
Size Does Matter: The Influence of Bulb Size on the Phytochemical and Nutritional Profile of the Sweet Onion Landrace “Premanturska Kapula” (Allium cepa L.)

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Sample Preparation
2.3. Total Phenolic Content and Total Antioxidant Capacity
2.4. Determination of the Onion Bulb Sugar Profile
2.5. Determination of the Onion Bulb Phenolic Profile
2.6. Statistical Analysis
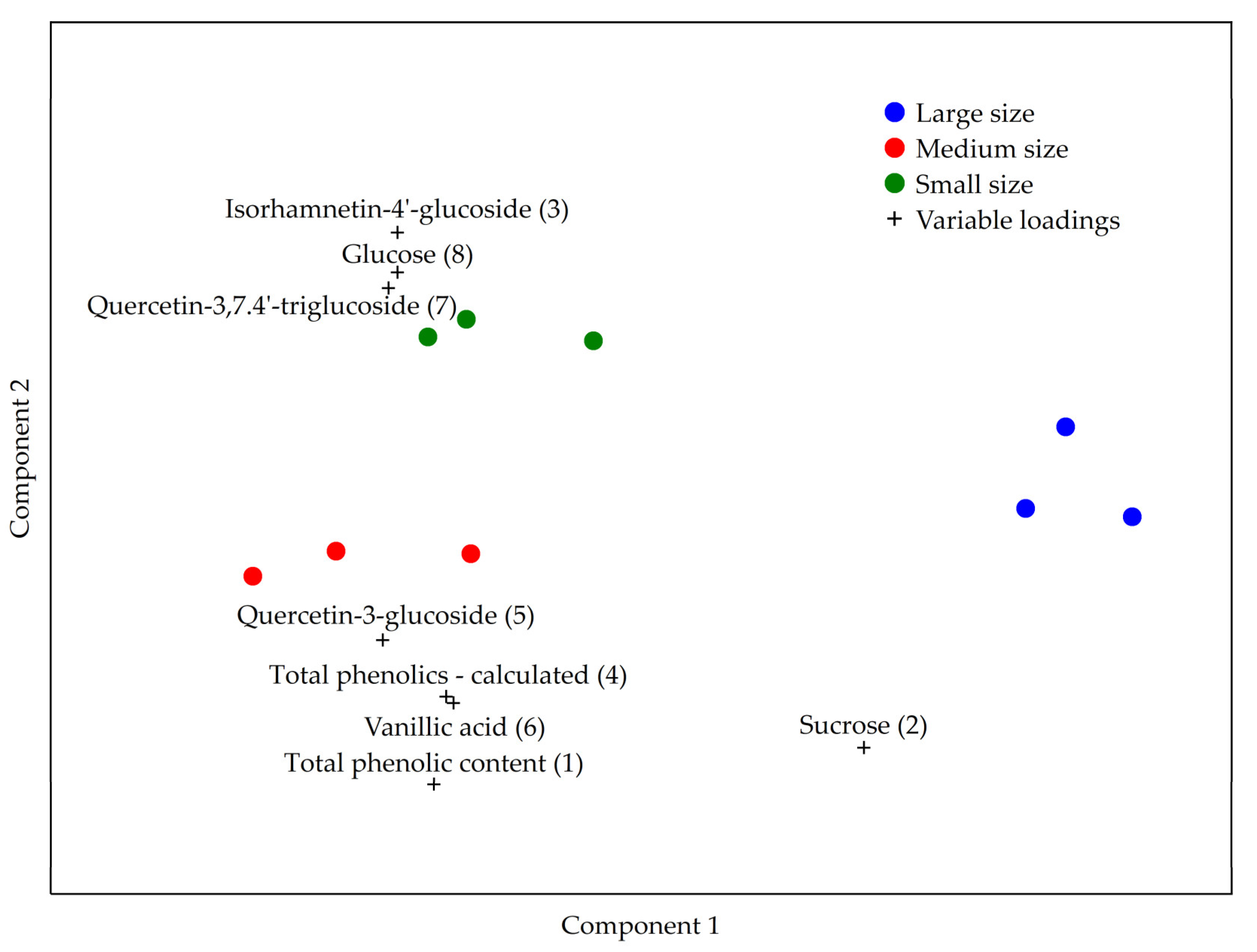
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nourbakhsh, S.S.; Cramer, C.S. Onion Plant Size Measurements as Predictors for Onion Bulb Size. Horticulturae 2022, 8, 682. [Google Scholar] [CrossRef]
- Breu, W. Allium cepa L. (Onion) Part 1: Chemistry and Analysis. Phytomedicine 1996, 3, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Suleria, H.A.R.; Butt, M.S.; Anjum, F.M.; Saeed, F.; Khalid, N. Onion: Nature Protection Against Physiological Threats. Crit. Rev. Food Sci. Nutr. 2014, 55, 50–66. [Google Scholar] [CrossRef]
- Mallor, C.; Carravedo, M.; Estopañan, G.; Mallor, F. Characterization of Genetic Resources of Onion (Allium cepa L.) from the Spanish Secondary Centre of Diversity. Span. J. Agric. Res. 2011, 9, 144–155. [Google Scholar] [CrossRef]
- Pérez, M.R.; Merkt, N.; Zikeli, S.; Zörb, C. Quality Aspects in Open-Pollinated Onion Varieties from Western Europe. J. Appl. Bot. Food Qual. 2018, 91, 69–78. [Google Scholar] [CrossRef]
- Fritsch, R.M.; Friesen, N. Evolution, Domestication and Taxonomy. In Allium Crop Science: Recent Advances; CABI Publishing: Wallingford, UK, 2002; pp. 5–30. [Google Scholar] [CrossRef]
- Vågen, I.M.; Slimestad, R. Amount of Characteristic Compounds in 15 Cultivars of Onion (Allium cepa L.) in Controlled Field Trials. J. Sci. Food Agric. 2008, 88, 404–411. [Google Scholar] [CrossRef]
- Augusti, K.T. Therapeutic and Medicinal Values of Onions and Garlic. In Onions Allied Crop; CRC Press: Boca Raton, FL, USA, 2020; pp. 93–108. [Google Scholar] [CrossRef]
- Zohri, A.N.; Abdel-Gawad, K.; Saber, S. Antibacterial, Antidermatophytic and Antitoxigenic Activities of Onion (Allium cepa L.) Oil. Microbiol. Res. 1995, 150, 167–172. [Google Scholar] [CrossRef]
- Lawande, K.E. Onion. Handb. Herbs Spices Second Ed. 2012, 1, 417–429. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, Y.; Ding, Q.; Huang, X.; Zhang, Y.; Zou, Z.; Li, M.; Cui, L.; Zhang, J. Association Mapping of Main Tomato Fruit Sugars and Organic Acids. Front. Plant Sci. 2016, 7, 1286. [Google Scholar] [CrossRef] [Green Version]
