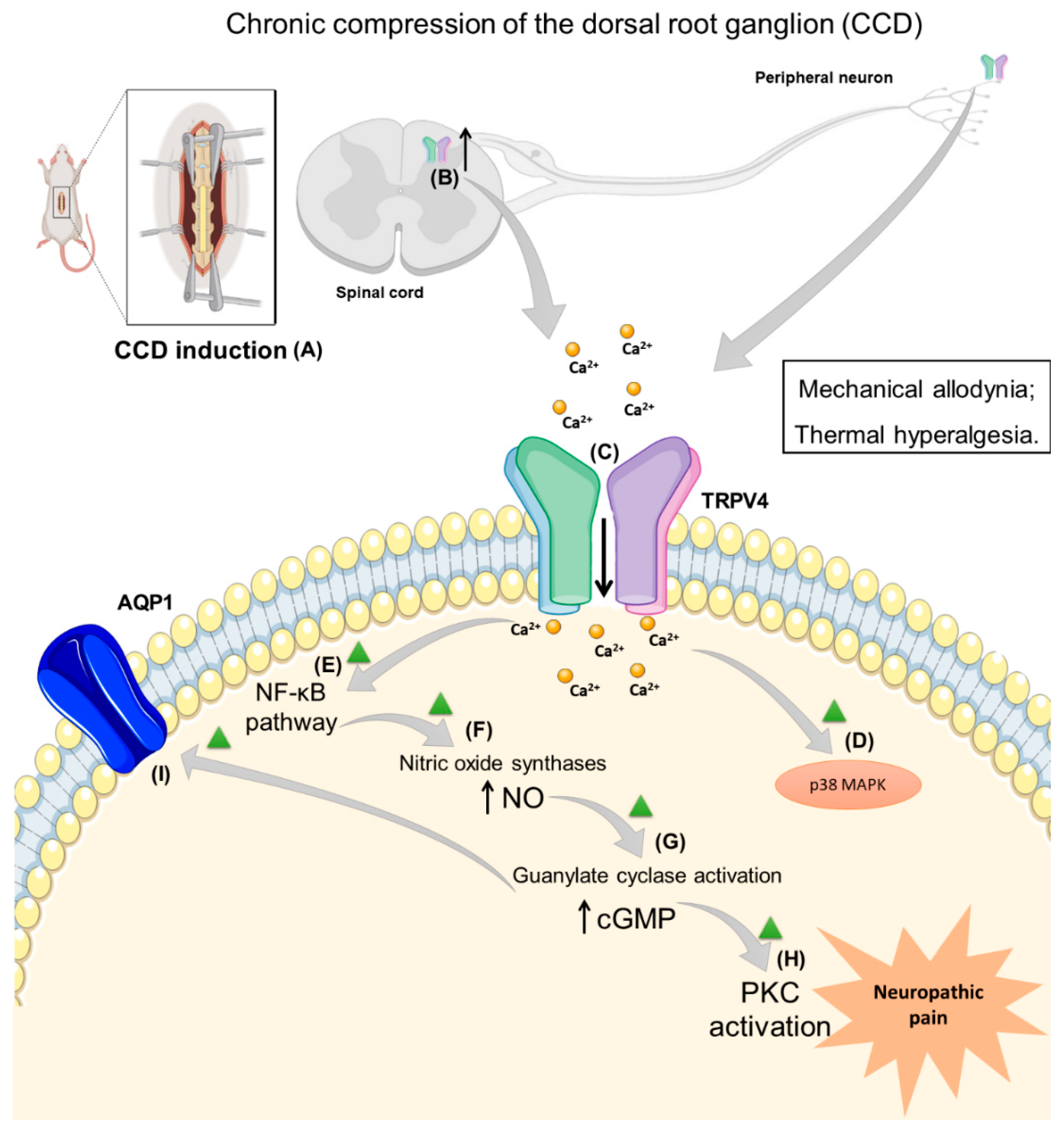
3. Neuropathic Pain Induced by Trauma and Surgery and TRPV4 Role
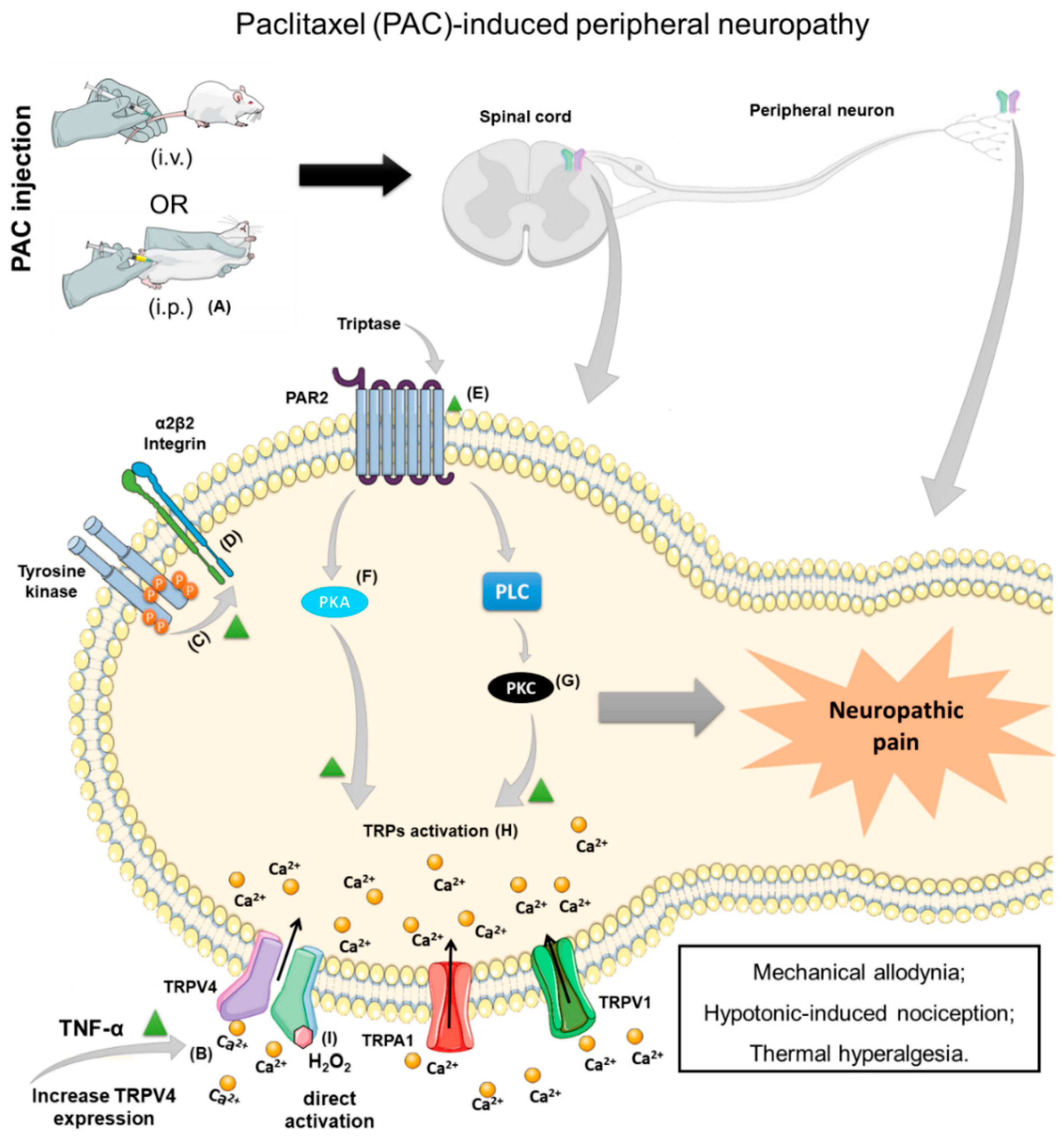
4. Chemotherapy-Induced Neuropathic Pain (CINP) and TRPV4 Participation
5. Other Neuropathic Pain Models with TRPV4 Involvement
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 20,30-Dideoxycytidine | ddC |
| 5′,6′-Epoxyeicosatrienoic acid | 5′,6′-EET |
| 4α-Phorbol 12,13-didecanoate | 4α-PDD |
| Botulinum toxin type A | BoNT-A |
| Calcitonin gene-related peptide | CGRP |
| Chronic compression of the dorsal root ganglion | CCD |
| Cyclic adenosine monophosphate | cAMP |
| Cyclic guanosine monophosphate | cGMP |
| Chemotherapy-induced neuropathic pain | CINP |
| Dorsal root ganglion | DRG |
| Folding recognition domain | FRD |
| Herpes simplex virus | HSV |
| Hydrogen peroxide | H2O2 |
| Human immunodeficiency virus | HIV |
| Immunohistochemistry | IHC |
| Infraorbital nerve injury | IONI |
| Inhibitory-kappa B | I-κB |
| Insulin growth factor 1 | IGF-1 |
| Interleukin-1β | IL-1β |
| Interleukin-6 | IL-6 |
| Interleukin-8 | IL-8 |
| International Association for the Study of Pain | IASP |
| International Classification of Diseases | ICD-11 |
| Intragastric | i.g. |
| Intraperitoneal | i.p. |
| Intravenously | i.v. |
| Lentiviral vector negative control | LV-NC |
| Mitogen-activated protein kinases | MAPK |
| Nitric oxide | NO |
| Nitric oxide synthase | NOS |
| Nuclear factors kappa B | NF-κB |
| Phospholipase C | PLC |
| Pyrrolidine dithiocarbamate | PDTC |
| Protein kinase C | PKC |
| Protease activating receptor 2 | PAR2 |
| Protein kinase A | PKA |
| Protein kinase G | PKG |
| Selective serotonin and norepinephrine reuptake inhibitor | SSNRI |
| Spinal cord injury | SCI |
| Subcutaneous | s.c. |
| Transient receptor potential ankyrin | TRPA |
| Transient receptor potential canonical | TRPC |
| Transient receptor potential melastatin | TRPM |
| Transient receptor potential mucolipin | TRPML |
| Transient receptor potential polycystin | TRPP |
| Transient receptor potential vanilloid | TRPV |
| TRPV4 knockout | Trpv4−/− |
| Toll-like receptor 4 | TLR-4 |
| Tumor necrosis factor-α | TNF-α. |
| Ultraviolet B-rays | UVB |
References
- Treede, R.-D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. Chronic pain as a symptom or a disease: The IASP Classification of Chronic Pain for the International Classification of Diseases (ICD-11). Pain 2019, 160, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, J.; Finnerup, N.B.; Attal, N.; Aziz, Q.; Baron, R.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Cruccu, G.; Davis, K.D.; et al. The IASP classification of chronic pain for ICD-11: Chronic neuropathic pain. Pain 2019, 160, 53–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Prim. 2017, 3, 17002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnerup, N.B.; Kuner, R.; Jensen, T.S. Neuropathic Pain: From Mechanisms to Treatment. Physiol. Rev. 2021, 101, 259–301. [Google Scholar] [CrossRef]
- Yamasaki, H.; Funai, Y.; Funao, T.; Mori, T.; Nishikawa, K. Effects of Tramadol on Substantia Gelatinosa Neurons in the Rat Spinal Cord: An In Vivo Patch-Clamp Analysis. PLoS ONE 2015, 10, e0125147. [Google Scholar] [CrossRef]
