Abstract
Short-term dietary nitrate (NO3−) supplementation has the potential to enhance performance during submaximal endurance, and short-duration, maximal-intensity exercise. However, it has yet to be determined whether NO3− supplementation before and during submaximal endurance exercise can improve performance during a short-duration, maximal-intensity end-sprint. In a randomised, double-blind, crossover study, 9 recreationally active men ingested NO3−-rich (BR: 8 mmol NO3−/day) and NO3−-depleted (PL: 0.75 mmol NO3−/day) beetroot powder for 7 days. On day 7, participants completed 2 h of moderate-intensity cycling, which immediately transitioned into a 60 s maximal-intensity end-sprint, with supplements ingested 2 h before and 1 h into the moderate-intensity exercise bout. Plasma [NO3−] and [NO2−] were higher in BR compared to PL pre- and post-exercise (p < 0.05). Post-exercise plasma [NO3−] was higher than pre-exercise (562 ± 89 µM vs. 300 ± 73 µM; p < 0.05) and plasma [NO2−] was not significantly different pre- (280 ± 58 nM) and post-exercise (228 ± 63 nM) in the BR condition (p > 0.05). Mean power output during the final 30 s of the end-sprint was greater after BR (390 ± 38 W) compared to PL (365 ± 41 W; p < 0.05). There were no differences between BR and PL in any muscle oxygenation variables during moderate-intensity cycling (p > 0.05), but muscle [deoxyhaemoglobin] kinetics was faster during the end-sprint in BR (6.5 ± 1.4 s) compared to PL (7.3 ± 1.4 s; p < 0.05). These findings suggest that NO3− supplementation has the potential to improve end-sprint performance in endurance events when ingested prior to and during exercise.
1. Introduction
Dietary supplementation with inorganic nitrate (NO3−) has emerged as a strategy to bolster performance in a variety of exercise settings in recreationally active individuals [1]. The ergogenic effects of NO3− ingestion have been attributed to the stepwise reduction of NO3− to nitrite (NO2−) and, subsequently, NO2− to nitric oxide (NO) [2]. It is now recognised that NO3− is absorbed in the upper gastrointestinal tract after ingestion with ~25% of circulating NO3− actively taken up by the salivary glands and concentrated in saliva [3,4]. Anaerobic bacteria in the oral cavity then reduce salivary NO3− to NO2− [5]. Once swallowed, a portion of this NO2− is further reduced to NO and other reactive nitrogen intermediates (RNIs), such as S-nitrosothiols (RSNO), in the stomach, with the remainder entering the systemic circulation, increasing plasma [NO2−], as well as [RSNO] [6,7]. More recent evidence also suggests skeletal muscle [NO3−] and [NO2−] can be increased following dietary NO3− supplementation and that skeletal muscle is an important NO3− storage organ [2,8,9]. Subsequently, circulating systemic and local muscle [NO3−] and [NO2−] can be used as substrates for O2 independent NO synthesis [2,8], with the potential to modulate an array of physiological processes [2,10,11].
Initial studies reported improved performance during continuous, submaximal endurance exercise after dietary NO3− ingestion [1,12,13], with NO3− supplementation now appearing to confer greater ergogenic potential for shorter duration higher intensity (<15 min), compared to longer duration lower intensity (>1000 s), endurance exercise [1]. Consistent with this observation, an initial study reported that dietary NO3− supplementation did not improve 50 mile cycling time trial (TT) performance (>120 min completion time); however, a negative correlation was observed between the increase in plasma [NO2−] and the lowering of 50 mile TT completion time after NO3− ingestion [14]. Accordingly, these data suggest greater potential for an ergogenic effect during longer duration endurance exercise in individuals presenting with higher plasma [NO2−] after NO3− ingestion. More recently, Tan et al. [15] reported that NO3− ingestion prior to and after 1 h of a 2 h moderate-intensity cycling task prevented the decline in plasma [NO2−], and blunted the progressive increases in the oxygen cost of exercise and muscle glycogen depletion, compared with placebo ingestion during exercise and when NO3− was only consumed pre-exercise. However, and despite better maintenance of plasma [NO2−] and improved metabolic efficiency after NO3− supplementation prior to and during exercise, performance in a subsequent 100 kJ cycling TT was not improved [15]. A necessary limitation of this study [15] was an interruption between the end of the moderate-intensity exercise bout and transition into the TT performance test to obtain muscle biopsy samples to assess metabolic responses. Such an interruption does not typically reflect real life competitive scenarios and may have permitted partial recovery of perturbations to muscle phosphocreatine (PCr), inorganic phosphate (Pi) and adenosine diphosphate (ADP), which are known to recover exponentially following exercise cessation [16]. Since NO3− supplementation has been reported to attenuate perturbation to these intramuscular phosphorous substrates and metabolites concomitant with enhanced exercise tolerance [12], this may account for the lack of an ergogenic effect in this previous study by Tan et al. [15]. It is also possible that, since the increase in muscle [NO3−] after NO3− supplementation declines during exercise [9] and increases rapidly (within 30 min) following acute NO3− ingestion [17], an in-exercise NO3− top-up bolus can better maintain the local increases in muscle [NO3−] and [NO2−] achieved after pre-exercise NO3− ingestion. Therefore, further research is required to evaluate the ergogenic efficacy of an in-exercise NO3− top-up dose for improving prolonged endurance performance.
Analysis of pacing strategies indicates that adopting a relatively even pacing strategy for the majority of the race followed by an end-of-race sprint (end-sprint) or ‘kick’ is commonly applied in competitive endurance events [18]. Although NO3− supplementation has been reported to improve muscle contractile function in a non-fatigued state [19,20,21], there is evidence to suggest that NO3− supplementation may be more likely to improve neuromuscular function after completing fatigue-inducing exercise [22]. Similarly, while NO3− supplementation has been reported to improve time to peak power output, peak power output and/or mean power output during maximal sprints initiated from an unfatigued state in some studies [23,24,25,26], it has yet to be determined whether these findings are reproducible when the sprint is commenced after a prolonged period of moderate-intensity endurance exercise.
As prolonged, submaximal exercise unfolds, there is a progressive recruitment of fast-twitch (type II) muscle fibres to maintain the required force production, as some of the initially recruited slow-twitch (type I) muscle fibres become substrate depleted and fatigued [27]. It is also well documented that the proportional recruitment of type II muscle fibres is positively associated with exercise intensity [28,29,30]. Accordingly, greater type II muscle fibre recruitment would be expected during the latter stages of an endurance event, particularly when terminated with an end-sprint. It is established that PO2 and pH are lowered to a greater extent in contracting type II relative to type I muscle [31,32], and that hypoxia and acidosis facilitate both NO2− reduction to NO [33,34] and NO3− reduction to NO2− in skeletal muscle [35]. As such, the physiological and metabolic conditions evoked by a prolonged period of moderate-intensity endurance exercise terminated with a maximal end-sprint has the potential to elicit an ergogenic effect after NO3− supplementation, but this postulate has not been experimentally tested. Moreover, it has previously been reported that muscle [deoxyhaemoglobin + deoxymyoglobin] ([HHb]) kinetics was faster, suggestive of enhanced muscle fractional O2 extraction during high-intensity exercise initiated after 4 min of moderate-intensity cycling exercise, concomitant with improved time to exhaustion [36]. However, it has yet to be determined whether [HHb] kinetics is altered during an end-sprint completed after prolonged moderate-intensity cycling exercise.
