
Effects of Crocus sativus L. Floral Bio-Residues Related to Skin Protection



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extract Obtention
2.3. Phytochemical Analysis
2.3.1. Total Phenolic, Flavonoid, and Anthocyanin Content
2.3.2. Determination of the Polyphenolic Profile by UHPLC-ESI(±)-QTOF-MS Analysis
2.4. In Vitro Scavenging Activity
2.4.1. DPPH Assay
2.4.2. Radical ABTS Scavenging Assay
2.4.3. Superoxide Anion Scavenging Assay
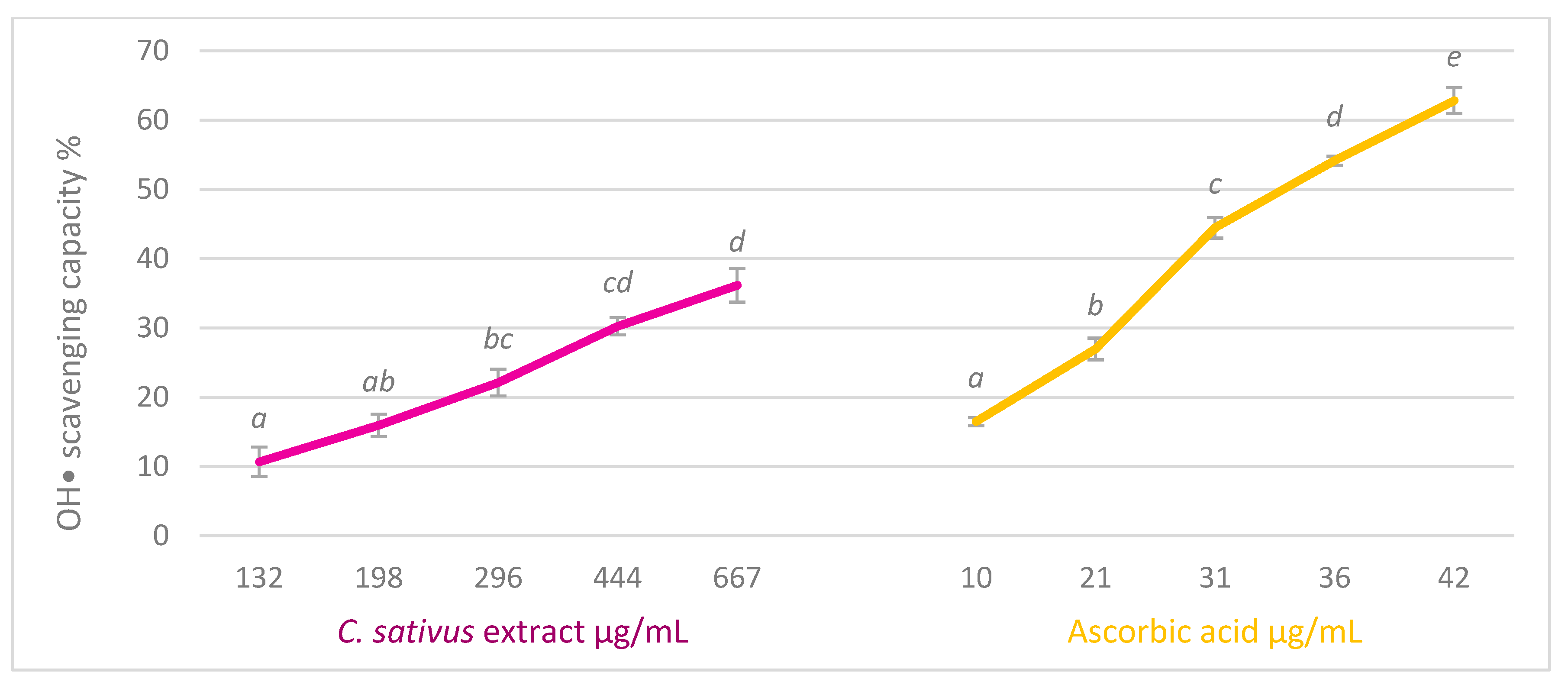
2.4.4. Hydroxyl Radical Scavenging Test
2.4.5. Nitrogen Free Radicals Scavenging Test
2.5. Radical Scavenging Activity in Cell Culture (HepG2 and Hs27)
2.5.1. Cell Culture
2.5.2. Cytotoxicity
2.5.3. Intracellular ROS Measurement
2.6. Enzymatic Activities
2.6.1. Elastase Inhibition Assay
2.6.2. Hyaluronidase Inhibition Assay
2.6.3. Collagenase Inhibition Assay
2.6.4. Tyrosinase Inhibition Assay
2.6.5. Xanthine Oxidase Inhibition Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. Chemical Analysis of the Extract
3.1.1. Total Phenolic, Flavonoid, and Anthocyanins Content
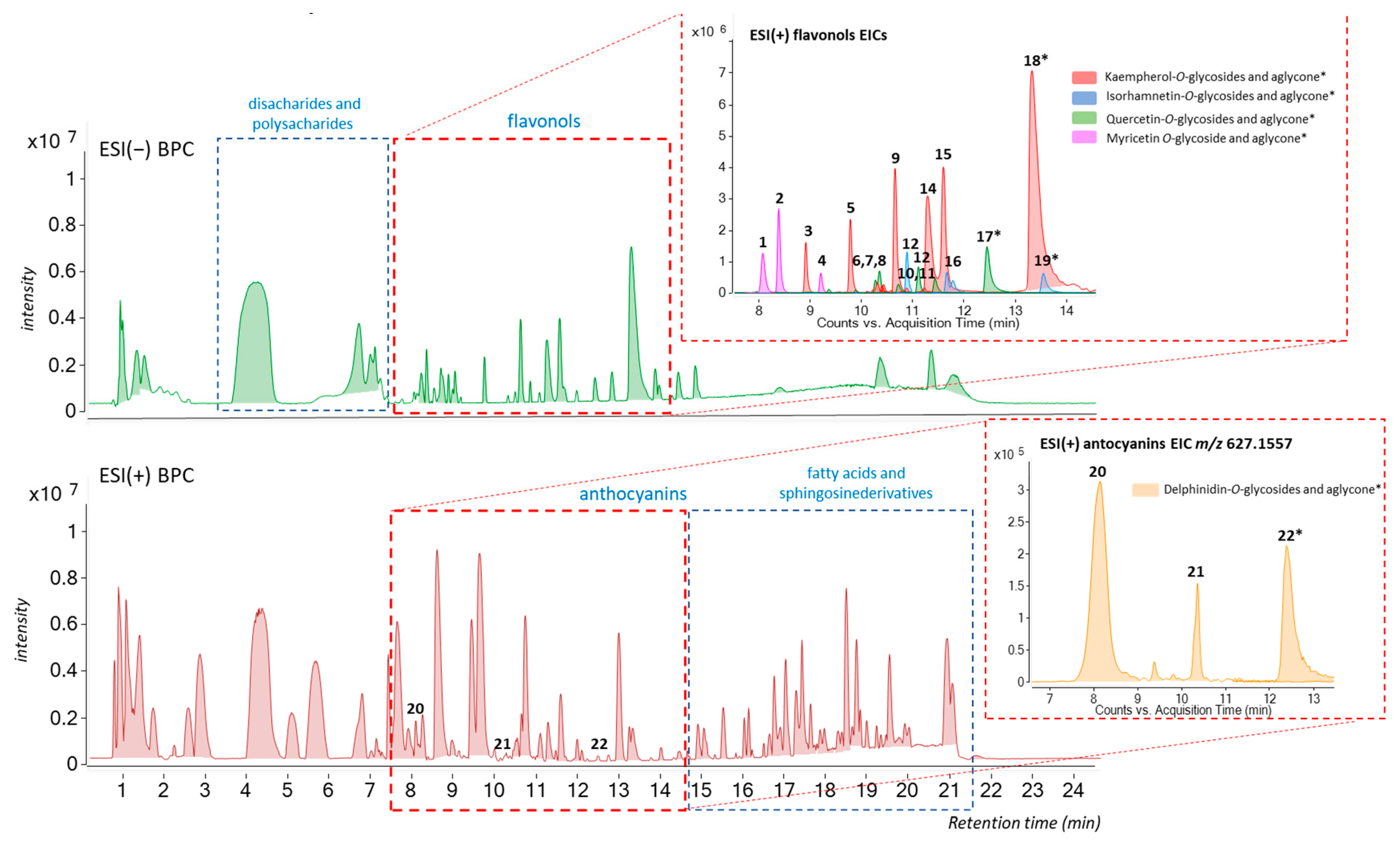
3.1.2. UHPLC-ESI(±)-QTOF-MS Analysis
3.2. In Vitro Scavenging Activity
3.3. Radical Scavenging Activity in Cell Culture (HepG2 and Hs27)
3.4. Dermo-Protective Capacity: In Vitro Enzymatic Inhibition Capacity
3.4.1. Elastase and Collagenase Inhibition Assays
3.4.2. Hyaluronidase Inhibition Assay
3.4.3. Tyrosinase Inhibition Assay
3.4.4. Xanthine Oxidase Assay
4. Conclusions
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Papaccio, F.; D′Arino, A.; Caputo, S.; Bellei, B. Focus on the contribution of oxidative stress in skin aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef]
- Widowati, W.; Fauziah, N.; Herdiman, H.; Afni, M.; Afifah, E.; Kusuma, H.S.W.; Nufus, H.; Arumwardana, S.; Rihibiha, D.D. Antioxidant and anti aging assays of Oryza sativa extracts, vanillin and coumaric acid. J. Nat. Remedies 2016, 16, 88–99. [Google Scholar] [CrossRef]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef]