- Dalenogare, D.P.; Theisen, M.C.; Peres, D.S.; Fialho, M.F.P.; Luckemeyer, D.D.; Antoniazzi, C.T.D.; Kudsi, S.Q.; Ferreira, M.A.; Ritter, C.D.S.; Ferreira, J.; et al. TRPA1 activation mediates nociception behaviors in a mouse model of relapsing-remitting experimental autoimmune encephalomyelitis. Exp. Neurol. 2020, 328, 113241. [Google Scholar] [CrossRef]
- Koivisto, A.P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: From target validation to clinical studies. Nat. Rev. Drug Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef]
- Iannone, L.F.; De Logu, F.; Geppetti, P.; De Cesaris, F. The role of TRP ion channels in migraine and headache. Neurosci. Lett. 2022, 768, 136380. [Google Scholar] [CrossRef]
- Gu, Q.; Lee, L.Y. TRP channels in airway sensory nerves. Neurosci. Lett. 2021, 748, 135719. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef]
- Moran, M.M. TRP Channels as Potential Drug Targets. Annu. Rev. Pharm. Toxicol. 2018, 58, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A. Transient Receptor Potential (TRP) Channels in Health and Disease. Cells 2019, 8, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pumroy, R.A.; Fluck, E.C., 3rd; Ahmed, T.; Moiseenkova-Bell, V.Y. Structural insights into the gating mechanisms of TRPV channels. Cell Calcium 2020, 87, 102168. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.; Campbell, J.N.; Chung, M.K. Fight fire with fire: Neurobiology of capsaicin-induced analgesia for chronic pain. Pharmacol. Ther. 2021, 220, 107743. [Google Scholar] [CrossRef]
- Seebohm, G.; Schreiber, J.A. Beyond Hot and Spicy: TRPV Channels and their Pharmacological Modulation. Cell Physiol. Biochem. 2021, 55, 108–130. [Google Scholar] [CrossRef]
- Taga, A.; Peyton, M.A.; Goretzki, B.; Gallagher, T.Q.; Ritter, A.; Harper, A.; Crawford, T.O.; Hellmich, U.A.; Sumner, C.J.; McCray, B.A. TRPV4 mutations causing mixed neuropathy and skeletal phenotypes result in severe gain of function. Ann. Clin. Transl. Neurol. 2022, 9, 375–391. [Google Scholar] [CrossRef]
- Ji, C.; McCulloch, C.A. TRPV4 integrates matrix mechanosensing with Ca(2+) signaling to regulate extracellular matrix remodeling. FEBS J. 2021, 288, 5867–5887. [Google Scholar] [CrossRef]
- Yu, S.; Huang, S.; Ding, Y.; Wang, W.; Wang, A.; Lu, Y. Transient receptor potential ion-channel subfamily V member 4: A potential target for cancer treatment. Cell Death Dis. 2019, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Paknejad, N.; Maksaev, G.; Sala-Rabanal, M.; Nichols, C.G.; Hite, R.K.; Yuan, P. Cryo-EM and X-ray structures of TRPV4 reveal insight into ion permeation and gating mechanisms. Nat. Struct. Mol. Biol. 2018, 25, 252–260. [Google Scholar] [CrossRef]
- Kumar, H.; Lee, S.-H.; Kim, K.-T.; Zeng, X.; Han, I. TRPV4: A Sensor for Homeostasis and Pathological Events in the CNS. Mol. Neurobiol. 2018, 55, 8695–8708. [Google Scholar] [CrossRef]
- Nguyen, T.N.; Siddiqui, G.; Veldhuis, N.A.; Poole, D.P. Diverse Roles of TRPV4 in Macrophages: A Need for Unbiased Profiling. Front. Immunol. 2021, 12, 828115. [Google Scholar] [CrossRef] [PubMed]