The purpose of the present study was to investigate whether ingestion of NO3−-rich beetroot powder before and during 2 h of moderate-intensity cycling influenced plasma [NO3−] and [NO2−], and end-sprint performance compared to a placebo condition. In addition, near-infrared spectroscopy (NIRS) was used to assess local muscle (de)oxygenation variables to provide physiological insight into any performance enhancement conferred by NO3− supplementation. It was hypothesized that NO3− supplementation before and during 2 h moderate-intensity exercise would: (1) further increase plasma [NO3−] and preserve elevated [NO2−]; (2) speed [HHb] kinetics; and (3) improve end-sprint performance.
2. Materials and Methods
2.1. Participant Characteristics
Nine recreationally active male participants (mean ± SD: age: 21 ± 1 years, stature: 1.81 ± 0.06 m, body mass: 77.4 ± 12.5 kg, O2peak: 49.0 ± 5.1 mL·kg−1·min−1) completed the study protocol. None of the participants were tobacco smokers. All experimental procedures were approved by Loughborough University Research Ethics Approvals Human Participants Sub Committee. Participants were fully informed of the risks and discomforts associated with all trials before providing written, informed consent. Participants were instructed to arrive at each laboratory testing session in a rested and hydrated state, having avoided strenuous exercise in the 24 h prior to each visit. Each participant was given a list of NO3−-rich foods to avoid consuming 24 h before testing sessions and asked to avoid caffeine and alcohol ingestion 12 h and 24 h before each trial, respectively. Participants were instructed to record their diet 24 h prior to their first experimental test and were asked to replicate this prior to each subsequent test. For the duration of the study, participants were asked to abstain from using antibacterial mouthwash since it blunts NO3− reduction to NO2− in the oral cavity [37].
2.2. Experimental Design
Participants reported to the laboratory on four occasions. During the first visit, participants completed a ramp incremental cycling test for determination of gas exchange threshold (GET), peak aerobic power (PAP) and peak oxygen uptake (O2peak). During the second visit, participants performed a 60 s all-out sprint on the cycle ergometer and were familiarised with the moderate-intensity exercise work rate. Thereafter, participants were assigned to receive two separate 7 day supplementation periods with a beetroot powder (TruBeet, Bio-gen Extracts, Bangalore, India) that was either NO3−-rich (BR) or NO3−-depleted (PL) as part of a randomised, cross-over experimental design. During the two main experimental testing sessions, which were completed on day 7 of the supplementation periods, participants completed 2 h of moderate-intensity cycling exercise which immediately transitioned into a 60 s end-sprint performance test. Muscle oxygenation variables were assessed, heart rate (HR) and ratings of perceived exertion (RPE) were determined, and venous blood samples were taken. All exercise testing was performed at the same time of day (±2 h) for each participant.
2.3. Incremental Test
During the first visit, participants completed a ramp incremental test on an electronically braked cycle ergometer (Lode Excalibur Sport, Groningen, The Netherlands). Initially, participants completed 4 min of baseline cycling at 20 W, after which the work rate increased linearly by 30 W/min until task failure. Task failure was recorded once the pedal rate fell ≥10 rpm below the participant’s self-selected cadence (70–90 rpm) for 5 s. The saddle and handlebar height and configuration were recorded and reproduced in subsequent tests. Breath-by-breath pulmonary gas exchange data were collected continuously (Vyntus CPX metabolic cart, Vyaire Medical, Chicago IL, USA) and averaged over consecutive 10 s periods. Participants wore a face mask and breathed through a low dead space, low resistance, digital volume transducer assembly. The inspired and expired gas volume and gas concentration signals were continuously sampled via a capillary line connected to the mouthpiece. The gas analyser was calibrated prior to testing with gases of known concentration. The turbine volume transducer was calibrated automatically and manually using a 3 L syringe (Hans Rudolph, Kansas City, MO, USA). O2peak was taken as the highest 30 s mean value attained prior to volitional exhaustion. GET was determined from a cluster of measurements including (1) the first disproportionate increase in CO2 production (CO2) from visual inspection of individual plots of
CO2 vs. O2, (2) an increase in minute ventilation (E)/O2 with no increase in E/CO2, and (3) an increase in end-tidal O2 tension with no fall in end-tidal CO2 tension. Work rates that would elicit 90% of GET (moderate-intensity exercise) and 50%Δ (GET plus 50% of the difference between power output at GET and PAP) were subsequently calculated, with two-thirds of the ramp rate deducted from the work rate at GET and PAP to account for the O2 mean response time.
2.4. Familiarisation Test
Participants completed a 60 s all-out sprint against a fixed and constant resistance. The resistance on the pedals was set using the cadence-dependent linear mode on the cycle ergometer so that participants attained a power output calculated to be 50%Δ upon attaining their self-selected cadence from the initial incremental test (linear factor = power output/preferred cadence2). Participants were provided with a 5 s countdown prior to the sprint and were instructed to attain peak power as quickly as possible and to continue exercising maximally for the duration of the effort. Participants were also familiarised with the moderate-intensity exercise work rate. This involved 2 min cycling at 25 W followed by a step increase in work rate equivalent to 90% GET for 10 min.
2.5. Supplementation Procedure
After the initial ramp test and familiarisation visits, participants were assigned to receive supplementation with BR containing 6% NO3− (8 mmol NO3−) or PL containing 0.56% NO3− (0.75 mmol NO3−) in a randomised, counter-balanced order. The PL powder was prepared by manipulating the processing control parameters (extraction time and temperature) during extract preparation to ensure that NO3− was not being completely extracted and was degraded/removed during the process. Specifically, raw beetroot was extracted with water for 1 h at >90 °C with the residue obtained from the first extraction processed to obtain the low NO3− grade material. The NO3− content of BR and PL were determined by treating the samples with a salicylic acid-sulphuric acid mixture, and subsequently 2 N sodium hydroxide, before measuring absorbance using a UV-Vis spectrophotometer (UV-1800, Shimadzu, Japan) with values compared to potassium NO3− standards. On days 1–6 of the supplementation periods, participants consumed 8.4 g of TruBeet powder mixed with ≥250 mL of water. On day 7 of supplementation, when the participants underwent the experimental testing procedures, participants were instructed to ingest the TruBeet powder 2 h before reporting to the laboratory to coincide with peak plasma [NO2−] [38]. In addition, 1 h into the exercise trials, participants were also given a top-up 8.4 g dose of TruBeet powder (PL or BR) in 250 mL water. Supplementation periods were separated by a minimum of 10 days washout and were administered double-blind.
2.6. Experimental Tests
After arrival at the laboratory and attachment of the NIRS device, participants rested in a supine position for 10 min before a venous blood sample was collected. Participants then commenced the exercise protocol. This consisted of 4 min baseline cycling at 25 W followed by 2 h cycling at 90% GET. HR and RPE were recorded every 10 min during the exercise test. The exercise bouts then immediately transitioned into a 60 s all-out sprint. Following completion of the exercise, participants rested supine for 10 min before a venous blood sample was taken.