- Lupu, M.; Gradisteanu Pircalabioru, G.; Chifiriuc, M.; Albulescu, R.; Tanase, C. Beneficial effects of food supplements based on hydrolyzed collagen for skin care (review). Exp. Ther. Med. 2019, 20, 12–17. [Google Scholar] [CrossRef]
- Sudha, P.N.; Rose, M.H. Beneficial effects of hyaluronic acid. Adv. Food Nutr. Res. 2014, 72, 137–176. [Google Scholar] [CrossRef] [PubMed]
- Jiratchayamaethasakul, C.; Ding, Y.; Hwang, O.; Im, S.-T.; Jang, Y.; Myung, S.-W.; Lee, J.M.; Kim, H.-S.; Ko, S.-C.; Lee, S.-H. In vitro screening of elastase, collagenase, hyaluronidase, and tyrosinase inhibitory and antioxidant activities of 22 halophyte plant extracts for novel cosmeceuticals. Fish. Aquatic Sci. 2020, 23, 6. [Google Scholar] [CrossRef]
- Skoczyńska, A.; Budzisz, E.; Trznadel-Grodzka, E.; Rotsztejn, H. Melanin and lipofuscin as hallmarks of skin aging. Postepy Dermatol. Alergol. 2017, 2, 97–103. [Google Scholar] [CrossRef]
- Boo, Y.C. Human skin lightening efficacy of resveratrol and its analogs: From in vitro studies to cosmetic applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef] [PubMed]
- Madan, K.; Nanda, S. In-vitro evaluation of antioxidant, anti-elastase, anti-collagenase, anti-hyaluronidase activities of safranal and determination of its sun protection factor in skin photoaging. Bioorg. Chem. 2018, 77, 159–167. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef]
- Serrano-Díaz, J.; Sánchez, A.M.; Maggi, L.; Martínez-Tomé, M.; García-Diz, L.; Murcia, M.A.; Alonso, G.L. Increasing the applications of Crocus sativus flowers as natural antioxidants. J. Food Sci. 2012, 77, 1162–1168. [Google Scholar] [CrossRef]
- Mottaghipisheh, J.; Mahmoodi Sourestani, M.; Kiss, T.; Horváth, A.; Tóth, B.; Ayanmanesh, M.; Khamushi, A.; Csupor, D. Comprehensive chemotaxonomic analysis of saffron crocus tepal and stamen samples, as raw materials with potential antidepressant activity. J. Pharm. Biomed. Anal. 2020, 184, 113183. [Google Scholar] [CrossRef] [PubMed]