- Lawhorn, B.G.; Brnardic, E.J.; Behm, D.J. TRPV4 antagonists: A patent review (2015–2020). Expert Opin. Ther. Patents 2021, 31, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.; Bicca, M.A.; Manjavachi, M.N.; Segat, G.C.; Dias, F.C.; Fernandes, E.S.; Calixto, J.B. Kinin Receptors Sensitize TRPV4 Channel and Induce Mechanical Hyperalgesia: Relevance to Paclitaxel-Induced Peripheral Neuropathy in Mice. Mol. Neurobiol. 2018, 55, 2150–2161. [Google Scholar] [CrossRef] [PubMed]
- Gamper, N.; Ooi, L. Redox and nitric oxide-mediated regulation of sensory neuron ion channel function. Antioxid. Redox Signal. 2015, 22, 486–504. [Google Scholar] [CrossRef]
- Sandireddy, R.; Yerra, V.G.; Areti, A.; Komirishetty, P.; Kumar, A. Neuroinflammation and Oxidative Stress in Diabetic Neuropathy: Futuristic Strategies Based on These Targets. Int. J. Endocrinol. 2014, 2014, 674987. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Hu, X.; Liang, F.; Liu, J.; Zhou, H.; Liu, J.; Wang, H.; Tang, H. Therapeutic effects of moxibustion simultaneously targeting Nrf2 and NF-κB in diabetic peripheral neuropathy. Appl. Biochem. Biotechnol. 2019, 189, 1167–1182. [Google Scholar] [CrossRef] [Green Version]
- Suo, J.; Wang, M.; Zhang, P.; Lu, Y.; Xu, R.; Zhang, L.; Qiu, S.; Zhang, Q.; Qian, Y.; Meng, J.; et al. Siwei Jianbu decoction improves painful paclitaxel-induced peripheral neuropathy in mouse model by modulating the NF-κB and MAPK signaling pathways. Regen. Med. Res. 2020, 8, 2. [Google Scholar] [CrossRef]
- Lim, H.; Lee, H.; Noh, K.; Lee, S.J. IKK/NF-kappaB-dependent satellite glia activation induces spinal cord microglia activation and neuropathic pain after nerve injury. Pain 2017, 158, 1666–1677. [Google Scholar] [CrossRef]
- Facer, P.; Casula, M.A.; Smith, G.D.; Benham, C.D.; Chessell, I.P.; Bountra, C.; Sinisi, M.; Birch, R.; Anand, P. Differential expression of the capsaicin receptor TRPV1 and related novel receptors TRPV3, TRPV4 and TRPM8 in normal human tissues and changes in traumatic and diabetic neuropathy. BMC Neurol. 2007, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Grace, M.S.; Bonvini, S.J.; Belvisi, M.G.; McIntyre, P. Modulation of the TRPV4 ion channel as a therapeutic target for disease. Pharmacol. Ther. 2017, 177, 9–22. [Google Scholar] [CrossRef]
- McCray, B.A.; Diehl, E.; Sullivan, J.M.; Aisenberg, W.H.; Zaccor, N.W.; Lau, A.R.; Rich, D.J.; Goretzki, B.; Hellmich, U.A.; Lloyd, T.E.; et al. Neuropathy-causing TRPV4 mutations disrupt TRPV4-RhoA interactions and impair neurite extension. Nat. Commun. 2021, 12, 1444. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado, J.D.D.; Popovich, P.G.; Partida-Sanchez, S.; Guerau-De-Arellano, M. Novel Markers to Delineate Murine M1 and M2 Macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maqboul, A.; Elsadek, B. Expression profiles of TRPV1, TRPV4, TLR4 and ERK1/2 in the dorsal root ganglionic neurons of a cancer-induced neuropathy rat model. PeerJ 2018, 6, e4622. [Google Scholar] [CrossRef] [Green Version]
- Dias, F.C.; Alves, V.S.; Matias, D.O.; Figueiredo, C.P.; Miranda, A.L.P.; Passos, G.F.; Costa, R. The selective TRPV4 channel antagonist HC-067047 attenuates mechanical allodynia in diabetic mice. Eur. J. Pharm. 2019, 856, 172408. [Google Scholar] [CrossRef]
- Sanchez, J.C.; Munoz, L.V.; Ehrlich, B.E. Modulating TRPV4 channels with paclitaxel and lithium. Cell Calcium 2020, 91, 102266. [Google Scholar] [CrossRef]