- Darbyshire, B. Changes in the Carbohydrate Content of Onion Bulbs Stored for Various Times at Different Temperatures. J. Hortic. Sci. 1978, 53, 195–201. [Google Scholar] [CrossRef]
- Shiomi, N.; Onodera, S.; Sakai, H. Fructo-Oligosaccharide Content and Fructosyltransferase Activity during Growth of Onion Bulbs. New Phytol. 1997, 136, 105–113. [Google Scholar] [CrossRef]
- Shiomi, N.; Benkeblia, N.; Onodera, S. The Metabolism of the Fructooligosaccharides in Onion Bulbs: A Comprehensive Review. J. Appl. Glycosci. 2005, 52, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Roberfroid, M. Dietary Fiber, Inulin, and Oligofructose: A Review Comparing Their Physiological Effects. Crit. Rev. Food Sci. Nutr. 1993, 33, 103–148. [Google Scholar] [CrossRef]
- Pareek, S.; Sagar, N.A.; Sharma, S.; Kumar, V. Onion (Allium cepa L.). In Fruit and Vegetable Phytochemicals: Chemistry and Human Health, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; Volume 2, pp. 1145–1162. [Google Scholar] [CrossRef]
- Van Acker, S.A.B.E.; Van Den Berg, D.J.; Tromp, M.N.J.L.; Griffioen, D.H.; Van Bennekom, W.P.; Van Der Vijgh, W.J.F.; Bast, A. Structural Aspects of Antioxidant Activity of Flavonoids. Free Radic. Biol. Med. 1996, 20, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Bystrická, J.; Musilová, J.; Vollmannová, A.; Timoracká, M.; Kavalcová, P. Bioactive Components of Onion (Allium cepa L.)—A Review. Acta Aliment. 2013, 42, 11–22. [Google Scholar] [CrossRef]
- Lanzotti, V. The Analysis of Onion and Garlic. J. Chromatogr. A 2006, 1112, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Sellappan, S.; Akoh, C.C. Flavonoids and Antioxidant Capacity of Georgia-Grown Vidalia Onions. J. Agric. Food Chem. 2002, 50, 5338–5342. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Mitchell, A.E. Quercetin and Isorhamnetin Glycosides in Onion (Allium cepa L.): Varietal Comparison, Physical Distribution, Coproduct Evaluation, and Long-Term Storage Stability. J. Agric. Food Chem. 2011, 59, 857–863. [Google Scholar] [CrossRef]
- Prakash, D.; Singh, B.N.; Upadhyay, G. Antioxidant and Free Radical Scavenging Activities of Phenols from Onion (Allium cepa). Food Chem. 2007, 102, 1389–1393. [Google Scholar] [CrossRef]
- Constant, H.L.; Slowing, K.; Graham, J.G.; Pezzuto, J.M.; Cordell, G.A.; Beecher, C.W.W. A General Method for the Dereplication of Flavonoid Glycosides Utilizing High Performance Liquid Chromatography/Mass Spectrometric Analysis. Phytochem. Anal. 1997, 8, 176–180. [Google Scholar] [CrossRef]
