
Targeted Metabolomics Analysis Suggests That Tacrolimus Alters Protection against Oxidative Stress
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Patients
2.2. Plasma Amino Acid Measurements
2.3. Pipecolic Acid Measurements
2.4. Immunosuppressant Measurements
2.5. Statistical Analysis
3. Results
3.1. Patients Characteristics
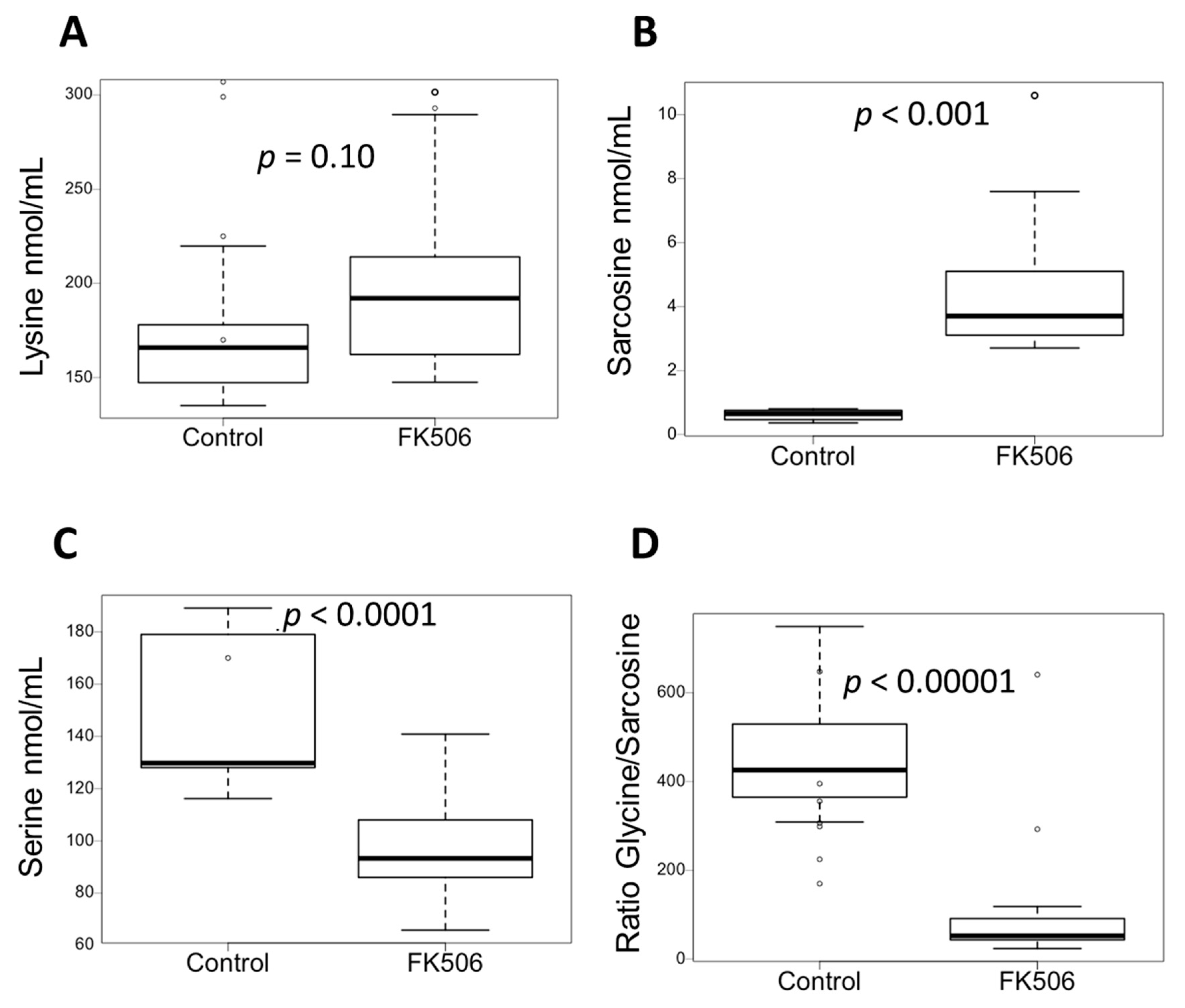
3.2. Targeted Metabolomics Showed Specific Metabolic Signature of Patient under FK506
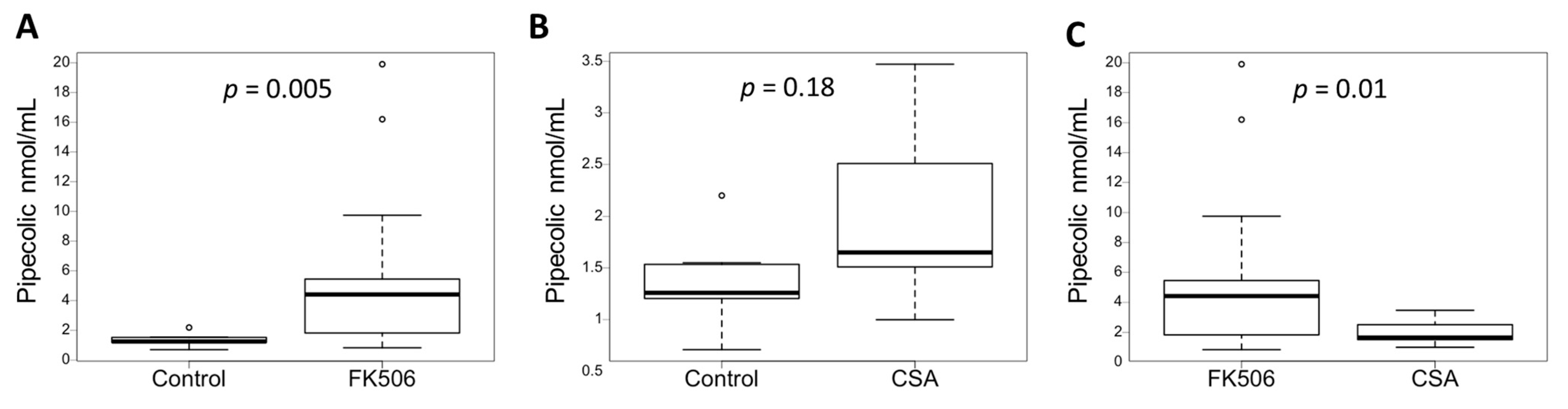
3.3. Plasma Pipecolic Acid Increase in Case of FK506 Treatment
3.4. Inhibition of the Calcineurin Pathway Does Not Increase the Concentration of Pipecolic Acid
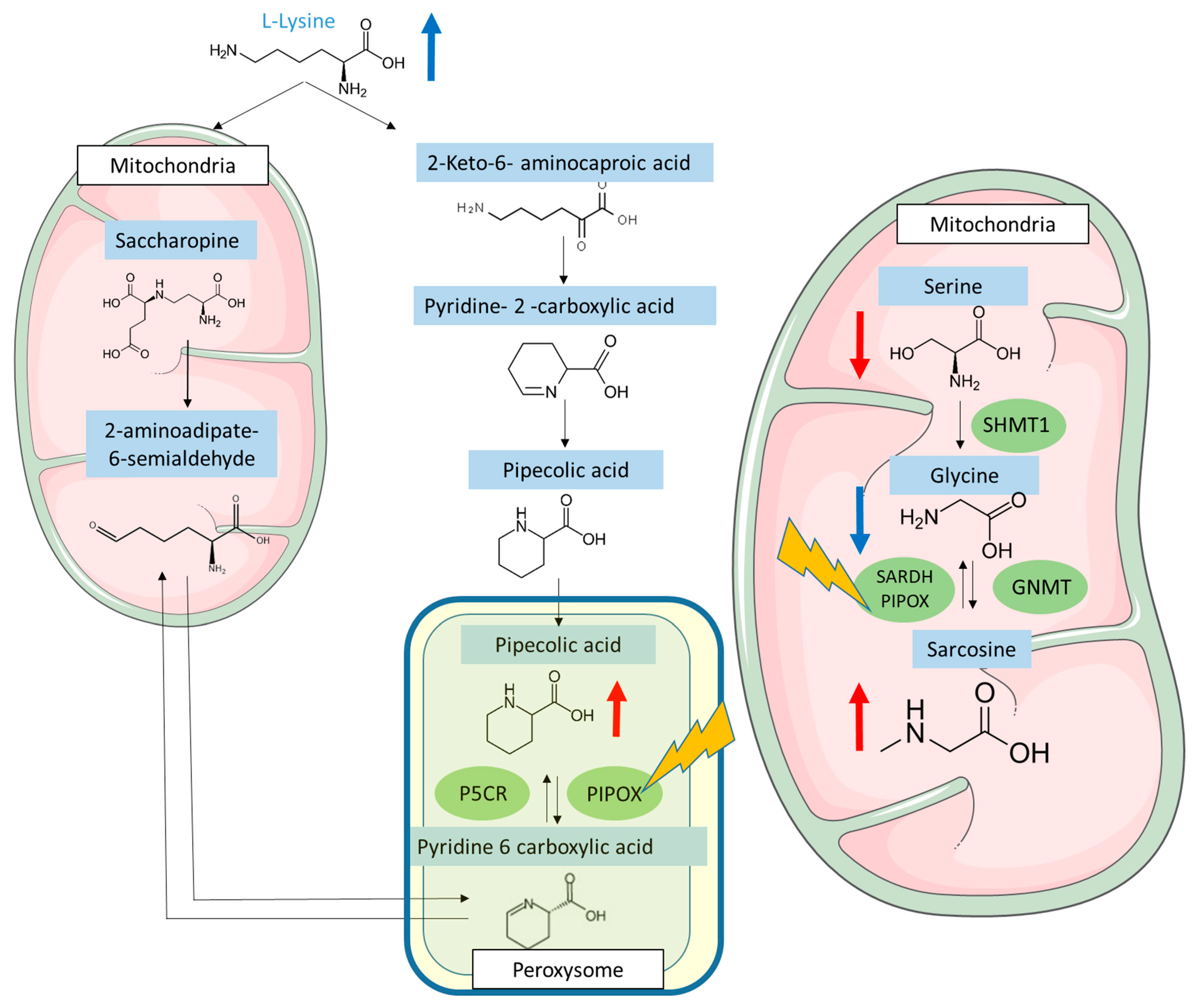
3.5. FK506 Appears to Alter the Metabolic Pathways Involving PIPOX
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallace, B.I.; Kenney, B.; Malani, P.N.; Clauw, D.J.; Nallamothu, B.K.; Waljee, A.K. Prevalence of Immunosuppressive Drug Use Among Commercially Insured US Adults, 2018–2019. JAMA Netw. Open 2021, 4, e214920. [Google Scholar] [CrossRef]
- Ho, S.; Clipstone, N.; Timmermann, L.; Northrop, J.; Graef, I.; Fiorentino, D.; Nourse, J.; Crabtree, G.R. The Mechanism of Action of Cyclosporin A and FK506. Clin. Immunol. Immunopathol. 1996, 80, S40–S45. [Google Scholar] [CrossRef]
- Koenig, J.M.; Matharoo, N.; Stegner, J.J.; Schowengerdt, K.O. Tacrolimus: In Vitro Effects on Myelopoiesis, Apoptosis, and CD11b Expression. J. Heart Lung Transplant. 2005, 24, 1332–1336. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.W.; Chung, M.W.; Choi, S.K.; Choi, S.J.; Choi, S.J.N.; Chung, S.Y. Tacrolimus-Induced Apoptosis Is Mediated by Endoplasmic Reticulum–Derived Calcium-Dependent Caspases-3,-12 in Jurkat Cells. Transplant. Proc. 2018, 50, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, H.; Tan, R.; Luo, C. Acute Pancreatitis Due to Tacrolimus in Kidney Transplant and Review of the Literature. J. Clin. Pharm. Ther. 2021, 46, 230–235. [Google Scholar] [CrossRef]
- Tory, R.; Sachs-Barrable, K.; Goshko, C.-B.; Hill, J.; Wasan, K. Tacrolimus-Induced Elevation in Plasma Triglyceride Concentrations After Administration to Renal Transplant Patients Is Partially Due to a Decrease in Lipoprotein Lipase Activity and Plasma Concentrations. Transplantation 2009, 88, 62–68. [Google Scholar] [CrossRef]
- Xia, T.; Fu, S.; Wang, Q.; Wen, Y.; Chan, S.; Zhu, S.; Gao, S.; Tao, X.; Zhang, F.; Chen, W. Targeted Metabolomic Analysis of 33 Amino Acids and Biogenic Amines in Human Urine by Ion-Pairing HPLC-MS/MS: Biomarkers for Tacrolimus Nephrotoxicity after Renal Transplantation. Biomed. Chromatogr. 2018, 32, e4198. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, M.; Xiong, X.; Du, Y.; Li, D.; Wang, Z.; Ge, W.; Zhu, Y. Plasma Metabolomic Profiling Reveals Factors Associated with Dose-Adjusted Trough Concentration of Tacrolimus in Liver Transplant Recipients. Front. Pharmacol 2022, 13, 1045843. [Google Scholar] [CrossRef]
