
Hydrogen: A Rising Star in Gas Medicine as a Mitochondria-Targeting Nutrient via Activating Keap1-Nrf2 Antioxidant System


1
Center for Mitochondrial Biology and Medicine, The Key Laboratory of Biomedical Information Engineering of Ministry of Education, School of Life Science and Technology, Xi’an Jiaotong University, Xi’an 710049, China
2
Cardiometabolic Innovation Center, Ministry of Education, Department of Cardiology, First Affiliated Hospital of Xi’an Jiaotong University, Xi’an 710061, China
3
School of Health and Life Sciences, University of Health and Rehabilitation Sciences, Qingdao 266071, China
*
Authors to whom correspondence should be addressed.
Antioxidants 2023, 12(12), 2062; https://doi.org/10.3390/antiox12122062
Submission received: 25 October 2023
/
Revised: 27 November 2023
/
Accepted: 28 November 2023
/
Published: 30 November 2023
(This article belongs to the Special Issue Recent Advances in Redox Biology Research in China)
Abstract
:The gas molecules O2, NO, H2S, CO, and CH4, have been increasingly used for medical purposes. Other than these gas molecules, H2 is the smallest diatomic molecule in nature and has become a rising star in gas medicine in the past few decades. As a non-toxic and easily accessible gas, H2 has shown preventive and therapeutic effects on various diseases of the respiratory, cardiovascular, central nervous system, and other systems, but the mechanisms are still unclear and even controversial, especially the mechanism of H2 as a selective radical scavenger. Mitochondria are the main organelles regulating energy metabolism in living organisms as well as the main organelle of reactive oxygen species’ generation and targeting. We propose that the protective role of H2 may be mainly dependent on its unique ability to penetrate every aspect of cells to regulate mitochondrial homeostasis by activating the Keap1-Nrf2 phase II antioxidant system rather than its direct free radical scavenging activity. In this review, we summarize the protective effects and focus on the mechanism of H2 as a mitochondria-targeting nutrient by activating the Keap1-Nrf2 system in different disease models. In addition, we wish to provide a more rational theoretical support for the medical applications of hydrogen.
1. Introduction
Gas molecules are increasingly being used for medical purposes and their application has developed into a separate field of medicine. The gases most widely used in medicine include oxygen (O2), nitric oxide (NO), methane (CH4), carbon monoxide (CO), hydrogen sulfide (H2S), and hydrogen (H2). As shown in Figure 1, the number of articles related to medical gas molecules has grown substantially from 1998 to 2022, especially articles related to O2 and H2.
O2 and NO are the two medical gas molecules that most attract researchers’ attention, with tens of thousands of studies focusing on these two gases published since as early as the 1990s. O2 is the most crucial gas for all living organisms on earth and accounts for around 1/5 of the volume of air. As an important gas to maintain human respiration, O2 is mainly used to provide supplemental respiration for the sick, astronauts traveling in space, mountaineers, etc. In addition, it has the function of destroying bacteria. Due to the importance of O2, the 2019 Nobel Prize in Physiology or Medicine was awarded to William G. Kaelin Jr., Sir Peter J. Ratcliffe, and Gregg L. Semenza, who discovered how cells sense and adapt to the availability of O2 [1]. NO, commonly applied in the biomedical field in the form of NO· as a free radical gas, was found to work as a mediator of cell-to-cell communication in vasodilatation, inflammation, and neurotransmission at the end of the last century. Robert F. Furchgott, Louis J. Ignarro, and Ferid Murad et al. demonstrated that NO· is an important signaling molecule in the cardiovascular system, and this discovery won the 1998 Nobel Prize in Physics or Medicine [2].
CH4 is the simplest of the organic compounds. For decades, CH4 was thought to have almost no physiological role, while in the last few years, scientists have realized that CH4 can play important biological roles such as anti-inflammatory, antioxidant, and antiapoptotic roles. As a result, CH4 has been used as a gastric decontaminant in emergency clinical settings of poisoning or drug overdose and serves as a passive indicator of colonic function [3].
CO and H2S have long been known as hazardous factors. Long-term exposure to environments which are rich in CO may be fatal. However, a growing amount of research suggests that CO is an important gaseous mediator along with NO· and H2S. Endogenously produced or inhaled CO has important physiological functions in regulating vascular function, inflammation, apoptosis, cell proliferation, and signaling pathways. Studies have shown that inhaled CO suppresses chronic inflammation in patients with stable chronic obstructive pulmonary disease (COPD) [4]. Initially, H2S was regarded as a harmful gas since exposure to H2S irritates the eyes and respiratory system. However, scientists have now shown that H2S is an essential physiological factor, as it is produced by bacteria in the human oral cavity and gastrointestinal tract. As the least appreciated of the three gaseous mediators (gas transport mediators), it is now considered to be an important gas transport mediator after NO· and CO. H2S has been shown to modulate many physiological processes such as vasodilation, anti-inflammation, resistance to oxidative stress, protection against ischemia-reperfusion injury, etc. [5].
As the smallest of all molecules, the functions of H2 have also caught the eye of scientists in the field of biomedicine. As early as the beginning of the last century, H2 was first tested as a diving gas, proving that it is the best breathing medium for medium and deep diving and is safe for the organism, with no toxic side effects found. To date, H2 biomedicine has investigated the effects and mechanisms by which H2 molecules, including H+ ions (pH) and H− ions (a powerful reducing agent existing as MgH2, CaH2, AlLiH4, etc.) and heavy H2 (deuterium and tritium), act in various diseases [6].
2. History of H2 Medicine
H2 is known to be a colorless, odorless, and tasteless gas that is chemically stable [7]. In general, around 35 mL to 321 mL of H2 is produced and released through bacterial fermentation by the human digestive system per day [8]. Several ways are used to ingest or consume H2, such as drinking or injecting H2 water (HW), inhalation of H2, bathing in HW, dropping H2 saline into the eyes, etc. H2 plays an anti-inflammatory and antiapoptotic role through its selective antioxidant properties and has become a unique cytoprotective agent [9].
H2 used to be considered an inert gas not involved in any life activity. It was not until 1975 that Dole et al. found significant regression of mouse skin tumors in squamous cell carcinoma mice exposed for a fortnight to a mixture of 97.5% H2 and 2.5% O2 at a total pressure of eight atmospheres, first confirming the medical usefulness of H2 [10]. Unfortunately, this study has not attracted academic attention due to the technical difficulties of applying hyperbaric H2 therapy in a clinical application.
In 1996, Chinese scientist Yuanwei Du noticed the significance of H2 for life [11]. Dr. Du believes that excessive accumulation of peroxides produced in the metabolic process is the root cause of various diseases and aging; the organism must have a certain mechanism to fight against these peroxides. H2 is a reducing agent which can eliminate peroxides naturally without side effects, making creatures achieve a balance in the sense of redox balance. In Du’s experiment, tritium gas was produced by electrolysis of tritium water. The tritium gas was then fed, instead of H2, into the mouse’s living environment. He found that tritium was present in all tissues and organs of mice, which means that tritium gas is involved in the life activities of living organisms by transforming into tritium ions prevalent in living organisms, indirectly proving that the H2 in air is both a constituent substance and an energetic substance of life. This experiment also proves the basic mechanism of H2 metabolism. A number of H2 medicine-related papers published by Yuanwei Du at the end of the 20th century further confirmed that H2 produced by water electrolysis has a pronounced effect on the vital activities of plants (lilac branches), animals (mice), as well as humans [12]. Du’s work creatively combines the physiological effects of H2 with the free radical aging theory, explains the antioxidant activity of H2 molecules, and confirms that H2 may have an immeasurable effect on a wide range of diseases.
In 2007, Ohsawa et al. from the Nippon Medical School published an important article on H2 medicine in the journal Nature Medicine [13]. This study used a low concentration of H2 (1–4%) for inhalation over a short period (35 min) by mice and found positive effects in the treatment of cerebral ischemia-reperfusion injury, showing that short-term inhalation of a low concentration of H2 for the treatment of the disease is feasible. They proposed a mechanism whereby H2 could act as a therapeutic antioxidant, selectively reducing cytotoxic oxygen radicals (•OH and ONOO−), leading to the inhibition of cerebral ischemia-reperfusion injury. Because this study was published in the prestigious Nature Medicine journal, it provided a broad prospect for both basic and clinical research on H2 and brought H2 medicine to the attention of a wide range of academic cycles. Since then, more and more scholars have joined the research on H2 medicine to explore its effects on various diseases such as inflammation, drug toxicity, and obesity. More than a thousand peer-reviewed research papers have been published to date.
In the beginning, scientists focused mainly on acute and chronic organ injuries related to oxidative stress, such as those found in animal experiments investigating drug toxic injury or ischemia-reperfusion injury in vital organs such as the heart and liver. During this period, researchers mostly used diverse injury models to validate the therapeutical effects of H2 inhalation. Between 2009 and 2012, more research began to appear on drinking H2-enriched water (HRW) [14], injecting H2-enriched saline (HRS) [15], as well as studies on boosting H2 replenishment through gut bacteria [16]. Meanwhile, a number of clinical studies have used HRW in the treatment of diseases including metabolic syndrome, Parkinson’s disease, hemodialysis, sports injuries, and rheumatoid arthritis [9]. For the past few years, on the foundation of previous studies, H2 medicine research has studied molecular mechanisms, especially focusing on the molecular pathways of inflammation and oxidative stress mediated by H2. However, regarding the molecular mechanism of H2, most scholars have followed the view of Ohsawa et al. in their paper in Nature Medicine: that H2 is a selective hydroxyl radical (•OH) scavenger. As a result, most scholars have focused on the antioxidant mechanisms of H2 based on this classification [17,18]. Nevertheless, some scholars have proposed that H2 plays a signaling role that may be involved in metabolic processes and may even provide energy for bacteria, which broadens thoughts on the development of H2 research [19,20,21].