- Simin, N.; Orcic, D.; Cetojevic-Simin, D.; Mimica-Dukic, N.; Anackov, G.; Beara, I.; Mitic-Culafic, D.; Bozin, B. Phenolic Profile, Antioxidant, Anti-Inflammatory and Cytotoxic Activities of Small Yellow Onion (Allium flavum L. Subsp. Flavum, Alliaceae). LWT Food Sci. Technol. 2013, 54, 139–146. [Google Scholar] [CrossRef]
- Benkeblia, N. Accumulation of Total and Reducing Sugars in the Green Leaves and Bulb Tissues during Growing and Bulbing of Onion (Allium cepa L.). Acta Hortic. 2012, 932, 291–294. [Google Scholar] [CrossRef]
- Darbyshire, B.; Henry, R.J. Differences in Fructan Content and Synthesis in Some Allium Species. New Phytol. 1981, 87, 249–256. [Google Scholar] [CrossRef]
- Henriksen, K.; Hansen, S.L. Increasing the Dry Matter Production in Bulb Onions (Allium cepa L.). Acta Hortic. 2001, 555, 147–152. [Google Scholar] [CrossRef]
- Yoo, K.S.; Pike, L.; Crosby, K.; Jones, R.; Leskovar, D. Differences in Onion Pungency Due to Cultivars, Growth Environment, and Bulb Sizes. Sci. Hortic. 2006, 110, 144–149. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.U.; Lee, J.H.; Choi, S.H.; Lee, J.S.; Ohnisi-Kameyama, M.; Kozukue, N.; Levin, C.E.; Friedman, M. Flavonoid Content in Fresh, Home-Processed, and Light-Exposed Onions and in Dehydrated Commercial Onion Products. J. Agric. Food Chem. 2008, 56, 8541–8548. [Google Scholar] [CrossRef]
- Patil, B.S.; Pike, L.M.; Hamilton, B.K. Changes in Quercetin Concentration in Onion (Allium cepa L.) Owing to Location, Growth Stage and Soil Type. New Phytol. 1995, 130, 349–355. [Google Scholar] [CrossRef]
- Mogren, L.M.; Olsson, M.E.; Gertsson, U.E. Quercetin Content in Field-Cured Onions (Allium cepa L.): Effects of Cultivar, Lifting Time, and Nitrogen Fertilizer Level. J. Agric. Food Chem. 2006, 54, 6185–6191. [Google Scholar] [CrossRef]
- Liu, J.; Yu, L.L.; Wu, Y. Bioactive Components and Health Beneficial Properties of Whole Wheat Foods. J. Agric. Food Chem. 2020, 68, 12904–12915. [Google Scholar] [CrossRef]
- Ren, F.; Reilly, K.; Gaffney, M.; Kerry, J.P.; Hossain, M.; Rai, D.K. Evaluation of Polyphenolic Content and Antioxidant Activity in Two Onion Varieties Grown under Organic and Conventional Production Systems. J. Sci. Food Agric. 2017, 97, 2982–2990. [Google Scholar] [CrossRef]
- Ipgri, E.C.P.; Avrdc, G.R. Descriptors for Allium (Allium spp.); International Plant Genetic Resources Institute: Rome, Italy, 2001. [Google Scholar]