- Verjee, S.; Garo, E.; Pelaez, S.; Fertig, O.; Hamburger, M.; Butterweck, V. Saffron flower extract promotes scratch wound closure of keratinocytes and enhances VEGF production. Planta Med. 2017, 83, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Fatehi, M.; Rashidabady, T.; Fatehi-Hassanabad, Z. Effects of Crocus sativus petals’ extract on rat blood pressure and on responses induced by electrical field stimulation in the rat isolated vas deferens and guinea-pig ileum. J. Ethnopharmacol. 2003, 84, 199–203. [Google Scholar] [CrossRef]
- Omidi, A.; Riahinia, N.; Torbati, M.B.M.; Behdani, M.-A. Hepatoprotective effect of Crocus sativus (Saffron) petals extract against acetaminophen toxicity in male wistar rats. Avicenna J. Phytomed 2014, 4, 330–336. [Google Scholar]
- Serrano-Díaz, J.; Sánchez, A.M.; Martínez-Tomé, M.; Winterhalter, P.; Alonso, G.L. Flavonoid determination in the quality control of floral bioresidues from Crocus sativus L. J. Agric. Food Chem. 2014, 62, 3125–3133. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Younesi, H.M. Antinociceptive and anti-inflammatory effects of Crocus sativus L. stigma and petal extracts in mice. BMC Pharmacol. 2002, 2, 7. [Google Scholar] [CrossRef]
- Abdelwahab, S.I.; Mariod, A.A.; Taha, M.M.E.; Zaman, F.Q.; Abdelmageed, A.H.A.; Khamis, S.; Sivasothy, Y.; Awang, K. Chemical composition and antioxidant properties of the essential oil of Cinnamomum altissimum Kosterm. (Lauraceae). Arab. J. Chem. 2014, 90, 131–135. [Google Scholar] [CrossRef]
- Muñoz Mingarro, D.; Plaza, A.; Galán, A.; Vicente, J.A.; Martínez, M.P.; Acero, N. The effect of five Taraxacum species on in vitro and in vivo antioxidant and antiproliferative activity. Food Funct. 2015, 6, 2787–2793. [Google Scholar] [CrossRef]
- Acero, N.; Gradillas, A.; Beltran, M.; García, A.; Muñoz Mingarro, D. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef]
- Acero, N.; Muñoz-Mingarro, D. Effect on tumor necrosis factor-α production and antioxidant ability of black alder, as factors related to its anti-inflammatory properties. J. Med. Food 2012, 15, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.; Tawata, S. Anti-oxidant, anti-aging, and anti-melanogenic properties of the essential oils from two varieties of Alpinia zerumbet. Molecules 2015, 20, 16723–16740. [Google Scholar] [CrossRef] [PubMed]