2.7. Measurements
2.7.1. Sprint Performance
Power output was recorded continuously at 5 Hz using the Lode Excalibur Sport software and data were subsequently exported, reduced to second-by-second values and analysed to derive time to peak power output, peak power output and mean power outputs between 0–60 s, 0–30 s and 30–60 s.
2.7.2. Muscle Oxygenation
Changes in the oxygenation status of the m. vastus lateralis of the right leg during the protocol were continuously assessed using NIRS (PortaMon, Artinis Medical Systems, Einsteinweg, The Netherlands). Initially, the area surrounding the muscle belly was shaved and cleaned. The NIRS probe was affixed over the midway point between the superior border of the iliac crest and fibular head using kinesiology tape. Elastic strapping was then used to secure the device in place and minimise the possibility of extraneous light influencing the signal. The exact location of the device was recorded to enable precise reproduction of the placement in subsequent tests. The NIRS device emitted near-infrared light from 3 diodes at 760 and 850 nm, at distances of 30, 35 and 40 mm from the receiver. Data were sampled at 10 Hz and recorded with Oxysoft software (Artinins, Netherlands). During offline analysis, data were exported and averaged to 1 s intervals for later analysis. Relative changes in tissue saturation index (TSI) and concentrations of oxyhaemoglobin + oxymyoglobin (O2Hb), deoxyhaemoglobin + deoxymyoglobin (HHb) and total haemoglobin + myoglobin were determined.
To provide information on muscle deoxygenation kinetics, the [HHb] response to exercise was fitted with a monoexponetial model. Specifically, the [HHb] kinetics during the end-sprint bouts were determined by fitting a monoexponential model from the first data point 1 SD above the baseline mean through to the point at which the response departed from a monoexponentially, as determined from the residual plot and described in the following equation:
where HHb (t) represents the relative HHb at a given time t; HHbbaseline represents the mean HHb in the baseline period; A, TD, and τ represent the amplitude, time delay, and time constant, respectively, describing the increase in HHb above baseline. The [HHb] TD and τ values were summed to provide information on the overall [HHb] response dynamics. It has been reported that NIRS kinetics is a reliable method to assess muscle oxidative capacity following moderate-intensity exercise [39]. The [HHb] signal was used to provide an estimate of fractional O2 extraction in the area under interrogation. TSI, calculated as:
provides a percentage oxygenation in the area under interrogation.
HHb (t) = HHbbaseline + A (1 − e − (t−TD/τ))
TSI (%) = [O2Hb]/([O2Hb] + [HHb]) × 100
2.7.3. Ratings of Perceived Exertion and Heart Rate
Both RPE, using the 6–20 scale, and HR (Polar monitor, Kempele, Finland) were recorded every 10 min during the exercise tests.
2.7.4. Plasma [NO3−] and [NO2−]
Venous blood samples were drawn from an antecubital vein into 6 mL lithium-heparin tubes (Sarstedt, Leicester, UK) at baseline and post exercise. Samples were centrifuged at 3500× g for 10 min at 4 °C, within 3 min of collection. Plasma was subsequently extracted and immediately frozen at −80 °C for later analysis of [NO3−] and [NO2−].
2.8. Data Analysis Procedures
[NO3−] and [NO3−] Determination
All glassware, utensils and surfaces were rinsed with deionised water to remove residual NO prior to [NO3−] and [NO2−] analysis. Plasma samples were deproteinised using zinc sulfate (ZnSO4)/sodium hydroxide (NaOH) precipitation prior to [NO3−] determination. Firstly, 250 μL of 0.36 M NaOH was added to 50 μL of sample followed by 5 min incubation at room temperature. Subsequently, samples were treated with 150 μL of aqueous ZnSO4 (10% w/v) and vortexed for 30 s before undergoing an additional 10 min incubation period at room temperature. Samples were then centrifuged at 17,000× g for 8 min and the supernatant was removed for subsequent analysis. The [NO3−] of the deproteinised plasma sample was determined by its reduction to NO in the presence of 0.8% (w/v) vanadium chloride in 1 M hydrochloric acid within an air-tight purging vessel. The spectral emission of electronically excited nitrogen dioxide, derived from the reaction of NO with ozone, was detected by a thermoelectrically cooled, red-sensitive photomultiplier tube house in a Sievers gas-phase chemiluminescence nitric oxide analyser (Sievers NOA 280i, Analytix Ltd., Durham, UK). The [NO3−] was determined by plotting signal (mV) against a calibration plot of sodium NO3− standards. Plasma samples were deproteinised using ice-cold ethanol precipitation prior to [NO2−] determination. Specifically, 500 μL of sample was treated with 1000 μL ice-cold ethanol followed by 30 min incubation on ice. Samples were then centrifuged at 17,000× g for 8 min and the supernatant was removed for subsequent analysis. The [NO2−] of deproteinised plasma was determined by its reduction to NO in the presence of glacial acetic acid and aqueous sodium iodide (4% w/v) and calibrated using sodium NO2− standards.
2.9. Statistical Analysis
Two way (time × supplement) repeated-measures ANOVAs were used to assess changes in RPE, HR, and plasma [NO3−] and [NO2−]. Significant effects were followed up with post hoc paired t-tests with Holm-Bonferroni correction. Paired t-tests were used to assess changes in sprint performance and NIRS variables during the end-sprint. All data are presented as mean ± SD unless otherwise stated. Effect size (ES) for the ANOVAs were calculated using partial eta squared with ES for paired t-tests calculated as Cohen’s dz (t/√n). Statistical analysis was performed using IBM SPSS Statistics version 27. Statistical significance was accepted at p < 0.05.
3. Results
The PL and BR supplements in this study were well tolerated with no adverse side effects reported. Participants were unable to distinguish between the PL and BR supplements.
3.1. Plasma [NO3−] and [NO2−]
There were main effects for supplement (ES = 0.97) and time (ES = 0.98), and a supplement × time interaction effect (ES = 0.96) for plasma [NO3−] (all p < 0.001). Plasma [NO3−] was higher pre- (ES = 3.23) and post-exercise (ES = 5.91) in BR compared to PL (all p < 0.001, Table 1). There was no difference between plasma [NO3−] pre-to-post exercise in the PL condition (p > 0.05, ES = 1.08), but post-exercise plasma [NO3−] was 87% higher than pre-exercise plasma [NO3−] in the BR condition after receiving the top-up BR dose (p < 0.001, ES = 5.46). There were main effects for supplement (p < 0.001, ES = 0.86) and time (p < 0.05, ES = 0.47), but no supplement × time interaction effect for plasma [NO2−] (p > 0.05, ES = 0.15). Plasma [NO2−] was higher pre- (ES = 3.04) and post-exercise (ES = 0.993) in BR compared to PL but was not different between time points within the BR (ES = 0.63) and PL (ES = 0.22) conditions (all p < 0.05, Table 1).

Table 1.
Plasma nitrate and nitrite concentrations at rest and following 2 h moderate-intensity exercise and a 60 s maximal-intensity end-sprint after ingestion of nitrate-rich and nitrate-depleted beetroot powder.