- Major, N.; Perković, J.; Palčić, I.; Bažon, I.; Horvat, I.; Ban, D.; Goreta Ban, S. The Phytochemical and Nutritional Composition of Shallot Species (Allium × Cornutum, Allium × Proliferum and A. Cepa Aggregatum) Is Genetically and Environmentally Dependent. Antioxidants 2022, 11, 1547. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Golubkina, N.; Caruso, G. Onion. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Academic Press: Cambridge, MA, USA, 2020; pp. 73–87. [Google Scholar] [CrossRef]
- Vavrina, C.S.; Smittle, D.A. Evaluating Sweet Onion Cultivars for Sugar Concentrations and Pungency. HortScience 1993, 28, 804–806. [Google Scholar] [CrossRef]
- Agricultural Research Service, U.S.D. of A. United States USDA National Nutrient Database for Standard Reference. Available online: https://fdc.nal.usda.gov/fdc-app.html#/ (accessed on 12 January 2023).
- Loredana, L.; Adiletta, G.; Nazzaro, F.; Florinda, F.; Marisa, D.M.; Donatella, A. Biochemical, Antioxidant Properties and Antimicrobial Activity of Different Onion Varieties in the Mediterranean Area. J. Food Meas. Charact. 2019, 13, 1232–1241. [Google Scholar] [CrossRef]
- Sharma, K.; Ko, E.Y.; Assefa, A.D.; Ha, S.; Nile, S.H.; Lee, E.T.; Park, S.W. Temperature-Dependent Studies on the Total Phenolics, Flavonoids, Antioxidant Activities, and Sugar Content in Six Onion Varieties. J. Food Drug Anal. 2015, 23, 243–252. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of Antioxidant Activities of Common Vegetables Employing Oxygen Radical Absorbance Capacity (ORAC) and Ferric Reducing Antioxidant Power (FRAP) Assays: A Comparative Study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef]
- Pérez-Gregorio, M.R.; Regueiro, J.; Simal-Gándara, J.; Rodrigues, A.S.; Almeida, D.P.F. Increasing the Added-Value of Onions as a Source of Antioxidant Flavonoids: A Critical Review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1050–1062. [Google Scholar] [CrossRef]
- Shon, M.Y.; Choi, S.D.; Kahng, G.G.; Nam, S.H.; Sung, N.J. Antimutagenic, Antioxidant and Free Radical Scavenging Activity of Ethyl Acetate Extracts from White, Yellow and Red Onions. Food Chem. Toxicol. 2004, 42, 659–666. [Google Scholar] [CrossRef]
- Hedges, L.; Lister, C. The Nutritional Attributes of Allium Species. Crop Food Res. Confid. Rep. 2007, 44, 1814. [Google Scholar]
- Hertog, M.G.L.; Hollman, P.C.H.; Katan, M.B. Content of Potentially Anticarcinogenic Flavonoids of 28 Vegetables and 9 Fruits Commonly Consumed in the Netherlands. J. Agric. Food Chem. 1992, 40, 2379–2383. [Google Scholar] [CrossRef]
- Cozzolino, R.; Malorni, L.; Martignetti, A.; Picariello, G.; Siano, F.; Forte, G.; De Giulio, B. Comparative Analysis of Volatile Profiles and Phenolic Compounds of Four Southern Italian Onion (Allium cepa L.) Landraces. J. Food Compos. Anal. 2021, 101, 103990. [Google Scholar] [CrossRef]