A number of Chinese researchers have devoted themselves to developing H2 medicine. These researchers has received more than 80 grants from the National Natural Science Foundation of China and have published hundreds of basic and clinical academic papers. Prof. Xuejun Sun of the Second Military Medical University is one of the leading figures in H2 medicine in China. Prof. Sun’s group engaged in the diving hyperbaric medicine research for a long time. The most important research object of diving hyperbaric medicine is the types of gases that can be breathed by human beings, with H2 being one of the key types of gases in the field of diving hyperbaric medicine. Sun’s group focuses on the biological effects of H2 and its application in medicine for the first time, revealing the value of H2 in medicine in China. Moreover, Prof. Sun participated in organizing several international symposiums on H2 medicine, inviting experts from all over the world to discuss the future of H2 medicine. His team collaborates with medical organizations around the world to carry out research on the application of H2 medicine and to expand the scope of H2 applications in the medical field.
Prof. Shucun Qin of Shandong First Medical University is another key promoter of H2 molecular medicine in China. Prof. Qin established the first H2 Biomedical Research Institute at the university in 2015, training a number of key researchers in H2 medicine. He established the standardized laboratory for H2 molecular biology that has published multiple placebo-controlled population trials, providing important clinical evidence for the translation of H2 into medicine. Qin’s recent review summarizes 51 clinical trials involving 1213 subjects in four areas of H2 biomedicine: basic research, exercise, dermatology, and healthcare [22]. The results showed that H2 can reduce oxidative stress damage caused by strenuous exercise, reduce lactic acid build-up after exercise, prevent exercise acidosis, and reduce exercise fatigue. In addition, H2 intervention can play a positive role in skin beauty and improve cardiovascular health.
Prof. Xuemei Ma’s team at the Beijing Institute of Technology was also an early group of H2 medicine researchers in China. Prof. Ma is committed to elucidating the biological basis of H2 medicine at the molecular, cellular, and holistic levels, conducting in-depth basic research and clinical translational research, especially on the mechanism of H2 molecules on tumor prevention. Her team has verified that H2 can inhibit the proliferation of gliomas (Gliomas) by inducing glial stem cell differentiation in in vitro and in vivo experiments [23].
In addition to these key researchers, there are hundreds of scientists doing work on H2 medicine, including the Chinese academicians Prof. Nanshan Zhong, Zhaofen Xia, Hongyang Wang, and young scientists like Prof. Qianjun, who proposed the concept of H2 nanomedicine to address the issues of H2 medicine by using functional micro/nanomaterials for augmented H2 therapy in cancer, and Wenbiao Shen, who is devoted to the application of H2 in agriculture. An academic association for H2 medicine with more than 400 members has been formed. As of today, current clinical studies on H2 are still continuously emerging, and the scale of the studies is gradually expanding. With its favorable biosafety and convenience, an H2 inhalation device has been included in the Chinese National Medical Products Administration’s new medical device development process (Registration Approval No. 20203080066). Moreover, in Japan, H2 has been approved as a food supplement [9].
3. H2: A Mitochondria-Targeting Molecule/Nutrient Rather Than a Selective •OH Scavenger
Sustained oxidative stress leads to the onset and progression of many common diseases. To date, little has been achieved in this regard, although a large number of studies have attempted to develop an effective antioxidant without side effects. Mitochondria, as a major source of oxidative stress, is considered a new therapeutic target for small molecule interventions [24]. H2 suppresses reactive oxygen species (ROS) accumulation, inhibits the cell death program, and maintains mitochondrial structure and function [25,26]. Preliminary clinical trials suggest that drinking H2 dissolved in water appears to improve the pathology of mitochondrial disease [27,28].
Mitochondria have a double membrane structure that forms the difference in potentials between the inner and outer membranes and controls the movement of diverse molecules and factors (e.g., ions) in and out of the organelle while affecting mitochondrial stability. Although the outer membrane is comparatively permeable to small molecules and large proteins (which are transported by diffusion or transposases), the inner mitochondrial membrane is highly impermeable to most molecules [29]. Special membrane transport proteins (e.g., TIM-TOM (preprotein translocase of the inner membrane of mitochondria-preprotein translocase of the outer membrane of mitochondria) complex, etc.) are needed for all ions and molecules to enter or leave the mitochondrial matrix. This means most antioxidants cannot enter the mitochondria to effectively scavenge •OH [30,31]. The difference with other antioxidants is that, as the smallest molecule in nature, H2 can easily spread and penetrate into the cell membrane to react with organelles such as mitochondria and the nucleus [32].
While the idea that H2 is a selective antioxidant has been popularized [7], it is still not known whether the effects of H2 arise from the direct reaction with •OH or from the inhibition of •OH production. Let us first provide some basic information on free radicals.
As we know, •OH is generated by the Haber−Weiss reaction:
O2•− + H2O2 → O2 + •OH + OH−
This reaction is thermodynamically feasible but kinetically too slow. So, •OH is mainly generated by the Fenton reaction:
Fe2+ + H2O2 → Fe3+ + •OH + OH−
The three main properties of •OH are below: 1. Short life: •OH has a very short half-life (10−9 s, or 1 ns, whereas the half-life of superoxide is 15 s) and requires no time to diffuse (no more than 50 molecular diameters from the site of formation), so the reaction is local with the antioxidant found where •OH is produced. 2. High reactivity: •OH is the ROS with the highest reduction potential; compared to other oxygen species, it reacts with extremely high rate constants (high reactivity) that approach diffusion-limited rate constants, with rate constants of 109–1010 M−1 s−1. So, •OH is the strongest (most powerful) oxidant of the oxyradicals. 3. Unselective and indiscriminate: •OH reacts unselectively and indiscriminately with almost every type of molecule found in living cells, including lipids, proteins, amino acids, DNA, RNA, and sugars. Therefore, the best antioxidant is not a •OH scavenger but, rather, an iron chelate to prevent the generation of •OH.
The reaction with many substances in the body occurs at a rate that exceeds that of H2, which means that H2 has difficulty competing with these molecules effectively in the body, especially when H2 is at a relatively lower concentration than other endogenous substances. Biokinetic analyses of the intracellular reactions of •OH/ONOO− show that intracellular molecules, such as nucleic acids and amino acids, react with •OH more readily at a significantly faster rate than H2 [33,34], which implies that H2 can hardly act as an •OH scavenger or barely directly react with •OH.
In 2005, we first proposed the new concept of “mitochondrial nutrient”. The so-called “Mitochondrial nutrients” refer to any compound that can protect mitochondria from damage, repair mitochondria injury, and promote mitochondrial function. Their mechanisms of action may include (1) protecting mitochondrial enzymes and/or stimulating enzyme activity by increasing the levels of substrate and cofactors; (2) inducing the activation of endogenous antioxidant systems, such as phase II enzymes, to enhance antioxidant defense; (3) preventing mitochondria from producing ROS and removing ROS in mitochondria, and (4) protecting and repairing mitochondrial damage, including energy promoters [35,36,37].
Researchers in our lab reported that in an LPS-induced lung injury mouse model, hyperoxic HRS effectively reduced mitochondrial swelling and cristae breaks and significantly reversed the reduction of mitochondrial complex I, IV, and V activities [38,39]. Not coincidentally, in a high-fat diet (HFD)-induced liver injury model, coral calcium hydride (CCH, a solid form of molecular H2 carrier made from coral calcium) treatment improved glucose and lipid metabolism, ameliorated hepatic mitochondria abnormalities, restored the protein expression and the activity of complex II, and activated phase II enzymes [38,39]. These studies imply that H2, as a highly promising mitochondrial nutrient, is able to target mitochondria.
Ohsawa et al. [13] used antimycin A (an inhibitor of mitochondrial respiratory complex III) to induce excess O2•− production. In this model, O2•− rapidly converted to H2O2, which was further converted to •OH. Their result showed that H2 treatment prevented the decrease in mitochondrial membrane potential caused by antimycin A treatment. They concluded that H2 protects mitochondria from •OH damage. The researchers hypothesized that H2 enters the mitochondria and acts on the mitochondrial respiratory chain, weakening the Fenton reaction by inhibiting transition metal activity and, ultimately, inhibiting •OH production but not scavenging •OH directly [40]. Lebaron et al. suggested that H2 exerts a hormetic-like effect as a redox adaptogen because it can exhibit pro-oxidative activities while also reducing excess oxidative stress [41].
Accordingly, H2 is considered as a potential and promising mitochondria-targeting molecule or nutrient that acts as a redox homeostasis regulator [42].
As is well known, H2 is a moderate/mild reducing agent (the standard reduction potential of H+/H2 at PH7 is −0.42, stronger than NAD+/NADH (−0.32) but weaker than acetate/acetaldehyde (−0.60)), barely able to scavenge •OH directly in a living body (Figure 2). Because mitochondria are the main sites of ROS generation and the targets of ROS, we suggest that the more important mechanism of the H2 molecule may be that it can easily enter cells and subcellular organelles, including mitochondria, to play a protective role through its strong penetration ability, subsequently activating the Keap1-Nrf2 (Kelch-1ike ECH-associated protein l, nuclear factor erythroid 2-related factor 2) antioxidant defense system to inhibit oxidative damage and improve the mitochondrial function, and, finally, improving the prevention of various diseases. H2 has been shown to significantly activate the Keap1-Nrf2 system, regulate the activities of endogenous antioxidants, and enhance the ability of cells to fight against damage [43].
4. The Mechanisms of H2 as an Nrf2 Activator
Nrf2 is a key factor in the regulation of oxidative stress which belongs to the CNC-BZIP transcription factor family. Upon normal physiological conditions, Nrf2 binds to Keap1 to form a complex present in the cytoplasm in a low-activity state [44]. When the organism is stimulated by oxidative stress or other pathological conditions, the cysteine residue of Keap1 is modified or Nrf2 is phosphorylated; then, Nrf2 is released from the complex and translocated to the nucleus, where it binds to the antioxidant response elements (AREs) sequence in the nucleus, initiating NRF2-mediated transcriptional processes to activate a series of phase II antioxidant enzymes to generate antioxidants to scavenge ROS and other harmful substances.