3.2. Sprint Performance
Time to peak power output (p = 0.49, ES = 0.24) and peak power output (p = 0.61, ES = 0.18) did not differ between PL and BR (Table 2). Mean power output between 0–60 s was not significantly different between PL and BR (p = 0.058, ES = 0.74); Table 2). However, mean power output was greater in the BR condition between 30–60 s (p < 0.01, ES = 1.82; Figure 1 and Figure 2).
Table 2.
Time to peak power output, peak power output and mean power output variables during a maximal-intensity 60 s end-sprint following 2 h moderate-intensity exercise after ingestion of nitrate-rich and nitrate-depleted beetroot powder.
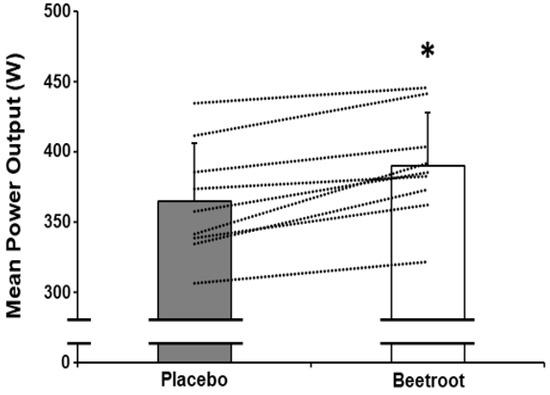
Figure 1.
Mean power output between 30–60 s of a maximal 60 s end-sprint following 2 h moderate-intensity exercise after ingestion of nitrate-depleted beetroot (placebo, grey bar) and nitrate-rich beetroot (beetroot, open bar) powder. Dashed lines represent individual responses. Data presented as mean ± SD. * indicates significantly higher than placebo (p < 0.001).
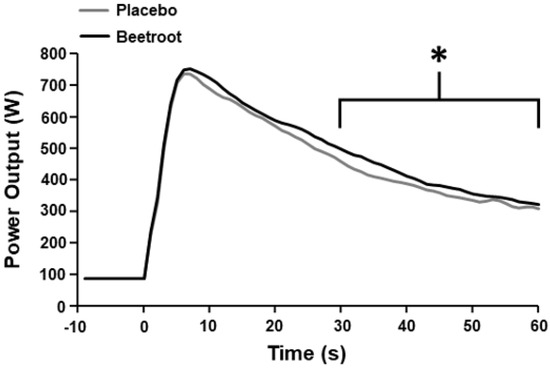
Figure 2.
Mean power output during the 60 s end-sprint following 2 h moderate-intensity exercise after ingestion of nitrate-depleted beetroot (placebo) and nitrate-rich beetroot (beetroot) powder. * Higher than placebo (p < 0.001). Error bars are omitted for clarity.
3.3. Muscle Oxygenation
After initial inspection, NIRS data from 7 of 9 participants was of sufficient quality for analysis and was used in the current paper. There were no between-condition differences in TSI during 2 h moderate-intensity cycling exercise or during the end-sprint (p > 0.05; Table 3). However, during the end-sprint muscle [HHb] τ + TD was lower in the BR compared to the PL condition (6.5 ± 1.4 s vs. 7.3 ± 1.4 s; p < 0.001, ES = 2.50; Figure 3).
Table 3.
Tissue saturation index during 2 h moderate-intensity exercise terminating in a maximal-intensity 60 s end-sprint after ingestion of nitrate-rich and nitrate-depleted beetroot powder.
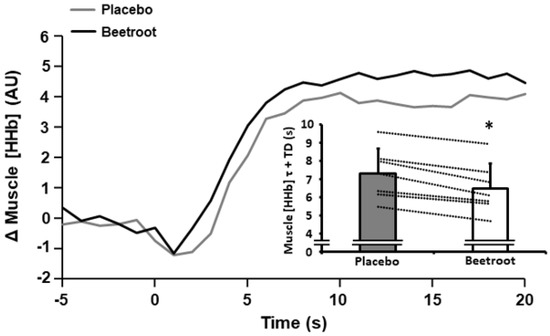
Figure 3.
Muscle deoxyhaemoglobin + deoxymyooglobin concentration ([HHb]) response profiles during the initial stages of a 60 s end-sprint following 2 h moderate-intensity exercise after ingestion of nitrate-depleted beetroot (placebo) and nitrate-rich beetroot (beetroot) powder in a representative individual. Data are expressed as the change (Δ) from baseline. Inset illustrates muscle [HHb] time constant (τ) + time delay (TD) after ingestion of placebo (grey bar) and beetroot (open bar) powder. Dashed lines represent individual responses. Data presented as mean ± SD. * indicates significantly lower than placebo (p < 0.001).
3.4. Ratings of Perceived Exertion and Heart Rate
There was a main effect of time (p < 0.001), but no effect of supplement (p > 0.05), or time × supplement interaction (p > 0.05) for RPE and HR (data not reported).
4. Discussion
Ingestion of NO3− prior to and 1 h into 2 h of moderate-intensity cycling culminating in a 60 s end-sprint, increased post-exercise compared to pre-exercise plasma [NO3−] and blunted the decline in plasma [NO2−] that typically occurs when NO3− is only consumed before exercise [15,40]. In addition, NO3− consumption before and during exercise speeded muscle [HHb] kinetics over the initial stages, and improved mean power output over the latter stages, of the 60 s end-sprint. These observations are consistent with our experimental hypotheses and suggest that short-term NO3− supplementation for 6 days, coupled with acute NO3− ingestion 2 h prior to and 1 h into a 2 h moderate-intensity exercise bout on day 7, has the potential to improve end-sprint performance. These findings may have implications for recreationally active participants aiming to enhance prolonged endurance exercise performance via dietary NO3− supplementation.
4.1. Effect of BR Supplementation on Plasma [NO3−] and [NO2−]
Plasma [NO3−] and [NO2−] were elevated pre-exercise following 7 days of NO3− supplementation with the final 8 mmol NO3− consumed 2 h prior to exercise, compared to PL. These findings are in line with previous research which has consistently reported elevations in both plasma [NO3−] and [NO2−] following similar dietary NO3− supplementation protocols [12,13,15]. The pharmacokinetic responses to acute dietary NO3− ingestion are well described, with peak plasma [NO3−] and [NO2−] expected to be, respectively obtained ~1 h and 2–3 h post ingestion of 8 mmol NO3− [38]. Plasma [NO3−] is relatively stable during exercise, at least up to 1 h, after NO3− ingestion [15]. In the present study, participants consumed an additional 8 mmol NO3− top-up bolus 1 h into a 2 h moderate-intensity cycle bout which was terminated with a 60 s maximal sprint with venous blood sampled 10 min thereafter. Compared to pre-exercise values after NO3− supplementation, the ingestion of an 8 mmol NO3− top-up bolus during exercise elicited a further increase in plasma [NO3−], with no changes between the pre-and post-exercise values after PL supplementation, consistent with the previous observations by Tan et al. [15]. In contrast, ingestion of an 8 mmol NO3− top-up bolus did not increase plasma [NO2−] post-exercise compared to pre-exercise values. Tan et al. [15] also observed no increase in post-exercise compared to pre-exercise plasma [NO2−] after ingestion of a NO3− top-up bolus 1 h into 2 h of moderate-intensity exercise followed by a 100 kJ cycling TT; conversely, these authors observed lower post-exercise compared to pre-exercise plasma [NO2−] in this condition as well as NO3− supplementation without the additional top-up bolus. While plasma [NO2−] is relatively stable during moderate-intensity exercise after NO3− ingestion [15,40], plasma [NO2−] has been reported to decline during high-intensity exercise in numerous studies [15,40,41,42]. The differences in the plasma [NO2−] dynamics between the present study and the previous study by Tan et al. [15] may be attributed to inter-study differences in the supplementation procedures and exercise testing protocol. Specifically, an 8 mmol NO3− top-up bolus was administered in the current study with 6 mmol administered in the study of Tan et al. [15], and the 2 h moderate-intensity cycle stage was followed by a 60 s maximal sprint in the current study compared to a longer 100 kJ cycling TT (completion time of ~400 s) in the study of Tan et al. [15]. Therefore, an important original contribution of the present study is that the ingestion of an 8 mmol NO3− top-up bolus can attenuate the decline in plasma [NO2−] that occurs during high-intensity exercise after only ingesting NO3− pre-exercise.