- Fenwick, G.R.; Hanley, A.B. Chemical Composition; CRC Press: Boca Raton, FL, USA, 2020; ISBN 9780429355752. [Google Scholar]
- Panisset, B.; Tissut, M. Flavonoid Metabolism in the Isolated Scales of 8 Onion Varieties—Effects of Treatments with Gibberellic-Acid and Aminozide. Physiol. Veg. 1983, 21, 49–57. [Google Scholar]
- Slimestad, R.; Fossen, T.; Vågen, I.M. Onions: A Source of Unique Dietary Flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef]
- Olsson, M.E.; Gustavsson, K.E.; Vågen, I.M. Quercetin and Isorhamnetin in Sweet and Red Cultivars of Onion (Allium cepa L.) at Harvest, after Field Curing, Heat Treatment, and Storage. J. Agric. Food Chem. 2010, 58, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorsi, P.; Caristi, C.; Gargiulli, C.; Leuzzi, U. Flavonol Glucosides in Allium Species: A Comparative Study by Means of HPLC–DAD–ESI-MS–MS. Food Chem. 2008, 107, 1668–1673. [Google Scholar] [CrossRef]
- Pinho, C.; Soares, M.T.; Almeida, I.F.; Aguiar, A.A.R.M.; Mansilha, C.; Ferreira, I.M.P.L.V.O. Impact of Freezing on Flavonoids/Radical-Scavenging Activity of Two Onion Varieties. Czech J. Food Sci. 2015, 33, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Gorinstein, S.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Najman, K.; Drzewiecki, J.; Cvikrová, M.; Martincová, O.; Katrich, E.; Trakhtenberg, S. Comparison of the Main Bioactive Compounds and Antioxidant Activities in Garlic and White and Red Onions after Treatment Protocols. J. Agric. Food Chem. 2008, 56, 4418–4426. [Google Scholar] [CrossRef]
- Juániz, I.; Ludwig, I.A.; Huarte, E.; Pereira-Caro, G.; Moreno-Rojas, J.M.; Cid, C.; De Peña, M.P. Influence of Heat Treatment on Antioxidant Capacity and (Poly)Phenolic Compounds of Selected Vegetables. Food Chem. 2016, 197, 466–473. [Google Scholar] [CrossRef]
- Mallor, C.; Balcells, M.; Mallor, F.; Sales, E. Genetic Variation for Bulb Size, Soluble Solids Content and Pungency in the Spanish Sweet Onion Variety Fuentes de Ebro. Response to Selection for Low Pungency. Plant Breed. 2011, 130, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, J.E.; Triggs, C.M.; De Ruiter, J.M.; Gandar, P.W. Bulbing in Onions: Photoperiod and Temperature Requirements and Prediction of Bulb Size and Maturity. Ann. Bot. 1996, 78, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.X.; Lin, F.J.; Li, H.; Li, H.B.; Wu, D.T.; Geng, F.; Ma, W.; Wang, Y.; Miao, B.H.; Gan, R.Y. Recent Advances in Bioactive Compounds, Health Functions, and Safety Concerns of Onion (Allium cepa L.). Front. Nutr. 2021, 8, 669805. [Google Scholar] [CrossRef]