It is reported that Nrf2 can be activated in various ways, among which the Keap1-Nrf2 pathway is the most classical Nrf2 activation pathway. Keap1 contains multiple oxidative stress response sensor proteins which have different physiological functions in response to different forms of stress. To date, several studies have demonstrated that H2 activated Nrf2 through the Keap1-Nrf2 system [45,46], but the clear mechanism of the activation is not known.
Nrf2 inducers are diverse; most are electrophilic and readily react with Keap1 through the cysteine thiol groups. Among them, Cys151/Cys273/Cys288 plays a fundamental role in the perception of electrophilic Nrf2-inducing chemicals. Therefore, Nrf2 inducers have been divided into different categories based on the different cysteine residues of Keap1 they react with (Table 1). The first class specifically targets the Cys151 sensor, such as medically relevant bardoxolone methyl. Bardoxolone methyl acts as an electrophilic inducer of Nrf2 that forms a covalent interaction with the Cys151 residue of Keap1, thereby inhibiting Nrf2 ubiquitination. In mice, the Cys151 point mutation in Keap1 eliminated Nrf2 signaling and the hepatoprotective effect of bardoxolone methyl in vivo [47]. The second class of inducer targets Cys288, and 15-deoxy-prostaglandin J2 (15d-PGJ2) has been identified in this group. 15d-PGJ2, one of the endogenous Nrf2 inducers synthesized from arachidonic acid, forms a covalent compound with Keap1 to compete for the Keap1-Nrf2 binding. Class III inducers, such as 4-hydroxynonenal (4-HNE), target Cys151/Cys273/Cys288. Mass spectrometry analysis revealed that 4-HNE directly modifies cysteine residues on Keap1 and deregulates its inhibition of Nrf2 by inhibiting Keap1, further increasing the expression levels of Nrf2 target genes (e.g., TXNRD1, thioredoxin reductase-1) [48]. Indeed, Nrf2 activation was significantly reduced when Cys151 was mutated, whereas Nrf2-induced target gene activation was only slightly affected when Cys273 and Cys288 residues were mutated [49,50].
In addition, we classify the electrophilic compound that activated Nrf2 on the cysteine residues other than Cys151/Cys273/Cys288 as Class IV. The compounds of this group include, for example, Pubescenoside A, which acts on Cys77/Cys434.
Moreover, several inducers activate Nrf2 in a more complex way than the previously identified electrophilic sensors that bind to Cys226, Cys613, Cys622, and Cys624. We classify them as Class V. Hydrogen peroxide (H2O2), a key ROS molecule important in cellular physiology, is representative of this classification. Suzuki et al. revealed that Keap1 uses cysteine residues to create a special mechanism to make a disulfide bond between any combination of Cys226, Cys613, Cys622, and Cys624 to sense H2O2 [51]. This sensing mechanism is different from that used by the electrophilic Nrf2 inducer.
There is also a type of inducer that does not act through the cysteine of Keap1; these have been classified as a Class VI, and they directly inhibit the interaction between Keap1 and Nrf2 and include non-electrophilic protein–protein interaction inhibitors (PPIs) [52]. Horie et al. suggested that Keap1 binding to Nrf2 is a “hinge and latch model”, with PPIs actively using a hinge-locking mechanism, whereas electrophilic Nrf2 activators do not use this mechanism when activating Nrf2 [53].
The mechanism of Nrf2 activation by H2 seems different from the mechanism of perception of electrophilic Nrf2 inducers but may be closer to the mechanism of Class V and VI (Figure 3). As the smallest and one of the simplest molecules, H2 molecules have the capacity to pass through the Keap1 and Nrf2 binding structure to inhibit the interaction between Keap1 and Nrf2, playing the role of Class VI activators [51]. It has also been suggested that the mechanism by which H2 activates Nrf2 may be similar to that of H2O2 (Class V), either by promoting mitochondrial respiratory activity, resulting in inducing excess ROS, or by opening the mitochondrial-(ATP) K+ channel to generate ROS, which then oxidizes intracellular Keap1, releasing Nrf2 [54].
Notably, recent studies have pointed out that the oxidized form of iron porphyrin bound to the OH group is considered to be a redox-related biosensor for H2, buffering the high oxidative electrophilicity of •OH. When the originally oxidizing and deleterious electrophilic properties of •OH are mitigated, the resulting electrophilic potency may activate Nrf2, with an effect similar to that of the hormone. However, this viewpoint is brand new and still needs further verification [55].
To date, the activation of Nrf2 and its mediated antioxidant enzyme system by H2 has been reported in a variety of tissue-associated diseases, including brain, lung, liver, heart, ovary, and kidney diseases [45,56,57]. In Nrf2-deficient mice, the ability of H2 against oxidative stress in the lung was significantly diminished [58].
The results of studies of neuroblastoma cells showed that exposure of SH-SY5Y cells to H2 increased the production of mitochondrial superoxide. This process was accompanied by Nrf2 nucleus translocation as well as increased expression of Nrf2-regulated antioxidant enzymes, suggesting that H2 alleviates mitochondrial oxidative stress through activating Nrf2 [59]. Inhaled H2 also reduces neuroinflammation in memory-related regions through increasing Nrf2 protein expression in a sepsis-induced blood–brain barrier impairment and memory dysfunction [60,61]. Interestingly, one of the studies we were involved in reported that H2 (2–4%) protected against delayed encephalopathy after acute carbon monoxide poisoning, and this protective effect was related to the involvement of Nrf2 and its mediated phase II enzyme system [62].
Similar results were obtained in the lung from a seawater instillation-induced acute lung injury rabbit or from cecal ligation and puncture-induced sepsis mice, which proved that H2 could regulate the expression of heme oxygenase-1 (HO-1), the Nrf2 downstream antioxidant protein [63,64]. Inhaled H2 significantly alleviated the drop in blood O2 during hyperoxic exposure, remitted lung inflammation, and upregulated HO-1 expression. In a sepsis-induced acute lung injury model, H2 molecules inhibited high-mobility group protein1 (HMGB1) expression by activating the Nrf2-HO-1 pathway [65,66]. The latest research has revealed that H2 also affected COVID-19-induced lung injury via Nrf2 [67].
Sun et al. [46] demonstrated that the administration of H2 reduced oxidative stress in LPS-treated mice livers through activation of the Keap1-Nrf2 system. Moreover, Liu et al. [56] reported that H2 improved lipid accumulation by modulating the miR-136/MEG3/Nrf2 pathway in non-alcoholic fatty liver disease.
In an ischemia model induced in the H9C2 cell line, a H2 gas-rich medium reduced the production of •OH, promoted Nrf2 nuclear translocation, and regulated the Nrf2-HO-1 pathway, suggesting that H2 can preserve ischemic cardiomyocytes by stimulating the Nrf2 pathway [68]. H2 ameliorated LPS-injured HUVECs and inflammatory responses through Nrf2 and its downstream protein HO-1 [69].
In a long-term cyclosporine A (CsA)-induced nephrotoxicity model, HRW reduced ROS and MDA levels, increased the activities of GSH and SOD, and then improved the vascular and renal functions of rats with renal damage. Meanwhile, HRW significantly decreased the level of Keap1 while increasing the expression of Nrf2, NADPH dehydrogenase quinone1, and HO-1, suggesting that HRW restored the balance of the redox state and improved CsA-induced renal function by activating the Keap1-Nrf2 signaling pathway [45].
In a rat model with ovarian injury induced by cisplatin, HRS recovered the activity of SOD and catalase, reduced MDA levels in serum and ovarian tissues, as well as increased ovarian Nrf2 expression [70]. Inhalation of 2% H2 also attenuated severe sepsis-induced intestinal injury by modulating HO-1 and HMGB1 release in mice [71].
5. The Medical Effects of H2: Focus on the Effect on Mitochondria
A great number of basic and clinical studies have found that H2 is an important physiological regulator that protects against tissue-related diseases, such as those of the lung, heart, central nervous system, kidney, pancreas, etc., through protective effects such as antioxidant, anti-inflammatory, and antiapoptotic effects. Mitochondrial dysfunction is closely related to disease development [36]. In this section, we focus on the effects of H2 on mitochondrial function in different diseases.
5.1. Effects of H2 on Respiratory System Diseases
To date, molecular H2 has been reported to have positive effects in the prevention and treatment of acute lung injury, chronic obstructive pulmonary disease, asthma, and pulmonary hypertension [67]. Of interest, the National Health Commission of China (NHC 7th Edition Trial: Beijing, 2020) and the Chinese Centre for Disease Control and Prevention (CDCP 6th Edition Trial: Beijing, 2020) recommend effective O2 therapy as one of the modalities for the general treatment of patients with COVID-19. They also noted that inhalation of a mixture of molecular H2 and O2 (66.6% H2 & 33.3% O2) is more effective than inhalation of O2 alone [72]. The research in our lab showed that H2 enriched and that oxygenated saline inhibited LPS-induced lung injury in C57BL/6 mice through the NF-κB/NLRP3 signaling pathway. H2 demonstrated a more significant effect in inflammatory and antiapoptotic mechanisms, while O2 enhanced the hypoxic effect of the organism, with the combined protective effect of the two gases being better than their respective effects [39].
Inhalation of 2% H2 improves mitochondria function through increased mitochondrial-membrane potential and ATP levels and promotes the activity of mitochondrial-respiration complex I and complex II. H2 also regulates mitochondria dynamics, which decreases the expression of mitochondria fission protein Drp1 but increases the expression of mitochondria fusion protein mitofusin-2 (MFN2) [73].