- Illsinger, S.; Göken, C.; Brockmann, M.; Thiemann, I.; Bednarczyk, J.; Schmidt, K.-H.; Lücke, T.; Hoy, L.; Janzen, N.; Das, A.M. Effect of Tacrolimus on Energy Metabolism in Human Umbilical Endothelial Cells. Ann. Transpl. 2011, 16, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Phapale, P.B.; Kim, S.-D.; Lee, H.W.; Lim, M.; Kale, D.D.; Kim, Y.-L.; Cho, J.-H.; Hwang, D.; Yoon, Y.-R. An Integrative Approach for Identifying a Metabolic Phenotype Predictive of Individualized Pharmacokinetics of Tacrolimus. Clin. Pharmacol. Ther. 2010, 87, 426–436. [Google Scholar] [CrossRef]
- Aouad, H.; Faucher, Q.; Sauvage, F.-L.; Pinault, E.; Barrot, C.-C.; Arnion, H.; Essig, M.; Marquet, P. A Multi-Omics Investigation of Tacrolimus off-Target Effects on a Proximal Tubule Cell-Line. Pharmacol. Res. 2023, 192, 106794. [Google Scholar] [CrossRef] [PubMed]
- Borovecki, A.; Borovecki, A.; Mlinaric, A.; Mlinaric, A.; Horvat, M.; Horvat, M.; Smolcic, V.S.; Smolcic, V.S.; Smolcic, V.S. Informed Consent and Ethics Committee Approval in Laboratory Medicine. Biochem. Med. 2018, 28, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Grzych, G.; Vonghia, L.; Bout, M.-A.; Weyler, J.; Verrijken, A.; Dirinck, E.; Joncquel, M.; Van Gaal, L.; Paumelle, R.; Francque, S.; et al. Plasma BCAA Changes in Patients with NAFLD Are Sex Dependent. J. Clin. Endocrinol. Metab. 2020, 105, dgaa175. [Google Scholar] [CrossRef] [PubMed]
- Filee, R.; Schoos, R.; Boemer, F. Evaluation of Physiological Amino Acids Profiling by Tandem Mass Spectrometry. JIMD Rep. 2013, 13, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazin, C.; Guinedor, A.; Barau, C.; Gozalo, C.; Grimbert, P.; Duvoux, C.; Furlan, V.; Massias, L.; Hulin, A. Evaluation of the Architect® Tacrolimus Assay in Kidney, Liver, and Heart Transplant Recipients. J. Pharm. Biomed. Anal. 2010, 53, 997–1002. [Google Scholar] [CrossRef]