- Khokhar, K.M. Environmental and Genotypic Effects on Bulb Development in Onion—A Review. J. Hortic. Sci. Biotechnol. 2017, 92, 448–454. [Google Scholar] [CrossRef]
- Sinclair, P.J.; Blakeney, A.B.; Barlow, E.W.R. Relationships Between Bulb Dry Matter Content, Soluble Solids Concentration and Non-Structural Carbohydrate Composition in the Onion (Allium cepa). J. Sci. Food Agric. 1995, 69, 203–209. [Google Scholar] [CrossRef]
- Benkeblia, N.; Onodera, S.; Yoshihira, T.; Kosaka, S.; Shiomi, N. Effect of Temperature on Soluble Invertase Activity, and Glucose, Fructose and Sucrose Status of Onion Bulbs (Allium cepa) in Store. Int. J. Food Sci. Nutr. 2009, 55, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz, S.; Kolniak-Ostek, J.; Oszmiański, J.; Wiśniewski, R. Comparison of Phenolic Content and Antioxidant Capacity of Bear Garlic (Allium ursinum L.) in Different Maturity Stages. J. Food Process. Preserv. 2017, 41, e12921. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, S.; Ha, I.; Min, B.; Hwang, H.; Lee, S. Comparison of Bulb and Leaf Quality, and Antioxidant Compounds of Intermediate-Day Onion from Organic and Conventional Systems. Hortic. Environ. Biotechnol. 2015, 56, 427–436. [Google Scholar] [CrossRef]
- Leskovar, D.I.; Cantamutto, M.; Marinangelli, P.; Gaido, E. Comparison of Direct-Seeded, Bareroot, and Various Tray Seedling Densities on Growth Dynamics and Yield of Long-Day Onion. Agronomie 2004, 24, 35–40. [Google Scholar] [CrossRef]
- Russo, V.M. Greenhouse-Grown Transplants as an Alternative to Bare-Root Transplants for Onion. HortScience 2004, 39, 1267–1271. [Google Scholar] [CrossRef] [Green Version]
- Boyhan, G.E.; Torrance, R.L.; Cook, J.; Cliff, R.; Hill, C.R. Plant Population, Transplant Size, and Variety Effect on Transplanted Short-Day Onion Production. Horttechnology 2009, 19, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, J.; Lamei, J.; Khasmakhi-Sabet, A.; Olfati, J.A.; Peyvast, G. Effect of Irrigation Methods and Transplant Size on Onion Cultivars Yield and Quality. J. Food Agric. Environ. 2010, 8, 158–160. [Google Scholar]
- Cheng, A.; Chen, X.; Jin, Q.; Wang, W.; Shi, J.; Liu, Y. Comparison of Phenolic Content and Antioxidant Capacity of Red and Yellow Onions. Czech J. Food Sci. 2013, 31, 501–508. [Google Scholar] [CrossRef]


{kind=link}
| Parameter | Average | Minimum | Maximum | Standard Deviation | |
|---|---|---|---|---|---|
| Dry matter | % | 7.33 | 7.10 | 7.55 | 0.13 |
| Soluble sugars | |||||