- Liu, W.; Pu, B.; Liu, M.; Zhang, X.; Zeng, R. Down-regulation of MAPK pathway alleviates TRPV4-mediated trigeminal neuralgia by inhibiting the activation of histone acetylation. Exp. Brain Res. 2021, 239, 3397–3404. [Google Scholar] [CrossRef] [PubMed]
- Attal, N. Spinal cord injury pain. Rev. Neurol. 2021, 177, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Song, X.J.; Hu, S.J.; Greenquist, K.W.; Zhang, J.M.; LaMotte, R.H. Mechanical and thermal hyperalgesia and ectopic neuronal discharge after chronic compression of dorsal root ganglia. J. Neurophysiol. 1999, 82, 3347–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, Y.; Kawamoto, H.; Nakanishi, O. Characterization of heat-hyperalgesia in an experimental trigeminal neuropathy in rats. Exp. Brain Res. 1997, 116, 97–103. [Google Scholar] [CrossRef]
- Sommer, C. Neuropathic Pain Model, Chronic Constriction Injury. In Encyclopedia of Pain; Schmidt, R.F., Willis, W.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1290–1292. [Google Scholar] [CrossRef]
- Ropper, A.E.; Neal, M.T.; Theodore, N. Acute management of traumatic cervical spinal cord injury. Pract. Neurol. 2015, 15, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Challa, S.R. Surgical animal models of neuropathic pain: Pros and Cons. Int. J. Neurosci. 2015, 125, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.J.; Xing, J.L. An experimental model for chronic compression of dorsal root ganglion produced by intervertebral foramen stenosis in the rat. Pain 1998, 77, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Song, K.-S.; Cho, J.H.; Hong, J.-Y.; Lee, J.H.; Kang, H.; Ham, D.-W.; Ryu, H.-J. Neuropathic Pain Related with Spinal Disorders: A Systematic Review. Asian Spine J. 2017, 11, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.-Y.; Yang, J.; Li, H.-M.; Hu, S.-J.; Xing, J.-L. Dorsal root ganglion compression as an animal model of sciatica and low back pain. Neurosci. Bull. 2012, 28, 618–630. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Y.H.; Ge, H.Y.; Arendt-Nielsen, L.; Wang, R.; Yue, S.W. A transient receptor potential vanilloid 4 contributes to mechanical allodynia following chronic compression of dorsal root ganglion in rats. Neurosci. Lett. 2008, 432, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Todaka, H.; Taniguchi, J.; Satoh, J.; Mizuno, A.; Suzuki, M. Warm temperature-sensitive transient receptor potential vanilloid 4 (TRPV4) plays an essential role in thermal hyperalgesia. J. Biol. Chem. 2004, 279, 35133–35138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.L.; Wang, Y.H.; Ning, L.P.; Zhang, Y.; Ge, H.Y.; Jiang, H.; Wang, R.; Yue, S.W. Involvement of TRPV4-NO-cGMP-PKG pathways in the development of thermal hyperalgesia following chronic compression of the dorsal root ganglion in rats. Behav. Brain Res. 2010, 208, 194–201. [Google Scholar] [CrossRef]
- Wang, C.; Ning, L.-P.; Wang, Y.-H.; Zhang, Y.; Ding, X.-L.; Ge, H.-Y.; Arendt-Nielsen, L.; Yue, S.-W. Nuclear factor-kappa B mediates TRPV4–NO pathway involved in thermal hyperalgesia following chronic compression of the dorsal root ganglion in rats. Behav. Brain Res. 2011, 221, 19–24. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.W.; Zhang, Y.; Yin, C.P.; Yue, S.W. Ca(2+) influx mediates the TRPV4-NO pathway in neuropathic hyperalgesia following chronic compression of the dorsal root ganglion. Neurosci. Lett. 2015, 588, 159–165. [Google Scholar] [CrossRef]