4.2. Effect of BR Supplementation on Maximal End-Sprint Performance
Compared to PL, 7 days of 8 mmol·day−1 NO3− supplementation with an 8 mmol top-up NO3− bolus consumed 1 h into a 2 h moderate-intensity cycle bout that was terminated with a 60 s maximal sprint, improved mean power output over the final 30 s of the sprint. To our knowledge, these are the first data to support enhanced performance after short-term NO3− supplementation with the consumption of an in-exercise NO3− top-up bolus. In a previous study by Tan et al. [15], 3 days of 12.4 mmol·day−1 NO3− supplementation with a 6.2 mmol top-up NO3− bolus consumed 1 h into a 2 h moderate-intensity cycle bout terminated with a 100 kJ cycling TT, did not enhance performance in the TT. These disparities may be linked to differences in supplementation procedures, performance test, and the interlude between the completion of the 2 h moderate-intensity exercise and commencement of the TT in the study of Tan et al. [15] to enable muscle biopsy sampling. Indeed, dietary NO3− intake was increased for 7 days versus 3 days, and the in-exercise NO3− top-up dose was 8 mmol versus 6 mmol in the present study versus the study by Tan et al. [15], respectively. Therefore, the greater total duration and dose of NO3− administered in the days preceding the performance test, and the greater in-exercise NO3− top-up dose may have contributed to the ergogenic effects reported in the current study and not in the study by Tan et al. [15]. It is also possible that the greater degree of skeletal muscle hypoxia and acidosis [12,40,43], and greater recruitment of type II muscle fibres [28,29,30], that would be expected during a maximal-intensity 60 s sprint versus a lower intensity longer duration TT test may be more conducive for NO2− reduction to NO [33,34], which may account for the greater ergogenic effect of NO3− supplementation in the present study compared to the previous study by Tan et al. [15]. Furthermore, the experimental protocol used by Tan et al. [15] incorporated a rest period between the moderate-intensity exercise bout and TT for muscle biopsy sampling, which may have permitted partial PCr, ADP and Pi recovery. Since NO3− supplementation may be ergogenic, at least in part through attenuating perturbations to muscle [PCr], [ADP] and [Pi], such effects may have been lost prior to commencing the TT in the study of Tan et al. [15] given the exponential recovery of these molecules [16], thereby limiting the ergogenic potential of NO3− supplementation in that study.
Although performance was improved over the final 30 s of the sprint in the present study, time to peak power output, peak power output and mean power output over the entire 60 s sprint did not differ between PL and BR. These results conflict with some, but not all, studies reporting enhanced sprint performance after NO3− supplementation when the sprint is completed in the absence of a 2 h moderate-intensity exercise pre-load [23,24,25,26]. While neuromuscular fatigue was not determined after 2 h moderate-intensity cycling exercise in the current study, neuromuscular fatigue development has been observed following extended moderate-intensity exercise [44]. Therefore, our observations are consistent with previous findings indicating that NO3− supplementation may be particularly effective at enhancing muscle function after exercise-induced neuromuscular fatigue [22]. The finding of enhanced performance over the second half, but not the first half, of the 60 s end-sprint is also consistent with this interpretation.
4.3. Effect of BR Supplementation on Muscle Oxygenation Variables
There was no effect of NO3− supplementation on NIRS-derived TSI during the moderate-intensity exercise bout in the current study. This observation conflicts with some, but not all, previous reports of improved lower limb muscle oxygenation during moderate-intensity exercise following dietary NO3− supplementation [13,40,41]. While muscle TSI was not altered during the 60 s end-sprint, HHb τ + TD was lower (faster) after NO3− supplementation. Faster muscle HHb kinetics in the transition from moderate-intensity to higher intensity, constant work rate, submaximal exercise after NO3− supplementation has previously been reported by Breese et al. [36]; however, our observations extent these previous findings to suggest that such effects can also occur after prolonged moderate-intensity exercise that transitions into a maximal-intensity end-sprint. Muscle [HHb] kinetics has been suggested to offer a non-invasive proxy for fractional muscle O2 extraction during exercise [45]. In the previous study by Breese et al. [36], faster muscle [HHb] kinetics (determined via frequency-domain NIRS) after NO3− supplementation was accompanied by faster pulmonary O2 kinetics, a non-invasive surrogate for muscle O2 kinetics [46]. While it is recognised that a limitation of the current study is the lack of assessment of pulmonary gas exchange variables, it is possible that faster muscle [HHb] kinetics after NO3− supplementation facilitated faster O2 kinetics. Since time to peak power output, peak power output and mean power over the first 30 s of the 60 s sprint did not differ between PL and BR, faster [HHb] kinetics were not a function of greater power output or cadence over the initial stages of the sprint [47]. Expedited muscle [HHb] kinetics may, therefore, have increased the proportional oxidative energy turnover, with a corresponding reduction in anaerobic energy turnover, to facilitate enhanced performance over the latter stages of the 60 s end-sprint. Indeed, such metabolic changes would be expected to lower the decline in muscle PCr utilisation and ADP and Pi accumulation, which contribute to neuromuscular fatigue development [48] and have been previously reported after NO3− supplementation [12].
4.4. Experimental Considerations and Implications
Improved performance during a 60 s end-sprint following 2 h moderate-intensity exercise after NO3− supplementation in the current study may have implications for improving race performance during longer duration endurance events. Indeed, it has been reported that a common pacing strategy during longer duration endurance events is a relatively even pace for the majority of a race with increased speed over the latter stages of the event. Specifically, over 10,000 m to half-marathon running events, most studies indicate that the optimal pacing strategy is a relatively even-pace with the race terminating in an all-out end-sprint [49]. Such pacing strategies are also employed to great effect by “sprinters” to win some Grand Tour cycling stages. As such, through improving end-sprint performance, NO3− supplementation has potential to improve performance outcomes in settings where a prolonged bout of relatively evenly paced endurance exercise is terminated with an end-sprint. However, it is acknowledged that alternative pacing strategies can be employed to deliver successful race outcomes, such as adopting a relatively constant, but higher mean race pace, or completing stochastic periods of increased mean race pace before returning to a relatively even race pace. Further research is required to assess the ergogenic potential of NO3− supplementation in such settings. Moreover, since better trained endurance athletes seem less receptive to an ergogenic effect of acute and short-term NO3− supplementation on endurance performance [50,51], further research is required to address whether NO3− supplementation prior to and during longer duration endurance exercise can elicit an ergogenic effect in this population.