| Sum of sugars | g/100 g DW | 61.5 | 59.4 | 64.6 | 1.7 |
| Glucose | 28.0 | 22.5 | 31.8 | 3.0 | |
| Fructose | 21.2 | 18.6 | 23.0 | 1.3 | |
| Sucrose | 10.2 | 7.6 | 13.4 | 2.3 | |
| Fructooligosaccharides | 2.09 | 0.55 | 5.90 | 2.23 | |
| Total antioxidant capacity | |||||
| TPC | µg GAE/g DW | 3270 | 2908 | 3767 | 327 |
| DPPH | µmol TE/g DW | 3.00 | 2.00 | 4.01 | 0.66 |
| FRAP | 4.86 | 4.20 | 5.57 | 0.50 | |
| ORAC | 79.8 | 72.0 | 92.9 | 7.1 | |
| Phenolic compounds | |||||
| Total phenolics (calculated) | µg/g DW | 4572 | 3587 | 5392 | 703 |
| Quercetin-3,4′-diglucoside | 2272 | 1907 | 2768 | 306 | |
| Quercetin-4′-glucoside | 1210 | 932 | 1453 | 205 | |
| Protocatehuic acid | 729 | 502 | 934 | 151 | |
| Isorhamnetin-4′-glucoside | 271 | 176 | 342 | 62 | |
| Quercetin-3-glucoside | 50.2 | 37.5 | 63.1 | 10.2 | |
| Quercetin-3,7-diglucoside | 26.4 | 20.2 | 33.9 | 4.4 | |
| Quercetin-3,7.4′-triglucoside | 9.1 | 7.6 | 10.6 | 1.1 | |
| Vanillic acid | 2.55 | 2.03 | 3.28 | 0.44 | |
| Quercetin | 2.46 | 1.85 | 2.89 | 0.35 | |
| Bulb Size | |||||
|---|---|---|---|---|---|
| Parameter | Large | Medium | Small | p-Value | |
| Dry matter | % | 7.4 ± 0.02 | 7.28 ± 0.04 | 7.3 ± 0.13 | ns |
| Soluble sugars | |||||
| Total sugars | g/100 g DW | 62.1 ± 1.4 | 61.6 ± 0.5 | 60.7 ± 1.2 | ns |
| Glucose | 24.4 ± 1.1 b 1 | 29.4 ± 0.5 a | 30.4 ± 0.7 a | ** | |
| Fructose | 19.8 ± 0.8 | 21.7 ± 0.3 | 22 ± 0.5 | ns | |
| Sucrose | 12.9 ± 0.3 a | 9.9 ± 0.3 b | 7.7 ± 0.1 c | *** | |
| Fructooligosaccharides | 5.02 ± 0.44 a | 0.63 ± 0.07 b | 0.64 ± 0 b | *** | |
| Total antioxidant capacity | |||||
| TPC | µg GAE/g DW | 3011 ± 56 b | 3684 ± 71 a | 3116 ± 58 b | *** |
| DPPH | µmol TE/g DW | 2.25 ± 0.15 b | 3.54 ± 0.24 a | 3.21 ± 0.21 a | ** |
| FRAP | 4.41 ± 0.12 | 5.14 ± 0.29 | 5.04 ± 0.28 | ns | |
| ORAC | 76.3 ± 2.9 | 83.8 ± 6.2 | 79.2 ± 2.7 | ns | |
| Phenolic compounds | |||||
| Total phenolics (calculated) | µg/g DW | 3710 ± 84 b | 5182 ± 160 a | 4824 ± 191 a | ** |
| Quercetin-3,4′-diglucoside | 1924 ± 9 b | 2525 ± 129 a | 2366 ± 106 a | * | |
| Quercetin-4′-glucoside | 961 ± 24 b | 1392 ± 56 a | 1278 ± 49 a | ** | |
| Protocatehuic acid | 556 ± 43 b | 874 ± 44 a | 756 ± 27 a | ** | |
| Isorhamnetin-4′-glucoside | 196 ± 10 c | 284 ± 12 b | 332 ± 9 a | *** | |
| Quercetin-3-glucoside | 39.0 ± 1.3 c | 61.9 ± 0.9 a | 49.6 ± 2.1 b | *** | |
| Quercetin-3,7-diglucoside | 21.6 ± 0.8 b | 29.7 ± 2.5 a | 27.9 ± 1.2 a | * | |
| Quercetin-3,7.4′-triglucoside | 7.81 ± 0.10 b | 9.55 ± 0.42 a | 9.97 ± 0.35 a | * | |
| Vanillic acid | 2.21 ± 0.10 b | 3 ± 0.26 a | 2.44 ± 0.11 ab | * | |