Post-transplant morbidities, such as graft ischemia-reperfusion damage and graft-versus-host disease, are key challenges in transplantation. H2 acted as a prophylactic agent against post-transplant complications in several animal models of organ transplantation [74]. In a rat lung transplantation model, the combination of mechanical ventilation and prolonged cold ischemia resulted in a significant reduction of gas exchange in rat lung tissue (treatment with 98% O2 plus 2% nitrogen), while treatment with 98% O2 plus 2% H2 inhibited the increased tendency of pro-inflammatory cytokines and apoptotic molecules and upregulated the expression of HO-1 in the lung grafts [75]. Not only that, H2 molecules inhibited the levels of proapoptotic proteins caspase-3 and caspase-8 in lung grafts, activated the expression of antiapoptotic proteins Bcl-2 and Bcl-xL, and stabilized the mitochondrial outer membrane, preventing the release of cytochrome c into the cytosol [76]. In addition, advanced treatment of rat lung donors with H2 induces the gene expression of stress response and ATP synthesis [77].
5.2. Effects of H2 on Cardiovascular System Diseases
Molecular H2 has shown many benefits in cardiovascular disease (CVD) applications and can be used to treat a wide range of CVDs that cover ischemia-reperfusion injury, atherosclerosis, cardiac hypertrophy, radiation-induced cardiac damage, and chemotherapy-induced cardiotoxicity [80,81,82]. We evaluated the influence of inhaled H2 on heart and nerve function after cardiopulmonary resuscitation by comparing the effects of H2 inhalation in a rat model of cardiac arrest asphyxiation. The results showed that compared with O2, serum troponin T and S100B were significantly reduced after inhaling H2. In the meantime, left ventricular ejection fraction, cardiac function, and neurological function were significantly improved after H2 inhalation [82].
H2 increases autophagy by promoting autophagic flow, thereby alleviating harmful stress [83]. HRS was found to promote PINK1/Parkin-mediated autophagy, activate mitochondrial autophagy, cause damaged mitochondria to be engaged by lysosomes, and further ameliorate the inflammatory response and apoptosis induced by myocardial ischemia/reperfusion (MI/R) [84]. Feng et al. reported that HRS combined with early aerobic exercise enhances acute myocardial infarction-induced superoxide dismutase levels and total antioxidant capacity, promotes mitochondrial and DNA repair by partially regulating the expression of antioxidant-associated proteins and mitochondria-associated proteins, and protects against myocardial injury after MI [85].
HRW protects cardiac and aortic graft recipients from inflammation-related deterioration and improves allograft survival by decreasing endothelial cell proliferation, inhibiting T-cell proliferation, and reducing oxidative stress in a heterotopic heart transplantation rat model [86]. This protection mechanism also correlates with ATP levels and increases the enzyme activity of complex II, III, and V on the mitochondrial respiratory chain.
Sepsis is associated with systemic infections and inflammatory responses induced by the cardiovascular system [87]. In a sepsis-induced, myocardial-injured mouse model, molecules H2 promoted protein increase of HO-1, MFN2, and PGC1-1α expression, inhibited sepsis-induced mitochondrial dysfunction, and remodeled fatty acid oxidation in the heart in the sepsis model by increasing myocardial energy [88,89].
Oxidative stress is a major risk factor for worsening LV hypertrophy. Yu et al. found that H2 saline water improves mitochondria function by restoring electron transport chain enzyme activity, inhibiting ROS formation, and increasing ATP production in spontaneously hypertensive rats with LV hypertrophy. H2 saline water also inhibits oxidative stress, inflammatory processes, and angiotensin II [90].
Zhang et al. found that HRS treatment ameliorates vascular functional abnormalities, such as aortic hypertrophy and endothelial dysfunction, in spontaneously hypertensive rats by alleviating oxidative stress, restoring pressure receptor function, preserving mitochondrial function, and increasing NO· bioavailability [91].
5.3. Effects of H2 on Nervous System Diseases
H2 is engaged in the restoration of neurodegenerative diseases [92,93]. Research in our laboratory administered HRW to Alzheimer’s disease (AD) mice for 3 consecutive months to study its effect on cognitive function. The result showed that HRW significantly improved cognitive behaviors and ameliorated oxidative stress and inflammatory responses in the brains of female AD mice. Moreover, estrogen levels are closely related to mitochondrial function, e.g., 17β-estradiol enhances mitochondrial signaling clusters. Our results suggest that the effects of molecular H2 in female AD mice were most likely attributable to estrogen ERβ signaling [94].
Chen et al. reported that H2 treatment blocks the opening of the mitochondrial permeability transition pore in neurons. Inhalation of 75% H2 ameliorates mechanical damage to spinal cord neurons in a dose-dependent manner, significantly inhibits the production of ROS and oxidative stress markers, inhibits neuronal apoptosis, and restores mitochondrial function [25].
The results of a clinical trial on Parkinson’s disease showed that H2 significantly improved neurodegenerative symptoms with a therapeutic effect comparable to non-ergot dopamine treatment. Researchers hypothesized that this may be achieved by H2 improving cellular energy metabolism by targeting mitochondria [95]. In another experiment, H2 treatment significantly increased the levels of ATP and Δψm in neuroblastoma [57], further confirming the role of H2 in activating oxidative phospho-rylation and mitochondrial energy.
HRS improves neuronal ischemia-reperfusion by improving mitochondrial function and reducing oxidative stress [96]. Earlier studies found that H2 restored mitochondrial structural damage while reducing microRNA-210 in a hypoxia-reperfusion neural model [97]. HRS also ameliorated the activation of caspase-3, attenuated ROS accumulation, closed mitochondrial permeability transition pores, and restored mitochondrial membrane potential in isoflurane-induced, cognitively impaired mice. This suggests that HRS has the potential to attenuate anesthetic neurotoxicity [98].
5.4. Effects of H2 on Digestive System Diseases
The majority of gastrointestinal microbial species show a genetic ability to metabolize H2, which means that H2 may influence the composition of gut bacteria and modulate digestive-related diseases [99,100]. It was found that HW inhibited rat intestinal I/R-induced oxidative stress, apoptosis, and inflammation [101].
Clinical data suggest that H2 may improve glucose metabolism by interfering with the gut microbiota of impaired fasting glucose patients [102]. Another study in patients with clinical stage IV colorectal cancer found that H2 inhalation activated PGC-1α expression and enhanced mitochondrial activity, thereby reducing the proportion of PD-1 and CD8+ T cells. The reduction of these cells was associated with improved cancer prognosis [103].
5.5. Effects of H2 on Metabolic Syndrome
Mitochondrial dysfunction results in reduced mitochondrial biogenesis and increased ROS, which has been involved in the pathogenesis of a number of metabolic diseases, including diabetes and obesity. It has been widely demonstrated that H2 can scavenge ROS directly by inhibiting ROS production or indirectly by enhancing antioxidant enzyme activity, suggesting that this may be contributing to the improved mitochondrial function in metabolic disorders. Numerous studies have proven the protective effects of H2 on metabolic syndrome, which include lowering total cholesterol, total triglycerides (TG), and low-density lipoprotein (LDL) [104], reducing serum glucose and insulin levels in mice [105], as well as modifying adiposity and body weight in db/db obese mice [106]. The protective effect of H2 on diabetes and its complications may be associated with the inhibition of oxidative stress, inflammation, apoptosis, activation of the mitochondrial ATP-sensitive potassium (Mito-K-ATP) pathway, etc. [107].
Ma et al. proved that H2 promotes fatty acid oxidation by transporting fatty acids to mitochondria and subsequent catabolism to ketone bodies in rats [108]. A clinical study evaluated the effects of H2 supplementation in ten middle-aged overweight women on the indicators such as hormonal status and mitochondrial function. The results showed a significant decrease in body fat, arm fat index, serum TG, and insulin levels after 4 weeks of oral administration of H2-generating minerals. Fasting blood lactate accumulation reflects mitochondrial dysfunction which, in turn, affects the risk of metabolic diseases. After H2 intervention lasting 4 weeks, blood lactate levels were significantly lower than those in the placebo group, implying that the improvement in mitochondrial function may be related to the anti-obesity effect of H2 [109]. However, due to the small number of subjects in this study, the reliability of this result is limited, a long-term large-scale trial is needed to further verify the improvement of H2 on obesity.
Another clinical study in our lab suggests that H2 may have a potentially beneficial effect on glucose metabolism by interfering with the gut microbiota of individuals with impaired fasting glucose. Not only that, HRW may play an important role in reducing body fat and reducing fatty liver. This suggests its potential as a therapeutic intervention to improve lipid metabolism and liver health [102].
5.6. The Others
H2 restores mitochondrial oxidoreductase activity while preventing the downward trend of mitochondrial membrane potential. It ameliorated tertbutyl hydroperoxide-induced THP-1 (human acute monocytic leukemia cell line) cytotoxicity by inhibiting fatty acid peroxidation and mitochondrial dysfunction [110].
Mikako et al. reported a 12-week double-blind trial of five patients with progressive muscular dystrophy (PMD), four patients with polymyositis/dermatomyositis (PM/DM), and five patients with mitochondrial myopathy (MM), in which the patients consumed 1.0 L of HRW per day, and 18 serum markers were measured every four weeks. The results showed a significant improvement in lactate levels in the MM patients after drinking HRW. The lactate-to-pyruvate ratio in patients with DM also showed a favorable response [28].
6. Conclusions and Perspectives
In conclusion, H2 medicine has risen as a bright star in gas medicine, but it faces a few problems. Firstly, in the H2 basic research area, although a large number of H2 medicine-related studies have been carried out, the mechanisms of H2 effects are quite controversial. People do not have a high level of awareness of H2 and doubts still exist about the efficacy and safety of H2. Therefore, more specific and clear mechanisms need to be clarified. This requires more outstanding scientists to join and expend greater efforts. This review attempts to challenge the view that H2 is a selective •OH scavenger by proposing that H2 is a mitochondria-targeting molecule/nutrient via activating the Keap1-Nrf2 antioxidant system. Of course, this is quite a premature idea and needs more and further investigations to test and challenge.