- Sudha, G.; Priya, M.S.; Shree, R.I.; Vadivukkarasi, S. In vitro free radical scavenging activity of raw pepino fruit (Solanum muriactum Aiton). Int. J. Curr. Pharm. Res. 2011, 3, 137–140. [Google Scholar]
- Goya, L.; Martin, M.; Ramos, S.; Mateos, R.; Bravo, L. A cell culture model for the assessment of the chemopreventive potential of dietary compounds. Curr. Nutr. Food Sci. 2009, 5, 56–64. [Google Scholar] [CrossRef][Green Version]
- Long, L.H.; Halliwell, B. Artefacts in cell culture: α-ketoglutarate can scavenge hydrogen peroxide generated by ascorbate and epigallocatechin gallate in cell culture media. Biochem. Biophys. Res. Commun. 2011, 406, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; AnilKumarReddy, B.; Hyun, C.; Lee, N. Compounds with elastase inhibition and free radical scavenging activities from Callistemon lanceolatus. J. Med. Plant Res. 2009, 3, 914–920. [Google Scholar]
- Thring, T.S.; Hili, P.; Naughton, D.P. Anti-collagenase, anti-elastase and anti-oxidant activities of extracts from 21 plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Burgos, C.; Muñoz-Mingarro, D.; Navarro, I.; Martín-Cordero, C.; Acero, N. Neuroprotective potential of verbascoside isolated from Acanthus mollis L. leaves through its enzymatic inhibition and free radical scavenging ability. Antioxidants 2020, 9, 1207. [Google Scholar] [CrossRef]
- Michalak, M. Plant-derived antioxidants: Significance in skin health and the ageing process. Int. J. Mol. Sci. 2022, 23, 585. [Google Scholar] [CrossRef]
- Zillich, O.V.; Schweiggert-Weisz, U.; Eisner, P.; Kerscher, M. Polyphenols as active ingredients for cosmetic products. Int. J. Cosmet. Sci. 2015, 37, 455–464. [Google Scholar] [CrossRef]
- Ferreira, M.; Magalhães, M.; Oliveira, R.; Sousa-Lobo, J.; Almeida, I. Trends in the use of botanicals in anti-aging cosmetics. Molecules 2021, 26, 3584. [Google Scholar] [CrossRef]
- Pantelić, M.M.; Dabić Zagorac, D.Č.; Davidović, S.M.; Todić, S.R.; Bešlić, Z.S.; Gašić, U.M.; Tešić, Ž.Lj.; Natić, M.M. Identification and quantification of phenolic compounds in berry skin, pulp, and seeds in 13 grapevine varieties grown in Serbia. Food Chem. 2016, 211, 243–252. [Google Scholar] [CrossRef]