- Goldstein, B.A.; Hubbard, A.E.; Cutler, A.; Barcellos, L.F. An Application of Random Forests to a Genome-Wide Association Dataset: Methodological Considerations & New Findings. BMC Genet. 2010, 11, 49. [Google Scholar] [CrossRef] [Green Version]
- Periman, L.M.; Mah, F.S.; Karpecki, P.M. A Review of the Mechanism of Action of Cyclosporine A: The Role of Cyclosporine A in Dry Eye Disease and Recent Formulation Developments. Clin. Ophthalmol. 2020, 14, 4187–4200. [Google Scholar] [CrossRef]
- Gatto, G.J.; Boyne, M.T.; Kelleher, N.L.; Walsh, C.T. Biosynthesis of Pipecolic Acid by RapL, a Lysine Cyclodeaminase Encoded in the Rapamycin Gene Cluster. J. Am. Chem. Soc. 2006, 128, 3838–3847. [Google Scholar] [CrossRef]
- Dodt, G.; Kim, D.G.; Reimann, S.A.; Reuber, B.E.; McCabe, K.; Gould, S.J.; Mihalik, S.J. L-Pipecolic Acid Oxidase, a Human Enzyme Essential for the Degradation of L-Pipecolic Acid, Is Most Similar to the Monomeric Sarcosine Oxidases. Biochem. J. 2000, 345, 487–494. [Google Scholar] [CrossRef]
- Natarajan, S.K.; Muthukrishnan, E.; Khalimonchuk, O.; Mott, J.L.; Becker, D.F. Evidence for Pipecolate Oxidase in Mediating Protection Against Hydrogen Peroxide Stress. J. Cell. Biochem. 2017, 118, 1678–1688. [Google Scholar] [CrossRef] [Green Version]
- Jin, K.B.; Hwang, E.A.; Han, S.Y.; Park, S.B.; Kim, H.C.; Ha, E.Y.; Suh, S.I.; Mun, K.C. Effects of Tacrolimus on Antioxidant Status and Oxidative Stress in Glioma Cells. Transplant. Proc. 2008, 40, 2740–2741. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | Control | FK506 | CSA |