Secondly, in the H2 industry, the market demand for H2 health products is insufficient. There are still many technical bottlenecks in the H2 medicine industry, such as low efficiency of H2 preparation and high storage and transport costs. In addition, the industrial chain of H2 medicine is incomplete and lacks the development of relevant standards. The H2 health industry involves a number of links, such as H2 preparation, storage, and transport; H2 generators; H2 testing; etc. At present, these links have not formed a complete industrial chain; the connection between the links is not smooth enough. Due to the lack of complete and well-defined standards, the H2 industry chain is difficult to regulate with high quality.
Thirdly, there is insufficient policy support for H2 medicine. While the H2 health industry has a great potential for development, the current government support for the H2 health industry is insufficient, and there are some deficiencies in the policy support; e.g., there is a lack of clear policy planning and support measures. Therefore, the market prospect of the H2 medicine industry is promising and urgently needs to be promoted.
Author Contributions
J.L. (Jiankang Liu) and L.Z. conceived and supervised the work; D.C. wrote the first draft of the manuscript and drew the illustrations; J.L. (Jiangang Long), L.Z. and J.L. (Jiankang Liu) reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Integrated Project of Major Research Plan of National Natural Science Foundation of China 92249303 and the General Projects of National Natural Science Foundation of China 32171102 and 31770917.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not report any data. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wilson, J.W.; Shakir, D.; Batie, M.; Frost, M.; Rocha, S. Oxygen-sensing mechanisms in cells. FEBS J. 2020, 287, 3888–3906. [Google Scholar] [CrossRef]
- Yu, B.; Ichinose, F.; Bloch, D.B.; Zapol, W.M. Inhaled nitric oxide. Br. J. Pharmacol. 2019, 176, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Roccarina, D.; Lauritano, E.C.; Gabrielli, M.; Franceschi, F.; Ojetti, V.; Gasbarrini, A. The role of methane in intestinal diseases. Am. J. Gastroenterol. 2010, 105, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.; Sethi, J.; Choi, A.M. Carbon monoxide-dependent signaling. Crit. Care Med. 2002, 30, S12–S17. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Motl, N.; Banerjee, R. H2S and its role in redox signaling. Biochim. Biophys. Acta 2014, 1844, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.F.; Sun, X.J.; Xia, Z.F. Hydrogen resuscitation, a new cytoprotective approach. Clin. Exp. Pharmacol. Physiol. 2011, 38, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Chen, S.; Zhang, J.M. Hydrogen as a selective antioxidant: A review of clinical and experimental studies. J. Int. Med. Res. 2010, 38, 1893–1903. [Google Scholar] [CrossRef] [PubMed]
- Levitt, M.D. Production and excretion of hydrogen gas in man. N. Engl. J. Med. 1969, 281, 122–127. [Google Scholar] [CrossRef]
- Du, D.; Zhao, L.; Shen, M.; Noda, M.; Qin, S.; Long, J.; Sun, X.; Liu, J. Hydrogen medicine: A rising star in gas medicine. Tradit. Med. Mod. Med. 2020, 3, 153–161. [Google Scholar] [CrossRef]
- Dole, M.; Wilson, F.R.; Fife, W.P. Hyperbaric hydrogen therapy: A possible treatment for cancer. Science 1975, 190, 152–154. [Google Scholar] [CrossRef]
- Du, Y.; Li, P.; Zhao, X. Experimental Confirmation on the Role of Hydrogen for the Meaning of Life. J. Zibo Norm. Coll. 2013, 1, 79–80. [Google Scholar]
- Du, Y.; Wei, C. New scientific topics—A preliminary study on the significance of hydrogen in life activities.ity of qi and its application prospect. J. Shandong Norm. Univ. 1999, 2, 196–197. [Google Scholar]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selec.tively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kajiyama, S.; Amano, A.; Kondo, Y.; Sasaki, T.; Handa, S.; Takahashi, R.; Fukui, M.; Hasegawa, G.; Nakamura, N.; et al. Hydrogen-rich pure water prevents superoxide formation in brain slices of vitamin C-depleted SMP30/GNL knockout mice. Biochem. Biophys. Res. Commun. 2008, 375, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Kang, Z.; Cai, J.; Liu, W.; Liu, Y.; Zhang, J.H.; Denoble, P.J.; Tao, H.; Sun, X. Hydrogen-rich saline protects myocardium against ischemia/reperfusion injury in rats. Exp. Biol. Med. 2009, 234, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Kajiya, M.; Sato, K.; Silva, M.J.; Ouhara, K.; Do, P.M.; Shanmugam, K.T.; Kawai, T. Hydrogen from intestinal bacteria is protective for Concanavalin A-induced hepatitis. Biochem. Biophys. Res. Commun. 2009, 386, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Kissner, R.; Koppenol, W.H. ONOOH does not react with H2: Potential beneficial effects of H2 as an antioxidant by selective reaction with hydroxyl radicals and peroxynitrite. Free Radic. Biol. Med. 2014, 75, 191–194. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, Y.; Bian, Y.; Li, Y.; Liu, L.; Zhang, H.; Xie, K.; Wang, G.; Yu, Y. Hydrogen Gas Protects against Intestinal Injury in Wild Type but not NRF2 Knockout Mice with Severe Sepsis by Regulating HO-1 and HMGB1 Release. Shock 2017, 48, 364–370. [Google Scholar] [CrossRef]
- Hyspler, R.; Ticha, A.; Schierbeek, H.; Galkin, A.; Zadak, Z. The Evaluation and Quantitation of Dihydrogen Metabolism Using Deuterium Isotope in Rats. PLoS ONE 2015, 10, e0130687. [Google Scholar] [CrossRef]
- Itoh, T.; Fujita, Y.; Ito, M.; Masuda, A.; Ohno, K.; Ichihara, M.; Kojima, T.; Nozawa, Y.; Ito, M. Molecular hydrogen suppresses FcepsilonRI-mediated signal transduction and prevents degranulation of mast cells. Biochem. Biophys. Res. Commun. 2009, 389, 651–656. [Google Scholar] [CrossRef]
- Yang, M.; Dong, Y.; He, Q.; Zhu, P.; Zhuang, Q.; Shen, J.; Zhang, X.; Zhao, M. Hydrogen: A Novel Option in Human Disease Treatment. Oxidative Med. Cell. Longev. 2020, 2020, 8384742. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, B.; Wu, F.; Xue, Y.; Qin, S. Progress in clinical research on the use of hydrogen molecules in preventive health care. Chin. J. Geriatr. Care 2023, 21, 117–122. [Google Scholar]
- Liu, M.Y.; Xie, F.; Zhang, Y.; Wang, T.T.; Ma, S.N.; Zhao, P.X.; Zhang, X.; Lebaron, T.W.; Yan, X.L.; Ma, X.M. Molecular hydrogen suppresses glioblastoma growth via inducing the glioma stem-like cell differentiation. Stem Cell Res. Ther. 2019, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.A.J.; Hartley, R.C.; Cochemé, H.M.; Murphy, M.P. Mitochondrial pharmacology. Trends Pharmacol. Sci. 2012, 33, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cui, J.; Zhai, X.; Zhang, J.; Gu, Z.; Zhi, X.; Weng, W.; Pan, P.; Cao, L.; Ji, F.; et al. Inhalation of Hydrogen of Different Concentrations Ameliorates Spinal Cord Injury in Mice by Protecting Spinal Cord Neurons from Apoptosis, Oxidative Injury and Mitochondrial Structure Damages. Cell Physiol. Biochem. 2018, 47, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Song, R.; Pang, Q.; Liu, Y.; Zhuang, J.; Chen, Y.; Hu, J.; Hu, J.; Liu, Y.; Liu, Z.; et al. Propofol inhibits parthanatos via ROS-ER-calcium-mitochondria signal pathway in vivo and vitro. Cell Death Dis. 2018, 9, 932. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M. Targeting molecular hydrogen to mitochondria: Barriers and gateways. Pharmacol. Res. 2015, 94, 51–53. [Google Scholar] [CrossRef]
- Ito, M.; Ibi, T.; Sahashi, K.; Ichihara, M.; Ito, M.; Ohno, K. Open-label trial and randomized, double-blind, placebo-controlled, crossover trial of hydrogen-enriched water for mitochondrial and inflammatory myopathies. Med. Gas Res. 2011, 1, 24. [Google Scholar] [CrossRef]
- Murphy, M.P.; Smith, R.A. Drug delivery to mitochondria: The key to mitochondrial medicine. Adv. Drug Deliv. Rev. 2000, 41, 235–250. [Google Scholar] [CrossRef]
- Herrmann, J.M.; Neupert, W. Protein transport into mitochondria. Curr. Opin. Microbiol. 2000, 3, 210–214. [Google Scholar] [CrossRef]
- Ohta, S. Recent progress toward hydrogen medicine: Potential of molecular hydrogen for preventive and therapeutic applications. Curr. Pharm. Des. 2011, 17, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S. Molecular hydrogen is a novel antioxidant to efficiently reduce oxidative stress with potential for the improvement of mitochondrial diseases. Biochim. Biophys. Acta BBA Gen. Subj. 2012, 1820, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Chen, X.; Ohta, S.; Sun, X. Review and prospect of the biomedical effects of hydrogen. Med. Gas Res. 2014, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ohta, S.; Zhang, J.H. Discovery of a hydrogen molecular target. Med. Gas Res. 2023, 13, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ames, B.N. Reducing mitochondrial decay with mitochondrial nutrients to delay and treat cognitive dysfunction, Alzheimer’s disease, and Parkinson’s disease. Nutr. Neurosci. 2005, 8, 67–89. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shen, W.; Zhao, B.; Wang, Y.; Wertz, K.; Weber, P.; Zhang, P. Targeting mitochondrial biogenesis for preventing and treating insulin resistance in diabetes and obesity: Hope from natural mitochondrial nutrients. Adv. Drug Deliv. Rev. 2009, 61, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Gao, P.; Shi, L.; Chen, L.; Liu, J.; Long, J. Central and Peripheral Metabolic Defects Contribute to the Pathogenesis of Alzheimer’s Disease: Targeting Mitochondria for Diagnosis and Prevention. Antioxid. Redox Signal. 2020, 32, 1188–1236. [Google Scholar] [CrossRef]