- Qu, Y.-J.; Zhang, X.; Fan, Z.-Z.; Huai, J.; Teng, Y.-B.; Zhang, Y.; Yue, S.-W. Effect of TRPV4-p38 MAPK Pathway on Neuropathic Pain in Rats with Chronic Compression of the Dorsal Root Ganglion. BioMed Res. Int. 2016, 2016, 6978923. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Gao, W.S.; Qi, L.; Jia, L.; Qu, Y.J.; Yue, S.W.; Zhang, Y. Effect of cGMP-activated aquaporin 1 on TRPV4 in rats with allodynia induced by chronic compression of the dorsal root ganglion. Neurosci. Lett. 2020, 716, 134630. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Lim, C.S.; Choi, H.; Joshi, H.P.; Kim, K.-T.; Kim, Y.H.; Park, C.-K.; Kim, H.M.; Han, I.-B. Elevated TRPV4 Levels Contribute to Endothelial Damage and Scarring in Experimental Spinal Cord Injury. J. Neurosci. 2020, 40, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, S.; Shinoda, M.; Hayashi, Y.; Saito, H.; Asano, S.; Kubo, A.; Shibuta, I.; Furukawa, A.; Toyofuku, A.; Iwata, K. Increase in IGF-1 Expression in the Injured Infraorbital Nerve and Possible Implications for Orofacial Neuropathic Pain. Int. J. Mol. Sci. 2019, 20, 6360. [Google Scholar] [CrossRef] [Green Version]
- Ando, M.; Hayashi, Y.; Hitomi, S.; Shibuta, I.; Furukawa, A.; Oto, T.; Inada, T.; Matsui, T.; Fukaya, C.; Noma, N.; et al. Oxytocin-Dependent Regulation of TRPs Expression in Trigeminal Ganglion Neurons Attenuates Orofacial Neuropathic Pain Following Infraorbital Nerve Injury in Rats. Int. J. Mol. Sci. 2020, 21, 9173. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, Q.; Lian, Y.; Chen, Y. Botulinum toxin type A reduces the expression of transient receptor potential melastatin 3 and transient receptor potential vanilloid type 4 in the trigeminal subnucleus caudalis of a rat model of trigeminal neuralgia. Neuroreport 2019, 30, 735–740. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, Y.; Fan, Z.Z.; Ge, H.Y.; Arendt-Nielsen, L.; Jiang, H.; Yao, W.; Yue, S.W. Effects of colchicine-induced microtubule depolymerization on TRPV4 in rats with chronic compression of the dorsal root ganglion. Neurosci. Lett. 2013, 534, 344–350. [Google Scholar] [CrossRef]
- Ning, L.; Wang, C.; Fan, X.; Ding, X.; Wang, Y.; Zhang, Y.; Wang, J.; Yue, S. Role of colchicine-induced microtubule depolymerization in hyperalgesia via TRPV4 in rats with chronic compression of the dorsal root ganglion. Neurol. Res. 2014, 36, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Prim. 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renton, T.; Yilmaz, Z.; Gaballah, K. Evaluation of trigeminal nerve injuries in relation to third molar surgery in a prospective patient cohort. Recommendations for prevention. Int. J. Oral Maxillofac. Surg. 2012, 41, 1509–1518. [Google Scholar] [CrossRef]
- Lin, C.-S.; Wu, S.-Y.; Huang, H.-Y.; Lai, Y.-L. Systematic Review and Meta-Analysis on Incidence of Altered Sensation of Mandibular Implant Surgery. PLoS ONE 2016, 11, e0154082. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Suttle, A.; Zhang, Q.; Wang, P.; Chen, Y. Transient Receptor Potential (TRP) Ion Channels in Orofacial Pain. Mol. Neurobiol. 2021, 58, 2836–2850. [Google Scholar] [CrossRef] [PubMed]