Whilst the data presented in the current study suggest that NO3− supplementation prior to and during 2 h moderate-intensity exercise can improve end-sprint performance concomitant with faster muscle [HHb] kinetics, better maintenance of plasma [NO2−] and a further increase in plasma [NO3−], there are some limitations with the current study that are recognised. Firstly, the small sample size is acknowledged as a limitation of the current study. This may have precluded the detection of improved mean power over the whole 60 s end-sprint (p = 0.058). In addition, venous blood was only sampled pre- and post-exercise, and as such, it is unclear whether plasma [NO2−] was increase after 2 h moderate exercise following consumption of the NO3− top-up at 1 h of the moderate exercise protocol compared to pre-exercise values. It is also recognised that, since the current study did not evaluate the independent effects of short-term NO3− supplementation with the last dose ingested 2 h prior to exercise, or acute NO3− supplementation prior to and during exercise, it is unclear whether these NO3− supplementation strategies may have been comparatively ergogenic to the one adopted in the current study. However, the findings of the present study, combined with the previous study by Tan et al. [15] which did isolate the effect of consuming an in-exercise NO3− top-up, collectively support a NO3− top-up supplementation strategy to improve physiological and performance responses during longer duration endurance exercise.
Faster muscle [HHb] kinetics at the start of the end-sprint after NO3− supplementation in the current study should be interpreted with some caution as data were collected using a continuous-wave NIRS system. However, our findings are consistent with previous observations of faster muscle [HHb] kinetics in the transition from moderate-intensity to higher-intensity exercise after NO3− supplementation when using a superior, frequency-domain NIRS system [36]. In addition, while pre-exercise plasma [NO3−] was increased after NO3− supplementation, an in-exercise NO3− top-up after 1 h moderate-intensity exercise elicited a further increase in plasma [NO3−] following an additional 1 h of moderate-intensity exercise with a 60 s end-sprint compared to pre-exercise values. Since the increase in muscle [NO3−] after NO3− supplementation declines during exercise [9] and increases rapidly (within 30 min) following acute NO3− ingestion [17], an in-exercise NO3− top-up bolus after short-term NO3− supplementation may maintain local increases in muscle [NO3−] and [NO2−]. These changes may increase muscle NO synthesis during skeletal muscle contraction [2,8] and elicit important physiological responses, such has enhanced skeletal muscle calcium handling [52,53], culminating in improved exercise performance. Further research is therefore required to assess whether the NO3− supplementation strategy administered in the current study can impact skeletal muscle [NO3−] and [NO2−] dynamics during exercise, and the intramuscular mechanisms that underpin its ergogenic effect on longer duration endurance exercise.
5. Conclusions
The current study investigated the effects of short-term NO3− supplementation, with NO3− ingested 2 h prior to and 1 h into 2 h moderate-intensity cycling exercise terminating with a 60 s maximal-intensity end-sprint on day 7 of supplementation, on plasma [NO3−] and [NO2−], muscle [HHb] kinetics and end-sprint performance. Compared to pre-exercise values after NO3− supplementation, consumption of an in-exercise NO3− top-up elicited a further increase in plasma [NO3−] and attenuated the decline in plasma [NO2−] typically reported when NO3− is only consumed pre-exercise. Short-term NO3− supplementation combined with ingestion of a NO3− top-up during exercise speeded muscle [HHb] kinetics during the initial stages, and improved mean power output over the final 30 s, of the end-sprint. The findings of the current study reveal a NO3− supplementation strategy with the potential to enhance aspects of performance during prolonged endurance exercise in recreationally active individuals.
Author Contributions
Conceptualization, S.N.R., R.T., G.P.R. and S.J.B.; methodology, S.N.R., R.T., G.P.R. and S.J.B.; formal analysis, S.N.R., M.D.B. and S.J.B.; data curation, S.N.R. and S.J.B.; writing—original draft preparation, S.N.R. and S.J.B.; writing—review and editing, all authors.; visualization, all authors.; supervision, E.O., L.J.J. and S.J.B.; funding acquisition, S.J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a research grant to S.J.B. from Bio-Gen Extracts PVT Limited.
Institutional Review Board Statement
All experimental procedures were approved by Loughborough University Research Ethics Approvals Human Participants Sub Committee.
Informed Consent Statement
Participants were fully informed of the risks and discomforts associated with all trials and provided written, informed consent.
Data Availability Statement
Data are available from the corresponding author upon reasonable request.
Acknowledgments
This research was supported by the NIHR Leicester Biomedical Research Centre.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Senefeld, J.W.; Wiggins, C.C.; Regimbal, R.J.; Dominelli, P.B.; Baker, S.E.; Joyner, M.J. Ergogenic Effect of Nitrate Supplementation: A Systematic Review and Meta-analysis. Med. Sci. Sports Exerc. 2000, 52, 2250–2261. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Vanhatalo, A.; Seals, D.R.; Rossman, M.J.; Piknova, B.; Jonvik, K.L. Dietary Nitrate and Nitric Oxide Metabolism: Mouth, Circulation, Skeletal Muscle, and Exercise Performance. Med. Sci. Sports Exerc. 2001, 53, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Spiegelhalder, B.; Eisenbrand, G.; Preussmann, R. Influence of dietary nitrate on nitrite content of human saliva: Possible relevance to in vivo formation of N-nitroso compounds. Food Cosmet. Toxicol. 1976, 14, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Govoni, M. Inorganic nitrate is a possible source for systemic generation of nitric oxide. Free Radic. Biol. Med. 2004, 37, 395–400. [Google Scholar] [CrossRef]