- Hou, C.; Wang, Y.; Zhu, E.; Yan, C.; Zhao, L.; Wang, X.; Qiu, Y.; Shen, H.; Sun, X.; Feng, Z.; et al. Coral calcium hydride prevents hepatic steatosis in high fat diet-induced obese rats: A potent mitochondrial nutrient and phase II enzyme inducer. Biochem. Pharmacol. 2016, 103, 85–97. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, J.; Feng, Z.; Cao, K.; Liu, J.; Xu, H. Hydrogen-rich and hyperoxygenate saline inhibits lipopolysaccharide-induced lung injury through mediating NF-κB/NLRP3 signaling pathway in C57BL/6 mice. Env. Toxicol. 2022, 37, 1575–1586. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Liu, C.; Zhou, L.; Qu, K.; Wang, R.; Tai, M.H.; Lei Lei, J.C.; Wu, Q.F.; Wang, Z.X. A review of hydrogen as a new medical therapy. Hepato-Gastroenterol. 2012, 59, 1026–1032. [Google Scholar] [CrossRef]
- LeBaron, T.W.; Laher, I.; Kura, B.; Slezak, J. Hydrogen gas: From clinical medicine to an emerging ergogenic molecule for sports athletes (1). Can. J. Physiol. Pharmacol. 2019, 97, 797–807. [Google Scholar] [CrossRef]
- Hirano, S.I.; Ichikawa, Y.; Kurokawa, R.; Takefuji, Y.; Satoh, F. A “philosophical molecule”, hydrogen may overcome senescence and intractable diseases. Med. Gas Res. 2020, 10, 47–49. [Google Scholar] [CrossRef]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, C.F.; Ping, N.N.; Sun, Y.Y.; Wang, Z.; Zhao, G.X.; Yuan, S.H.; Zibrila, A.I.; Soong, L.; Liu, J.J. Hydrogen-rich water alleviates cyclosporine A-induced nephrotoxicity via the Keap1/Nrf2 signaling pathway. J. Biochem. Mol. Toxicol. 2020, 34, e22467. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhao, N.; Wang, Y.; Su, Y.; Zhang, J.; Wang, Y.; Yu, Y.; Wang, G.; Wang, Z.; Xie, K. High concentration of hydrogen gas alleviates Lipopolysaccharide-induced lung injury via activating Nrf2 signaling pathway in mice. Int. Immunopharmacol. 2021, 101, 108198. [Google Scholar] [CrossRef] [PubMed]
- Gatbonton-Schwager, T.; Yagishita, Y.; Joshi, T.; Wakabayashi, N.; Srinivasan, H.; Suzuki, T.; Yamamoto, M.; Kensler, T.W. A Point Mutation at C151 of Keap1 of Mice Abrogates NRF2 Signaling, Cytoprotection in Vitro, and Hepatoprotection in Vivo by Bardoxolone Methyl (CDDO-Me). Mol. Pharmacol. 2023, 104, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Zhang, G.; Zheng, Y.; Yang, Y.; Chen, C.; Xia, J.; Liang, L.; Lei, C.; Hu, Y.; Cai, X.; et al. SLC27A5 deficiency activates NRF2/TXNRD1 pathway by increased lipid peroxidation in HCC. Cell Death Differ. 2020, 27, 1086–1104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D.; Hannink, M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol. Cell. Biol. 2003, 23, 8137–8151. [Google Scholar] [CrossRef]
- Shin, J.W.; Chun, K.S.; Kim, D.H.; Kim, S.J.; Kim, S.H.; Cho, N.C.; Na, H.K.; Surh, Y.J. Curcumin induces stabilization of Nrf2 protein through Keap1 cysteine modification. Biochem. Pharmacol. 2020, 173, 113820. [Google Scholar] [CrossRef]
- Suzuki, T.; Muramatsu, A.; Saito, R.; Iso, T.; Shibata, T.; Kuwata, K.; Kawaguchi, S.I.; Iwawaki, T.; Adachi, S.; Suda, H.; et al. Molecular Mechanism of Cellular Oxidative Stress Sensing by Keap1. Cell Rep. 2019, 28, 746–758.e744. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell. Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Horie, Y.; Suzuki, T.; Inoue, J.; Iso, T.; Wells, G.; Moore, T.W.; Mizushima, T.; Dinkova-Kostova, A.T.; Kasai, T.; Kamei, T.; et al. Molecular basis for the disruption of Keap1-Nrf2 interaction via Hinge & Latch mechanism. Commun. Biol. 2021, 4, 576. [Google Scholar] [CrossRef]
- Ohta, S. Molecular hydrogen may activate the transcription factor Nrf2 to alleviate oxidative stress through the hydrogen-targeted porphyrin. Aging Pathobiol. Ther. 2023, 5, 25–32. [Google Scholar] [CrossRef]
- Jin, Z.; Zhao, P.; Gong, W.; Ding, W.; He, Q. Fe-porphyrin: A redox-related biosensor of hydrogen molecule. Nano Res. 2023, 16, 2020–2025. [Google Scholar] [CrossRef]
- Liu, B.; Xue, J.; Zhang, M.; Wang, M.; Ma, T.; Zhao, M.; Gu, Q.; Qin, S. Hydrogen inhalation alleviates nonalcoholic fatty liver disease in metabolic syndrome rats. Mol. Med. Rep. 2020, 22, 2860–2868. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Ito, M.; Ohsawa, I. Molecular hydrogen protects against oxidative stress-induced SH-SY5Y neuroblastoma cell death through the process of mitohormesis. PLoS ONE 2017, 12, e0176992. [Google Scholar] [CrossRef]
- Kawamura, T.; Wakabayashi, N.; Shigemura, N.; Huang, C.S.; Masutani, K.; Tanaka, Y.; Noda, K.; Peng, X.; Takahashi, T.; Billiar, T.R.; et al. Hydrogen gas reduces hyperoxic lung injury via the Nrf2 pathway in vivo. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 304, L646–L656. [Google Scholar] [CrossRef]
- Iketani, M.; Sakane, I.; Fujita, Y.; Ito, M.; Ohsawa, I. H(2)-induced transient upregulation of phospholipids with suppression of energy metabolism. Med. Gas Res. 2023, 13, 133–141. [Google Scholar] [CrossRef]
- Jesus, A.A.; Passaglia, P.; Santos, B.M.; Rodrigues-Santos, I.; Flores, R.A.; Batalhão, M.E.; Stabile, A.M.; Cárnio, E.C. Chronic molecular hydrogen inhalation mitigates short and long-term memory loss in polymicrobial sepsis. Brain Res. 2020, 1739, 146857. [Google Scholar] [CrossRef]
- Yu, Y.; Feng, J.; Lian, N.; Yang, M.; Xie, K.; Wang, G.; Wang, C.; Yu, Y. Hydrogen gas alleviates blood-brain barrier impairment and cognitive dysfunction of septic mice in an Nrf2-dependent pathway. Int. Immunopharmacol. 2020, 85, 106585. [Google Scholar] [CrossRef]
- Shen, M.; Zheng, Y.; Zhu, K.; Cai, Z.; Liu, W.; Sun, X.; Liu, J.; Zhu, D. Hydrogen gas protects against delayed encephalopathy after acute carbon monoxide poisoning in a rat model. Neurol. Res. 2020, 42, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.; Zhang, S.; Wu, L.; Huan, L.; Huang, F.; Cui, Y.; Lin, Z. Hydrogen Gas Inhalation Attenuates Seawater Instillation-Induced Acute Lung Injury via the Nrf2 Pathway in Rabbits. Inflammation 2016, 39, 2029–2039. [Google Scholar] [CrossRef]
- Chen, H.G.; Xie, K.L.; Han, H.Z.; Wang, W.N.; Liu, D.Q.; Wang, G.L.; Yu, Y.H. Heme oxygenase-1 mediates the anti-inflammatory effect of molecular hydrogen in LPS-stimulated RAW 264.7 macrophages. Int. J. Surg. 2013, 11, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Yu, Y.; Pei, Y.; Hou, L.; Chen, S.; Xiong, L.; Wang, G. Protective effects of hydrogen gas on murine polymicrobial sepsis via reducing oxidative stress and HMGB1 release. Shock 2010, 34, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xie, K.; Chen, H.; Wang, G.; Yu, Y. Hydrogen gas inhibits high-mobility group box 1 release in septic mice by upregulation of heme oxygenase 1. J. Surg. Res. 2015, 196, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Fu, Z. Molecular hydrogen is a potential protective agent in the management of acute lung injury. Mol. Med. 2022, 28, 27. [Google Scholar] [CrossRef]
- Xie, Q.; Li, X.X.; Zhang, P.; Li, J.C.; Cheng, Y.; Feng, Y.L.; Huang, B.S.; Zhuo, Y.F.; Xu, G.H. Hydrogen gas protects against serum and glucose deprivation-induced myocardial injury in H9c2 cells through activation of the NF-E2-related factor 2/heme oxygenase 1 signaling pathway. Mol. Med. Rep. 2014, 10, 1143–1149. [Google Scholar] [CrossRef]
- Chen, H.; Xie, K.; Han, H.; Li, Y.; Liu, L.; Yang, T.; Yu, Y. Molecular hydrogen protects mice against polymicrobial sepsis by ameliorating endothelial dysfunction via an Nrf2/HO-1 signaling pathway. Int. Immunopharmacol. 2015, 28, 643–654. [Google Scholar] [CrossRef]
- Meng, X.; Chen, H.; Wang, G.; Yu, Y.; Xie, K. Hydrogen-rich saline attenuates chemotherapy-induced ovarian injury via regulation of oxidative stress. Exp. Ther. Med. 2015, 10, 2277–2282. [Google Scholar] [CrossRef]
- Harris, S.C.; Devendran, S.; Méndez-García, C.; Mythen, S.M.; Wright, C.L.; Fields, C.J.; Hernandez, A.G.; Cann, I.; Hylemon, P.B.; Ridlon, J.M. Bile acid oxidation by Eggerthella lenta strains C592 and DSM 2243(T). Gut Microbes 2018, 9, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Gallenga, C.E.; Tetè, G.; Caraffa, A.; Ronconi, G.; Younes, A.; Toniato, E.; Ross, R.; Kritas, S.K. How to reduce the likelihood of coronavirus-19 (CoV-19 or SARS-CoV-2) infection and lung inflammation mediated by IL-1. J. Biol. Regul. Homeost. Agents 2020, 34, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Dong, A.; Yu, Y.; Wang, Y.; Li, C.; Chen, H.; Bian, Y.; Zhang, P.; Zhao, Y.; Yu, Y.; Xie, K. Protective effects of hydrogen gas against sepsis-induced acute lung injury via regulation of mitochondrial function and dynamics. Int. Immunopharmacol. 2018, 65, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Shen, J. Hydrogen, a potential safeguard for graft-versus-host disease and graft ischemia-reperfusion injury? Clinics 2016, 71, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Huang, C.S.; Tochigi, N.; Lee, S.; Shigemura, N.; Billiar, T.R.; Okumura, M.; Nakao, A.; Toyoda, Y. Inhaled hydrogen gas therapy for prevention of lung transplant-induced ischemia/reperfusion injury in rats. Transplantation 2010, 90, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Fang, X.; Meng, C.; Xing, J.; Liu, J.; Yang, W.; Li, W.; Zhou, H. Lung inflation with hydrogen during the cold ischemia phase decreases lung graft injury in rats. Exp. Biol. Med. 2015, 240, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Shigemura, N.; Kawamura, T.; Noda, K.; Isse, K.; Stolz, D.B.; Billiar, T.R.; Toyoda, Y.; Bermudez, C.A.; Lyons-Weiler, J.; et al. Profiling molecular changes induced by hydrogen treatment of lung allografts prior to procurement. Biochem. Biophys. Res. Commun. 2012, 425, 873–879. [Google Scholar] [CrossRef]