- Denadai-Souza, A.; Martin, L.; De Paula, M.A.V.; De Avellar, M.C.W.; Muscará, M.N.; Vergnolle, N.; Cenac, N. Role of transient receptor potential vanilloid 4 in rat joint inflammation. Arthritis Rheum. 2012, 64, 1848–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Williams, S.H.; McNulty, A.L.; Hong, J.H.; Lee, S.H.; Rothfusz, N.E.; Parekh, P.K.; Moore, C.; Gereau, R.W.; Taylor, A.B.; et al. Temporomandibular joint pain: A critical role for Trpv4 in the trigeminal ganglion. Pain 2013, 154, 1295–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boll, S.; Almeida de Minas, A.C.; Raftogianni, A.; Herpertz, S.C.; Grinevich, V. Oxytocin and Pain Perception: From Animal Models to Human Research. Neuroscience 2018, 387, 149–161. [Google Scholar] [CrossRef]
- Matak, I.; Bölcskei, K.; Bach-Rojecky, L.; Helyes, Z. Mechanisms of Botulinum Toxin Type A Action on Pain. Toxins 2019, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.H.; Greenwald, M.K.; Schwartz, A.G. Chemotherapy-Induced Peripheral Neuropathy: Mechanisms and Therapeutic Avenues. Neurotherapeutics 2021, 18, 2384–2396. [Google Scholar] [CrossRef]
- Islam, B.; Lustberg, M.; Staff, N.P.; Kolb, N.; Alberti, P.; Argyriou, A.A. Vinca alkaloids, thalidomide and eribulin-induced peripheral neurotoxicity: From pathogenesis to treatment. J. Peripher. Nerv. Syst. 2019, 24, S63–S73. [Google Scholar] [CrossRef]
- Alessandri-Haber, N. Transient Receptor Potential Vanilloid 4 Is Essential in Chemotherapy-Induced Neuropathic Pain in the Rat. J. Neurosci. 2004, 24, 4444–4452. [Google Scholar] [CrossRef] [Green Version]
- Alessandri-Haber, N.; Dina, O.A.; Joseph, E.K.; Reichling, D.B.; Levine, J.D. Interaction of Transient Receptor Potential Vanilloid 4, Integrin, and Src Tyrosine Kinase in Mechanical Hyperalgesia. J. Neurosci. 2008, 28, 1046–1057. [Google Scholar] [CrossRef] [Green Version]
- Alessandri-Haber, N.; Dina, O.A.; Chen, X.; Levine, J.D. TRPC1 and TRPC6 Channels Cooperate with TRPV4 to Mediate Mechanical Hyperalgesia and Nociceptor Sensitization. J. Neurosci. 2009, 29, 6217–6228. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, C.; Wang, Z.J. Proteinase-activated receptor 2 sensitizes transient receptor potential vanilloid 1, transient receptor potential vanilloid 4, and transient receptor potential ankyrin 1 in paclitaxel-induced neuropathic pain. Neuroscience 2011, 193, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Materazzi, S.; Fusi, C.; Benemei, S.; Pedretti, P.; Patacchini, R.; Nilius, B.; Prenen, J.; Creminon, C.; Geppetti, P.; Nassini, R. TRPA1 and TRPV4 mediate paclitaxel-induced peripheral neuropathy in mice via a glutathione-sensitive mechanism. Pflügers Arch.-Eur. J. Physiol. 2012, 463, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, S.; Wu, I.; Mata, M.; Fink, D.J. Activation of TLR-4 to produce tumour necrosis factor-alpha in neuropathic pain caused by paclitaxel. Eur. J. Pain 2015, 19, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Logu, F.; Trevisan, G.; Marone, I.M.; Coppi, E.; Padilha Dalenogare, D.; Titiz, M.; Marini, M.; Landini, L.; Souza Monteiro De Araujo, D.; Li Puma, S.; et al. Oxidative stress mediates thalidomide-induced pain by targeting peripheral TRPA1 and central TRPV4. BMC Biol. 2020, 18, 197. [Google Scholar] [CrossRef]