|---|---|---|---|
| n | 7 | 19 | 9 |
| Sex (Women/Men) | 5/2 | 7/12 | 3/6 |
| Age (years) Blood IS (ng/mL) | 36 ± 12 | 58 ± 20 | 45+/14 |
| 0 | 18.65 ± 8.48 | 681 ± 695 | |
| Type of graft (kidney/liver/heart/BM) | 0/0/0/0 | 10/2/7/0 | 0/0/3/6 |
| Plasma PA (nmol/mL) | 1.38 ± 0.46 | 5.29 ± 5.09 | 1.97 ± 0.86 |
| Plasma sarcosine (nmol/mL) | 0.60 ± 0.17 | 4.5 ± 2.1 | 2.61 ± 2.58 |
| Plasma lysine (nmol/mL) Plasma glycine (nmol/mL) | 167 ± 28 | 201 ± 50 | 184 ± 28 |
| 261 ± 45 | 234 ± 53 | 261 ± 75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joncquel, M.; Labasque, J.; Demaret, J.; Bout, M.-A.; Hamroun, A.; Hennart, B.; Tronchon, M.; Defevre, M.; Kim, I.; Kerckhove, A.; et al. Targeted Metabolomics Analysis Suggests That Tacrolimus Alters Protection against Oxidative Stress. Antioxidants 2023, 12, 1412. https://doi.org/10.3390/antiox12071412
Joncquel M, Labasque J, Demaret J, Bout M-A, Hamroun A, Hennart B, Tronchon M, Defevre M, Kim I, Kerckhove A, et al. Targeted Metabolomics Analysis Suggests That Tacrolimus Alters Protection against Oxidative Stress. Antioxidants. 2023; 12(7):1412. https://doi.org/10.3390/antiox12071412
Chicago/Turabian StyleJoncquel, Marie, Julie Labasque, Julie Demaret, Marie-Adélaïde Bout, Aghilès Hamroun, Benjamin Hennart, Mathieu Tronchon, Magali Defevre, Isabelle Kim, Alain Kerckhove, and et al. 2023. "Targeted Metabolomics Analysis Suggests That Tacrolimus Alters Protection against Oxidative Stress" Antioxidants 12, no. 7: 1412. https://doi.org/10.3390/antiox12071412