- Bryan, N.S.; Burleigh, M.C.; Easton, C. The oral microbiome, nitric oxide and exercise performance. Nitric. Oxide 2022, 125–126, 23–30. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Cole, J.A.; Benjamin, N. Nitrate, bacteria and human health. Nat. Rev. Microbiol. 2004, 2, 593–602. [Google Scholar] [CrossRef]
- Abu-Alghayth, M.; Vanhatalo, A.; Wylie, L.J.; McDonagh, S.T.; Thompson, C.; Kadach, S.; Kerr, P.; Smallwood, M.J.; Jones, A.M.; Winyard, P.G. S-nitrosothiols, and other products of nitrate metabolism, are increased in multiple human blood compartments following ingestion of beetroot juice. Redox Biol. 2001, 43, 101974. [Google Scholar] [CrossRef]
- Piknova, B.; Schechter, A.N.; Park, J.W.; Vanhatalo, A.; Jones, A.M. Skeletal Muscle Nitrate as a Regulator of Systemic Nitric Oxide Homeostasis. Exerc. Sport Sci. Rev. 2022, 50, 2–13. [Google Scholar] [CrossRef]
- Wylie, L.J.; Park, J.W.; Vanhatalo, A.; Kadach, S.; Black, M.I.; Stoyanov, Z.; Schechter, A.N.; Jones, A.M.; Piknova, B. Human skeletal muscle nitrate store: Influence of dietary nitrate supplementation and exercise. J. Physiol. 2019, 597, 5565–5576. [Google Scholar] [CrossRef]
- Moon, Y.; Balke, J.E.; Madorma, D.; Siegel, M.P.; Knowels, G.; Brouckaert, P.; Buys, E.S.; Marcinek, D.J.; Percival, J.M. Nitric Oxide Regulates Skeletal Muscle Fatigue, Fiber Type, Microtubule Organization, and Mitochondrial ATP Synthesis Efficiency Through cGMP-Dependent Mechanisms. Antioxid Redox Signal. 2017, 26, 966–985. [Google Scholar] [CrossRef]
- Stamler, J.S.; Meissner, G. Physiology of nitric oxide in skeletal muscle. Physiol. Rev. 2001, 81, 209–237. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.J.; Fulford, J.; Vanhatalo, A.; Winyard, P.G.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation enhances muscle contractile efficiency during knee-extensor exercise in humans. J. Appl. Physiol. 2010, 109, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.J.; Winyard, P.; Vanhatalo, A.; Blackwell, J.R.; Dimenna, F.J.; Wilkerson, D.P.; Tarr, J.; Benjamin, N.; Jones, A.M. Dietary nitrate supplementation reduces the O2 cost of low-intensity exercise and enhances tolerance to high-intensity exercise in humans. J. Appl. Physiol. 2009, 107, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, D.P.; Hayward, G.M.; Bailey, S.J.; Vanhatalo, A.; Blackwell, J.R.; Jones, A.M. Influence of acute dietary nitrate supplementation on 50 mile time trial performance in well-trained cyclists. Eur. J. Appl. Physiol. 2012, 112, 4127–4134. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.; Wylie, L.J.; Thompson, C.; Blackwell, J.R.; Bailey, S.J.; Vanhatalo, A.; Jones, A.M. Beetroot juice ingestion during prolonged moderate-intensity exercise attenuates progressive rise in O2 uptake. J. Appl. Physiol. 2018, 124, 1254–1263. [Google Scholar] [CrossRef]
- Chidnok, W.; Fulford, J.; Bailey, S.J.; Dimenna, F.J.; Skiba, P.F.; Vanhatalo, A.; Jones, A.M. Muscle metabolic determinants of exercise tolerance following exhaustion: Relationship to the “critical power”. J. Appl. Physiol. 2013, 115, 243–250. [Google Scholar] [CrossRef]
- Kadach, S.; Piknova, B.; Black, M.I.; Park, J.W.; Wylie, L.J.; Stoyanov, Z.; Thomas, S.M.; McMahon, N.F.; Vanhatalo, A.; Schechter, A.N.; et al. Time course of human skeletal muscle nitrate and nitrite concentration changes following dietary nitrate ingestion. Nitric. Oxide 2022, 121, 1–10. [Google Scholar] [CrossRef]
- Thiel, C.; Foster, C.; Banzer, W.; De Koning, J. Pacing in Olympic track races: Competitive tactics versus best performance strategy. J. Sports Sci. 2010, 30, 1107–1115. [Google Scholar] [CrossRef]
- Coggan, A.R.; Leibowitz, J.L.; Kadkhodayan, A.; Thomas, D.P.; Ramamurthy, S.; Spearie, C.A.; Waller, S.; Farmer, M.; Peterson, L.R. Effect of acute dietary nitrate intake on maximal knee extensor speed and power in healthy men and women. Nitric. Oxide 2015, 48, 16–21. [Google Scholar] [CrossRef]
- Haider, G.; Folland, J.P. Nitrate supplementation enhances the contractile properties of human skeletal muscle. Med. Sci. Sports Exerc. 2014, 46, 2234–2243. [Google Scholar] [CrossRef]
- Whitfield, J.; Gamu, D.; Heigenhauser, G.J.F.; Van Loon, L.J.C.; Spriet, L.L.; Tupling, A.R.; Holloway, G.P. Beetroot Juice Increases Human Muscle Force without Changing Ca2+-Handling Proteins. Med. Sci. Sports Exerc. 2017, 49, 2016–2024. [Google Scholar] [CrossRef] [PubMed]
- Tillin, N.A.; Moudy, S.; Nourse, K.M.; Tyler, C.J. Nitrate Supplement Benefits Contractile Forces in Fatigued but Not Unfatigued Muscle. Med. Sci. Sports Exerc. 2018, 50, 2122–2131. [Google Scholar] [CrossRef] [PubMed]
- Jodra, P.; Domínguez, R.; Sánchez-Oliver, A.J.; Veiga-Herreros, P.; Bailey, S.J. Effect of Beetroot Juice Supplementation on Mood, Perceived Exertion, and Performance During a 30-Second Wingate Test. Int. J. Sports Physiol. Perform. 2020, 15, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Jonvik, K.L.; Nyakayiru, J.; Van Dijk, J.W.; Maase, K.; Ballak, S.B.; Senden, J.M.G.; Van Loon, L.J.C.; Verdijk, L.B. Repeated-sprint performance and plasma responses following beetroot juice supplementation do not differ between recreational, competitive and elite sprint athletes. Eur. J. Sport Sci. 2018, 18, 524–533. [Google Scholar] [CrossRef]
- Rimer, E.G.; Peterson, L.R.; Coggan, A.R.; Martin, J.C. Increase in Maximal Cycling Power with Acute Dietary Nitrate Supplementation. Int. J. Sports Physiol. Perform. 2016, 11, 715–720. [Google Scholar] [CrossRef]
- Wylie, L.J.; Bailey, S.J.; Kelly, J.; Blackwell, J.R.; Vanhatalo, A.; Jones, A.M. Influence of beetroot juice supplementation on intermittent exercise performance. Eur. J. Appl. Physiol. 2016, 116, 415–425. [Google Scholar] [CrossRef]
- Gollnick, P.D.; Piehl, K.; Saltin, B. Selective glycogen depletion pattern in human muscle fibres after exercise of varying intensity and at varying pedalling rates. J. Physiol. 1974, 241, 45–57. [Google Scholar] [CrossRef]
- Krustrup, P.; Söderlund, K.; Mohr, M.; Bangsbo, J. The slow component of oxygen uptake during intense, sub-maximal exercise in man is associated with additional fibre recruitment. Pflugers Arch. 2004, 447, 855–866. [Google Scholar] [CrossRef]
- Krustrup, P.; Söderlund, K.; Relu, M.U.; Ferguson, R.A.; Bangsbo, J. Heterogeneous recruitment of quadriceps muscle portions and fibre types during moderate intensity knee-extensor exercise: Effect of thigh occlusion. Scand. J. Med. Sci. Sports 2019, 19, 576–584. [Google Scholar] [CrossRef]