- Dalakas, M.C. Peripheral neuropathy and antiretroviral drugs. J. Peripher. Nerv. Syst. 2001, 6, 14–20. [Google Scholar] [CrossRef]
- Joseph, E.K.; Chen, X.; Khasar, S.G.; Levine, J.D. Novel mechanism of enhanced nociception in a model of AIDS therapy-induced painful peripheral neuropathy in the rat. Pain 2004, 107, 147–158. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Li, M.Y.; Li, Y.T.; Ning, J.Y.; Gou, X.C.; Shi, J.; Li, Y.Q. Expression and functional characterization of transient receptor potential vanilloid 4 in the dorsal root ganglion and spinal cord of diabetic rats with mechanical allodynia. Brain Res. Bull. 2020, 162, 30–39. [Google Scholar] [CrossRef]
- Cui, Y.-Y.; Xu, H.; Wu, H.-H.; Qi, J.; Shi, J.; Li, Y.-Q. Spatio-Temporal Expression and Functional Involvement of Transient Receptor Potential Vanilloid 1 in Diabetic Mechanical Allodynia in Rats. PLoS ONE 2014, 9, e102052. [Google Scholar] [CrossRef]
- Julian, T.; Glascow, N.; Syeed, R.; Zis, P. Alcohol-related peripheral neuropathy: A systematic review and meta-analysis. J. Neurol. 2019, 266, 2907–2919. [Google Scholar] [CrossRef]
- Behl, T.; Yadav, H.N.; Sharma, P.L. Alcoholic Neuropathy: Involvement of Multifaceted Signalling Mechanisms. Curr. Mol. Pharmacol. 2021, 14, 2–10. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Rodent Type | Model | Tissue | TRPV4 Expression Verification Method | Treatment | Dose, Time of Treatment, and Route of Administration | References |
|---|---|---|---|---|---|---|
| Rats (non-specific) | CCD | DRG | – | ODN | 40 µg i.t. once a day for 7 days | Zhang (2008) [46] |
| Wistar rats (male) | CCD | DRG | WB | ODN/antagonist (ruthenium red)/agonist (4α-PDD) | 40 µg i.t. once a day for 7 days/0.01–1 nmol i.t./1 nmol i.t. single administration | Ding (2010) [48] |
| Wistar rats (male) | CCD | DRG | – | Agonist (4α-PDD) | 1 nM i.t. single administration | Wang (2011) [49] |
| Wistar rats (male) | CCD | DRG | WB, RT-PCR, and IHC | – | – | Wei (2013) [52] |
| Wistar rats (male) | CCD | – | – | Agonist (4α-PDD) | 5 and 50 nmol/kg i.t. single administration | Ning (2014) [53] |
| Wistar rats (male) | CCD | DRG | WB, RT-PCR, and IHC | TRPV4 targeted siRNA (LV–TRPV4) | 10 µL/day for 3 days (continuously infused by i.t. catheter) | Wang (2015) [50] |
| Wistar rats (male) | CCD | DRG | WB | Antagonist (ruthenium red)/agonist (4α-PDD) | 1–100 nmol/l i.t. single administration | Qu (2016) [51] |
| Wistar rats (male) | CCD | DRG, and spinal cord | WB and RT-PCR | Agonist (GSK1016790A) | 0.5 µM i.t. single administration | Wei (2020) [52] |
| SD rats, C57BL/6 KO, and WT mice (female) | SCI | Spinal cord | WB, RT-PCR, and IHC | Antagonist (RN1734)/agonist (GSK1016790A) | 5 mg/kg, i.p./50 pmol i.t. single administration | Kumar (2020) [53] |
| SD rats (male) | IONI | TG | IHC | Antagonist (HC-067047) | 100 µg/kg s.c. single administration to the whisker pad | Sugawara (2019) [54] |
| SD rats (male) | IONI | TG | IHC | Antagonists (HC-067047 and RN1734) | HC067047 (30 mg/mL); RN1734 (0.4 g/mL) s.c. single administration to the whisker pad | Ando (2020) [55] |
| SD rats (male) | IONI | Vc | WB and IHC | – | – | Zhang (2019) [56] |
| Rodent Type | Model | Tissue | TRPV4 Expression Verification Method | Treatment | Dose, Time of Treatment and Route of Administration | References |