- Hu, Q.; Zhou, Y.; Wu, S.; Wu, W.; Deng, Y.; Shao, A. Molecular hydrogen: A potential radioprotective agent. Biomed. Pharmacother. 2020, 130, 110589. [Google Scholar] [CrossRef]
- Terasaki, Y.; Ohsawa, I.; Terasaki, M.; Takahashi, M.; Kunugi, S.; Dedong, K.; Urushiyama, H.; Amenomori, S.; Kaneko-Togashi, M.; Kuwahara, N.; et al. Hydrogen therapy attenuates irradiation-induced lung damage by reducing oxidative stress. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 301, L415–L426. [Google Scholar] [CrossRef]
- Saengsin, K.; Sittiwangkul, R.; Chattipakorn, S.C.; Chattipakorn, N. Hydrogen therapy as a potential therapeutic intervention in heart disease: From the past evidence to future application. Cell. Mol. Life Sci. CMLS 2023, 80, 174. [Google Scholar] [CrossRef]
- Li, L.; Li, X.; Zhang, Z.; Liu, L.; Zhou, Y.; Liu, F. Protective Mechanism and Clinical Application of Hydrogen in Myocardial Ischemia-reperfusion Injury. Pak. J. Biol. Sci. PJBS 2020, 23, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jia, L.; Chen, B.; Zhang, L.; Liu, J.; Long, J.; Li, Y. Hydrogen Inhalation is Superior to Mild Hypothermia in Improving Cardiac Function and Neurological Outcome in an Asphyxial Cardiac Arrest Model of Rats. Shock 2016, 46, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.L.; Cheng, L.; Ren, J.D.; Fang, C.; Xiang, K.; Xu, H.T.; Tang, L.J.; Wang, T.; Tian, F.Z. Hydrogen-rich saline protects against ischemia/reperfusion injury in grafts after pancreas transplantations by reducing oxidative stress in rats. Mediat. Inflamm. 2015, 2015, 281985. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Chen, H.; Wu, Q.; Xie, K. Hydrogen-rich saline alleviates inflammation and apoptosis in myocardial I/R injury via PINK-mediated autophagy. Int. J. Mol. Med. 2019, 44, 1048–1062. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Cai, M.; Wang, X.; Zhang, J.; Tian, Z. Early Aerobic Exercise Combined with Hydrogen-Rich Saline as Preconditioning Protects Myocardial Injury Induced by Acute Myocardial Infarction in Rats. Appl. Biochem. Biotechnol. 2019, 187, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Noda, K.; Tanaka, Y.; Shigemura, N.; Kawamura, T.; Wang, Y.; Masutani, K.; Sun, X.; Toyoda, Y.; Bermudez, C.A.; Nakao, A. Hydrogen-supplemented drinking water protects cardiac allografts from inflammation-associated deterioration. Transpl. Int. Off. J. Eur. Soc. Organ. Transplant. 2012, 25, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Liu, Y.; Zhang, J. Recent Advances in Studies of Molecular Hydrogen against Sepsis. Int. J. Biol. Sci. 2019, 15, 1261–1275. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, A.; Xie, K.; Yu, Y. Protective Effects of Hydrogen on Myocardial Mitochondrial Functions in Septic Mice. Biomed. Res. Int. 2020, 2020, 1568209. [Google Scholar] [CrossRef]
- Tao, B.; Liu, L.; Wang, N.; Tong, D.; Wang, W.; Zhang, J. Hydrogen-Rich Saline Attenuates Lipopolysaccharide-Induced Heart Dysfunction by Restoring Fatty Acid Oxidation in Rats by Mitigating C-Jun N-Terminal Kinase Activation. Shock 2015, 44, 593–600. [Google Scholar] [CrossRef]
- Yu, Y.S.; Zheng, H. Chronic hydrogen-rich saline treatment reduces oxidative stress and attenuates left ventricular hypertrophy in spontaneous hypertensive rats. Mol. Cell. Biochem. 2012, 365, 233–242. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, Q.; He, B.; Xiao, J.; Wang, Z.; Sun, X. Anti-inflammatory effect of hydrogen-rich saline in a rat model of regional myocardial ischemia and reperfusion. Int. J. Cardiol. 2011, 148, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Htun, Y.; Nakamura, S.; Kusaka, T. Hydrogen and therapeutic gases for neonatal hypoxic-ischemic encephalopathy: Potential neuroprotective adjuncts in translational research. Pediatr. Res. 2021, 89, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Noda, M.; Liu, J.; Long, J. Neuroprotective and Preventative Effects of Molecular Hydrogen. Curr. Pharm. Des. 2021, 27, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Peng, Y.; Qin, C.; Fan, F.; Liu, J.; Long, J. Hydrogen-rich water improves cognitive impairment gender-dependently in APP/PS1 mice without affecting Aβ clearance. Free Radic. Res. 2018, 52, 1311–1322. [Google Scholar] [CrossRef] [PubMed]
- Yoritaka, A.; Takanashi, M.; Hirayama, M.; Nakahara, T.; Ohta, S.; Hattori, N. Pilot study of H2 therapy in Parkinson’s disease: A randomized double-blind placebo-controlled trial. Mov. Disord. Off. J. Mov. Disord. Soc. 2013, 28, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, H.; Ji, M.; Jia, M.; Chen, H.; Yang, J.; Duan, M. Hydrogen-rich saline attenuates neuronal ischemia--reperfusion injury by protecting mitochondrial function in rats. J. Surg. Res. 2014, 192, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yu, P.; Zeng, Q.; Luo, B.; Cai, S.; Hui, K.; Yu, G.; Zhu, C.; Chen, X.; Duan, M.; et al. Neuroprotective Effect of Hydrogen-Rich Saline in Global Cerebral Ischemia/Reperfusion Rats: Up-Regulated Tregs and Down-Regulated miR-21, miR-210 and NF-κB Expression. Neurochem. Res. 2016, 41, 2655–2665. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hou, L.; Chen, D.; Lin, F.; Chang, T.; Li, M.; Zhang, L.; Niu, X.; Wang, H.; Fu, S.; et al. Hydrogen-rich saline attenuates isoflurane-induced caspase-3 activation and cognitive impairment via inhibition of isoflurane-induced oxidative stress, mitochondrial dysfunction, and reduction in ATP levels. Am. J. Transl. Res. 2017, 9, 1162–1172. [Google Scholar]
- Xia, C.; Liu, W.; Zeng, D.; Zhu, L.; Sun, X.; Sun, X. Effect of hydrogen-rich water on oxidative stress, liver function, and viral load in patients with chronic hepatitis B. Clin. Transl. Sci. 2013, 6, 372–375. [Google Scholar] [CrossRef]
- Li, T.; Chiang, J.Y. Bile acid signaling in metabolic disease and drug therapy. Pharmacol. Rev. 2014, 66, 948–983. [Google Scholar] [CrossRef]
- Hu, H.L.; Gao, J.; Guo, W.J.; Zhou, F.H.; Liu, H.Y.; Su, C.C. Anti-injury effect of hydrogen-enriched water in a rat model of liver injury induced by aflatoxin B(1). Sheng Li Xue Bao 2019, 71, 725–731. [Google Scholar] [PubMed]
- Liang, B.; Shi, L.; Du, D.; Li, H.; Yi, N.; Xi, Y.; Cui, J.; Li, P.; Kang, H.; Noda, M.; et al. Hydrogen-Rich Water Ameliorates Metabolic Disorder via Modifying Gut Microbiota in Impaired Fasting Glucose Patients: A Randomized Controlled Study. Antioxidants 2023, 12, 1245. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Yu, S.; Qin, M.; Mao, Y.; Jin, L.; Che, N.; Liu, S.; Ge, R. Hydrogen-Rich Saline Ameliorates Allergic Rhinitis by Reversing the Imbalance of Th1/Th2 and Up-Regulation of CD4+CD25+Foxp3+Regulatory T Cells, Interleukin-10, and Membrane-Bound Transforming Growth Factor-β in Guinea Pigs. Inflammation 2018, 41, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Guo, A.; Han, X.; Wu, S.; Chen, C.; Luo, C.; Li, H.; Li, S.; Hei, Z. Aerosol inhalation of a hydrogen-rich solution restored septic renal function. Aging 2019, 11, 12097–12113. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, H.; Hu, J.; Gu, Y.; Shen, Z.; Xu, L.; Jia, X.; Zhang, X.; Ding, X. Hydrogen-Rich Saline Alleviates Kidney Fibrosis Following AKI and Retains Klotho Expression. Front. Pharmacol. 2017, 8, 499. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Liao, K.S.; Zhao, K.L.; Wang, W.X.; Zuo, T.; Deng, W.H.; Chen, C.; Yu, J.; Guo, W.Y.; He, X.B.; et al. Hydrogen-rich saline attenuates acute renal injury in sodium taurocholate-induced severe acute pancreatitis by inhibiting ROS and NF-κB pathway. Mediat. Inflamm. 2015, 2015, 685043. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Song, Y.; Yi, Y.; Jiang, X.; Ma, S.; Ma, C.; Li, J.; Zhanghuang, Z.; Liu, M.; Zhao, P.; et al. Therapeutic Potential of Molecular Hydrogen in Metabolic Diseases from Bench to Bedside. Pharmaceuticals 2023, 16, 541. [Google Scholar] [CrossRef]
- Adzavon, Y.M.; Xie, F.; Yi, Y.; Jiang, X.; Zhang, X.; He, J.; Zhao, P.; Liu, M.; Ma, S.; Ma, X. Long-term and daily use of molecular hydrogen induces reprogramming of liver metabolism in rats by modulating NADP/NADPH redox pathways. Sci. Rep. 2022, 12, 3904. [Google Scholar] [CrossRef]
- Korovljev, D.; Trivic, T.; Drid, P.; Ostojic, S.M. Molecular hydrogen affects body composition, metabolic profiles, and mitochondrial function in middle-aged overweight women. Ir. J. Med. Sci. 2018, 187, 85–89. [Google Scholar] [CrossRef]
- Iuchi, K.; Nishimaki, K.; Kamimura, N.; Ohta, S. Molecular hydrogen suppresses free-radical-induced cell death by mitigating fatty acid peroxidation and mitochondrial dysfunction. Can. J. Physiol. Pharmacol. 2019, 97, 999–1005. [Google Scholar] [CrossRef]
Figure 1.