| Quercetin | 2.52 ± 0.21 | 2.34 ± 0.24 | 2.52 ± 0.22 | ns | |
| L | a | b | ||
|---|---|---|---|---|
| Bulb size | Small | 58.3 ± 1.5 a 1 | 7.3 ± 1 b | 4.4 ± 1.2 b |
| Medium | 53.2 ± 1.4 b | 9.2 ± 1.3 a | 3.6 ± 1.3 b | |
| Large | 61.0 ± 1.1 a | 6.0 ± 1.0 b | 6.7 ± 1.1 a | |
| p-value | ** | *** | *** | |
| Bulb part | Outer bulb flesh | 58.0 ± 1.2 | 11.6 ± 0.7 a | −0.2 ± 0.9 c |
| Inner bulb flesh | 58.6 ± 1.3 | 2.7 ± 0.2 b | 4.3 ± 0.3 b | |
| Outer dry peel | 54.2 ± 2.2 | 12.6 ± 1.3 a | 14.3 ± 1 a | |
| p-value | ns | *** | *** | |
| Bulb size × Bulb part | ||||
| Inner bulb flesh | Small | 60.7 ± 1.8 | 3.0 ± 0.3 d | 4.5 ± 0.7 |
| Medium | 53.3 ± 2.3 | 3.3 ± 0.4 d | 3.1 ± 0.5 | |
| Large | 61.7 ± 1.7 | 1.7 ± 0.3 d | 5.4 ± 0.4 | |
| Outer bulb flesh | Small | 55.2 ± 2.3 | 12.5 ± 1 b | -0.6 ± 2 |
| Medium | 55.5 ± 1.3 | 13.8 ± 1.2 ab | -2.5 ± 0.9 | |
| Large | 63.3 ± 1.1 | 8.3 ± 0.4 c | 2.6 ± 0.8 | |
| Outer dry peel | Small | 57.4 ± 5.2 | 9.1 ± 2.5 c | 11.8 ± 2.1 |
| Medium | 49.4 ± 3.4 | 16.1 ± 1 a | 14.5 ± 0.9 | |
| Large | 55.9 ± 2.3 | 12.7 ± 2.2 b | 16.7 ± 1.6 | |
| p-value | ns | *** | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Major, N.; Išić, N.; Kovačević, T.K.; Anđelini, M.; Ban, D.; Prelac, M.; Palčić, I.; Goreta Ban, S. Size Does Matter: The Influence of Bulb Size on the Phytochemical and Nutritional Profile of the Sweet Onion Landrace “Premanturska Kapula” (Allium cepa L.). Antioxidants 2023, 12, 1596. https://doi.org/10.3390/antiox12081596
Major N, Išić N, Kovačević TK, Anđelini M, Ban D, Prelac M, Palčić I, Goreta Ban S. Size Does Matter: The Influence of Bulb Size on the Phytochemical and Nutritional Profile of the Sweet Onion Landrace “Premanturska Kapula” (Allium cepa L.). Antioxidants. 2023; 12(8):1596. https://doi.org/10.3390/antiox12081596
Chicago/Turabian StyleMajor, Nikola, Nina Išić, Tvrtko Karlo Kovačević, Magdalena Anđelini, Dean Ban, Melissa Prelac, Igor Palčić, and Smiljana Goreta Ban. 2023. "Size Does Matter: The Influence of Bulb Size on the Phytochemical and Nutritional Profile of the Sweet Onion Landrace “Premanturska Kapula” (Allium cepa L.)" Antioxidants 12, no. 8: 1596. https://doi.org/10.3390/antiox12081596
APA StyleMajor, N., Išić, N., Kovačević, T. K., Anđelini, M., Ban, D., Prelac, M., Palčić, I., & Goreta Ban, S. (2023). Size Does Matter: The Influence of Bulb Size on the Phytochemical and Nutritional Profile of the Sweet Onion Landrace “Premanturska Kapula” (Allium cepa L.). Antioxidants, 12(8), 1596. https://doi.org/10.3390/antiox12081596