|---|---|---|---|---|---|---|
| SD rats (male) | Paclitaxel 1 mg/mL i.p. once a day for 10 days. | DRG | WB | ODN | 40 µg i.t. once a day for 3 days | Alessandri-Haber (2004) [69] |
| SD rats (male), KO and WT C57BL/6 mice (male) | Vincristine 200 µg/kg i.v. for 5 days (rats) or i.p. single injection (mice). Paclitaxel 1 mg/mL i.p. once a day for 10 days (rats) or 6 mg/kg single i.p. injection (mice). | – | – | ODN | 40 µg i.t. once a day for 3 days | Alessandri-Haber (2008) [70] |
| SD rats (male) | ddC 50 mg/kg i.v. single dose. | – | – | ODN | 40 µg i.t. once a day for 3 days | Alessandri-Haber (2008) [70] |
| SD rats (male) | Paclitaxel 1 mg/kg i.p. once a day for 10 days. | – | – | – | – | Alessandri-Haber (2009) [71] |
| ICR mice (male) | Paclitaxel 1 mg/kg i.p. every 2 days for 6 days. | – | – | Antagonist (RN1734) | 30 mg/kg i.p. single administration | Chen (2011) [72] |
| C57BL/6 WT and KO mice (male) | Paclitaxel 6 mg/kg i.p. single administration. | – | – | Antagonist (HC-067047) | 10 mg/kg i.p. single administration | Materazzi (2012) [73] |
| SD rats (male) | Paclitaxel 16 mg/kg i.p. once a week for 5 weeks. | DRG | WB, RT-qPCR, and IHC | – | – | Wu (2015) [74] |
| Swiss mice (male) | Paclitaxel 2 mg/kg i.p. for 5 days. | – | – | Antagonist (HC-067047) | 10 mg/kg, i.p. | Costa (2018) [23] |
| SD rats (male), C57BL/6J, WT and KO mice (male) | Thalidomide 1, 10, 50, and 100 mg/kg, pomalidomide 1 mg/kg, and lenalidomide 5 mg/kg, i.p. single injection. | spinal cord | – | Antagonist (HC-067047) | 10 mg/kg i.p. and 100 µg i.t. and i.pl. | De Logu (2020) [75] |
| Rodent Type | Model | Tissue | TRPV4 Expression Verification Method | Treatment | Dose, Time of Treatment, and Route of Administration | References |
|---|---|---|---|---|---|---|
| SD rats, KO and WT C57BL/6 mice (male). | Streptozotocin 50 mg/mL i.v. (rat) or 75 mg/kg i.p. (mice). | – | – | ODN | 40 µg i.t. once a day for 3 days. | Alessandri-Haber (2008) [70] |
| SD rats, KO and WT C57BL/6 mice (male). | Ethanol 6.5% i.g. for 4 days. | Saphenous nerve | WB | ODN | 40 µg i.t. once a day for 3 days. | Alessandri-Haber (2008) [70] |
| Swiss mice (male). | Streptozotocin 120 mg/kg i.p. for 2 consecutive days. | Sciatic nerve, DRG, and hind paw plantar skin | IHC | Antagonist (HC-067047) | 10 mg/kg, s.c. once a day for 6 consecutive days or 1 mg/kg once a day for 4 weeks. | Dias (2019) [34] |
| SD rats (male). | Streptozotocin 60 mg/kg i.p. single dose. | DRG, and spinal dorsal horn | WB, RT-PCR, and IHC | Antagonist (HC-067047), and agonist (GSK1016790A) | 400 ng/kg i.t. repeated injection (once a day for 7 consecutive days), or 200 ng/kg i.t. single injection. | Cui (2020) [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, P.; Ruviaro, N.A.; Trevisan, G. TRPV4 Role in Neuropathic Pain Mechanisms in Rodents. Antioxidants 2023, 12, 24. https://doi.org/10.3390/antiox12010024
Rodrigues P, Ruviaro NA, Trevisan G. TRPV4 Role in Neuropathic Pain Mechanisms in Rodents. Antioxidants. 2023; 12(1):24. https://doi.org/10.3390/antiox12010024
Chicago/Turabian StyleRodrigues, Patrícia, Náthaly Andrighetto Ruviaro, and Gabriela Trevisan. 2023. "TRPV4 Role in Neuropathic Pain Mechanisms in Rodents" Antioxidants 12, no. 1: 24. https://doi.org/10.3390/antiox12010024