The number of papers published from 1998 to 2022 about the most widely used gases (H2, CO, NO, O2, CH4, H2S) in medicine (data obtained after a keyword index search on PubMed with “gas name + medicine”).
Figure 1.
The number of papers published from 1998 to 2022 about the most widely used gases (H2, CO, NO, O2, CH4, H2S) in medicine (data obtained after a keyword index search on PubMed with “gas name + medicine”).
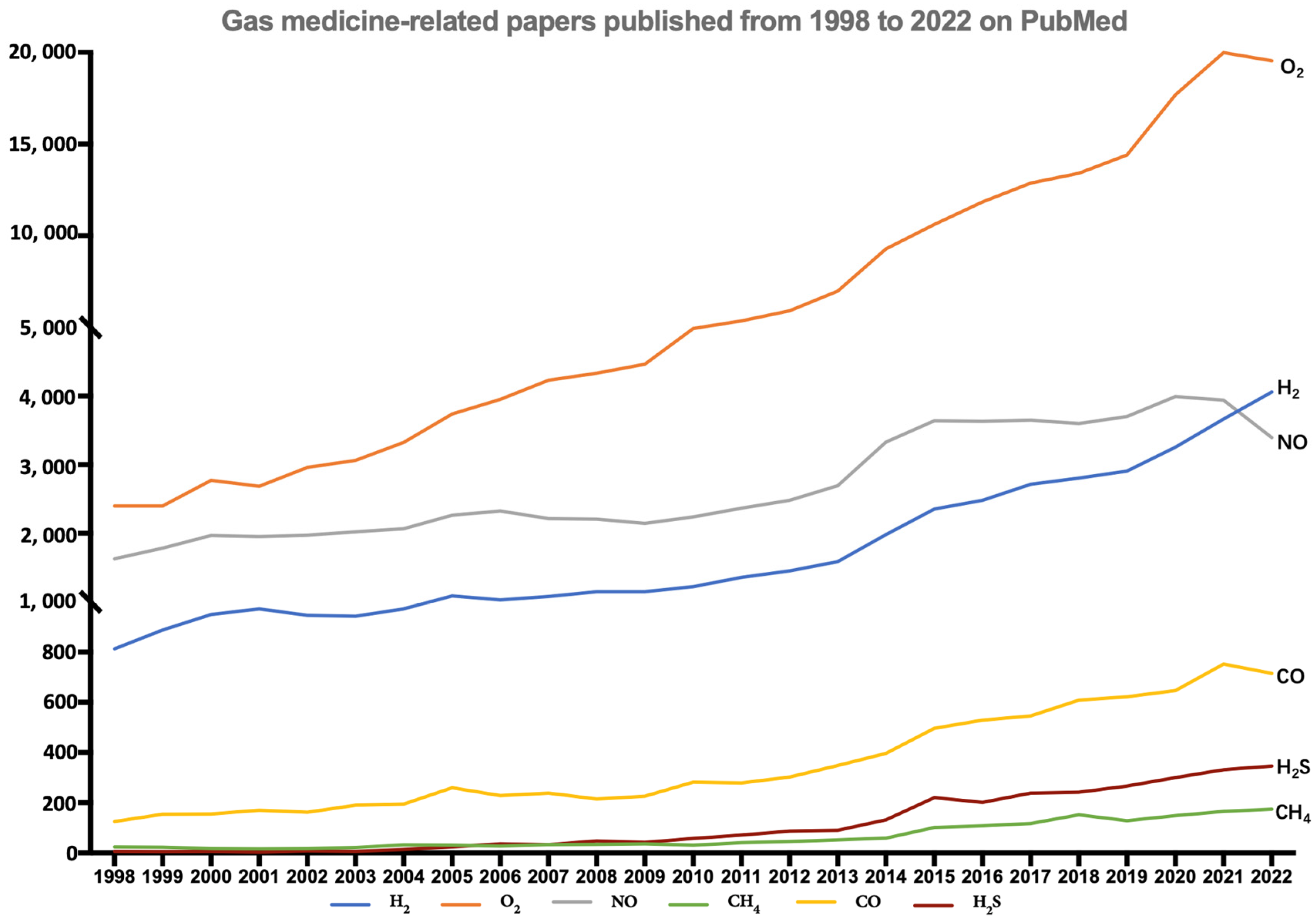
Figure 2.
Mechanisms of mitochondria-targeting by molecular H2: (1) barely reacting with •OH and ONOO− directly; (2) mainly activating Keap1-Nrf2 antioxidant systems indirectly (SOD: superoxide dismutase, CAT: catalase, GPX1: glutathione peroxidase 1, GSH: glutathione, GST: glutathione S-transferase, GPx: glutathione peroxidase, ETC: electron transfer chain, •OH: hydroxyl radical, ONOO−: nitrite peroxide, NO: nitric oxide radical).
Figure 2.
Mechanisms of mitochondria-targeting by molecular H2: (1) barely reacting with •OH and ONOO− directly; (2) mainly activating Keap1-Nrf2 antioxidant systems indirectly (SOD: superoxide dismutase, CAT: catalase, GPX1: glutathione peroxidase 1, GSH: glutathione, GST: glutathione S-transferase, GPx: glutathione peroxidase, ETC: electron transfer chain, •OH: hydroxyl radical, ONOO−: nitrite peroxide, NO: nitric oxide radical).

Figure 3.
H2 may activate Nrf2 and its mediated phase II enzyme system via non-electrophilic protein–protein interactions or by inducing the production of excess ROS to oxidized intracellular Keap1 and then activating Nrf2.
Figure 3.
H2 may activate Nrf2 and its mediated phase II enzyme system via non-electrophilic protein–protein interactions or by inducing the production of excess ROS to oxidized intracellular Keap1 and then activating Nrf2.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Classification of Nrf2 inducers targeting Keap1-Nrf2.
| Mechanism | Example | ||
|---|---|---|---|
| Class I | Electrophilic | Cys151-dependent compounds | Bardoxolone methyl Sulforaphane, dimethyl-fumarate |
| Class II | Targets Cys288 | 15d-PGJ2 | |
| Class III | Reacts with any of the three sensor cysteines Cys151/Cys273/Cys288 | 4-HNE, NaAsO2, 9-nitro-octadec-9-enoic acid | |
| Class IV | Targets cysteines Cys77/Cys434 | Pubescenoside A | |
| Class V | Non-electrophilic | Targets Cys226/Cys613/Cys622/Cys624 | H2O2, cadmium chloride, zinc chloride, prostaglandin A2 |
| Class VI | Protein–protein interaction inhibitors (PPIs) | CPUY192018 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cheng, D.; Long, J.; Zhao, L.; Liu, J. Hydrogen: A Rising Star in Gas Medicine as a Mitochondria-Targeting Nutrient via Activating Keap1-Nrf2 Antioxidant System. Antioxidants 2023, 12, 2062. https://doi.org/10.3390/antiox12122062
AMA Style
Cheng D, Long J, Zhao L, Liu J. Hydrogen: A Rising Star in Gas Medicine as a Mitochondria-Targeting Nutrient via Activating Keap1-Nrf2 Antioxidant System. Antioxidants. 2023; 12(12):2062. https://doi.org/10.3390/antiox12122062
Chicago/Turabian StyleCheng, Danyu, Jiangang Long, Lin Zhao, and Jiankang Liu. 2023. "Hydrogen: A Rising Star in Gas Medicine as a Mitochondria-Targeting Nutrient via Activating Keap1-Nrf2 Antioxidant System" Antioxidants 12, no. 12: 2062. https://doi.org/10.3390/antiox12122062
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.