- Vøllestad, N.K.; Blom, P.C. Effect of varying exercise intensity on glycogen depletion in human muscle fibres. Acta Physiol. Scand. 1985, 125, 395–405. [Google Scholar] [CrossRef]
- Harkema, S.J.; Adams, G.R.; Meyer, R.A. Acidosis has no effect on the ATP cost of contraction in cat fast- and slow-twitch skeletal muscles. Am. J. Physiol. 1997, 272, 485–490. [Google Scholar] [CrossRef] [PubMed]
- McDonough, P.; Behnke, B.J.; Padilla, D.J.; Musch, T.I.; Poole, D.C. Control of microvascular oxygen pressures in rat muscles comprised of different fibre types. J. Physiol. 2005, 563, 903–913. [Google Scholar] [CrossRef] [PubMed]
- Castello, P.R.; David, P.S.; McClure, T.; Crook, Z.; Poyton, R.O. Mitochondrial cytochrome oxidase produces nitric oxide under hypoxic conditions: Implications for oxygen sensing and hypoxic signaling in eukaryotes. Cell Metab. 2006, 3, 277–287. [Google Scholar] [CrossRef]
- Modin, A.; Björne, H.; Herulf, M.; Alving, K.; Weitzberg, E.; Lundberg, J.O. Nitrite-derived nitric oxide: A possible mediator of ‘acidic-metabolic’ vasodilation. Acta Physiol. Scand. 2001, 171, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Piknova, B.; Park, J.W.; Lam, K.K.J.; Schechter, A.N. Nitrate as a source of nitrite and nitric oxide during exercise hyperemia in rat skeletal muscle. Nitric. Oxide 2016, 255–256, 54–61. [Google Scholar] [CrossRef]
- Breese, B.C.; McNarry, M.A.; Marwood, S.; Blackwell, J.R.; Bailey, S.J.; Jones, A.M. Beetroot juice supplementation speeds O2 uptake kinetics and improves exercise tolerance during severe-intensity exercise initiated from an elevated metabolic rate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Govoni, M.; Jansson, E.A.; Weitzberg, E.; Lundberg, J.O. The increase in plasma nitrite after a dietary nitrate load is markedly attenuated by an antibacterial mouthwash. Nitric. Oxide 2008, 19, 333–337. [Google Scholar] [CrossRef]
- Wylie, L.J.; Kelly, J.; Bailey, S.J.; Blackwell, J.R.; Skiba, P.F.; Winyard, P.G.; Jeukendrup, A.E.; Vanhatalo, A.; Jones, A.M. Beetroot juice and exercise: Pharmacodynamic and dose-response relationships. J. Appl. Physiol. 2013, 115, 325–336. [Google Scholar] [CrossRef]
- de Aguiar, R.A.; Turnes, T.; Borszcz, F.K.; Raimundo, J.A.G.; Caputo, F. Near-infrared spectroscopy-derived muscle O2 kinetics after moderate running exercise in healthy males: Reliability and associations with parameters of aerobic fitness. Exp. Physiol. 2022, 107, 476–488. [Google Scholar] [CrossRef]
- Cocksedge, S.P.; Breese, B.C.; Morgan, P.T.; Nogueira, L.; Thompson, C.; Wylie, L.J.; Jones, A.M.; Bailey, S.J. Influence of muscle oxygenation and nitrate-rich beetroot juice supplementation on O2 uptake kinetics and exercise tolerance. Nitric. Oxide 2020, 99, 25–33. [Google Scholar] [CrossRef]
- Kelly, J.; Vanhatalo, A.; Bailey, S.J.; Wylie, L.J.; Tucker, C.; List, S.; Winyard, P.G.; Jones, A.M. Dietary nitrate supplementation: Effects on plasma nitrite and pulmonary O2 uptake dynamics during exercise in hypoxia and normoxia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Wylie, L.J.; Mohr, M.; Krustrup, P.; Jackman, S.R.; Ermιdis, G.; Kelly, J.; Black, M.I.; Bailey, S.J.; Vanhatalo, A.; Jones, A.M. Dietary nitrate supplementation improves team sport-specific intense intermittent exercise performance. Eur. J. Appl. Physiol. 2013, 113, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.S.; Noyszewski, E.A.; Kendrick, K.F.; Leigh, J.S.; Wagner, P.D. Myoglobin O2 desaturation during exercise. Evidence of limited O2 transport. J. Clin. Investig. 1995, 96, 1916–1926. [Google Scholar] [CrossRef] [PubMed]
- Iannetta, D.; Zhang, J.; Murias, J.M.; Aboodarda, S.J. Neuromuscular and perceptual mechanisms of fatigue accompanying task failure in response to moderate-, heavy-, severe-, and extreme-intensity cycling. J. Appl. Physiol. 2022, 133, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Koga, S.; Kano, Y.; Barstow, T.J.; Ferreira, L.F.; Ohmae, E.; Sudo, M.; Poole, D.C. Kinetics of muscle deoxygenation and microvascular PO2 during contractions in rat: Comparison of optical spectroscopy and phosphorescence-quenching techniques. J. Appl. Physiol. 2012, 112, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Krustrup, P.; Jones, A.M.; Wilkerson, D.P.; Calbet, J.A.; Bangsbo, J. Muscular and pulmonary O2 uptake kinetics during moderate- and high-intensity sub-maximal knee-extensor exercise in humans. J. Physiol. 2009, 587, 1843–1856. [Google Scholar] [CrossRef] [PubMed]
- Brock, K.; Antonellis, P.; Black, M.I.; DiMenna, F.J.; Vanhatalo, A.; Jones, A.M.; Bailey, S.J. Improvement of Oxygen-Uptake Kinetics and Cycling Performance with Combined Prior Exercise and Fast Start. Int. J. Sports Physiol. Perform. 2018, 13, 305–312. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- Casado, A.; Hanley, B.; Jiménez-Reyes, P.; Renfree, A. Pacing profiles and tactical behaviors of elite runners. J. Sport Health Sci. 2021, 10, 537–549. [Google Scholar] [CrossRef]
- Cermak, N.M.; Res, P.; Stinkens, R.; Lundberg, J.O.; Gibala, M.J.; van Loon, L.J. No improvement in endurance performance after a single dose of beetroot juice. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 470–478. [Google Scholar] [CrossRef]
- Robinson, G.P.; Killer, S.C.; Stoyanov, Z.; Stephens, H.; Read, L.; James, L.J.; Bailey, S.J. Influence of Dietary Nitrate Supplementation on High-Intensity Intermittent Running Performance at Different Doses of Normobaric Hypoxia in Endurance-Trained Males. Int. J. Sport Nutr. Exerc. Metab. 2001, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.J.; Gandra, P.G.; Jones, A.M.; Hogan, M.C.; Nogueira, L. Incubation with sodium nitrite attenuates fatigue development in intact single mouse fibres at physiological PO2. J. Physiol. 2019, 597, 5429–5443. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Schiffer, T.A.; Ivarsson, N.; Cheng, A.J.; Bruton, J.D.; Lundberg, J.O.; Weitzberg, E.; Westerblad, H. Dietary nitrate increases tetanic [Ca2+]i and contractile force in mouse fast-twitch muscle. J. Physiol. 2012, 590, 3575–3583. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).