- Domaszewska-Szostek, A.; Puzianowska-Kuźnicka, M.; Kuryłowicz, A. Flavonoids in skin senescence prevention and treatment. Int. J. Mol. Sci. 2021, 22, 6814. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Oskoueian, E.; Hendra, R.; Jaafar, H.Z.E. Evaluation of Crocus sativus L. stigma phenolic and flavonoid compounds and its antioxidant activity. Molecules 2010, 15, 6244–6256. [Google Scholar] [CrossRef]
- Diaconeasa, Z.; Știrbu, I.; Xiao, J.; Leopold, N.; Ayvaz, Z.; Danciu, C.; Ayvaz, H.; Stǎnilǎ, A.; Nistor, M.; Socaciu, C. Anthocyanins, vibrant color pigments, and their role in skin cancer prevention. Biomedicines 2020, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Moratalla-López, N.; Bagur, M.J.; Lorenzo, C.; Martínez-Navarro, M.E.; Salinas, M.R.; Alonso, G.L. Bioactivity and bioavailability of the major metabolites of Crocus sativus L. flower. Molecules 2019, 24, 2827. [Google Scholar] [CrossRef] [PubMed]
- Termentzi, A.; Kokkalou, E. LC-DAD-MS (ESI+) analysis and antioxidant capacity of Crocus sativus petal extracts. Planta Med. 2008, 74, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-K.; Shibamoto, T. Antioxidant assays for plant and food components. J. Agric. Food Chem. 2009, 57, 1655–1666. [Google Scholar] [CrossRef]
- Sariri, R.; Sabbaghzadeh, R.; Poumohamad, F. In-vitro antioxidant and anti-tyrosinase activity of methanol extracts from Crocus sativus flowers. Pharmacologyonline 2011, 3, 1–11. [Google Scholar]
- Montoro, P.; Maldini, M.; Luciani, L.; Tuberoso, C.I.G.; Congiu, F.; Pizza, C. Radical scavenging activity and LC-MS metabolic profiling of petals, stamens, and flowers of Crocus sativus L. J. Food Sci. 2012, 77, C893–C900. [Google Scholar] [CrossRef]
- Deng, S.-P.; Yang, Y.-L.; Cheng, X.-X.; Li, W.-R.; Cai, J.-Y. Synthesis, spectroscopic study and radical scavenging activity of kaempferol derivatives: Enhanced water solubility and antioxidant activity. Int. J. Mol. Sci. 2019, 20, 975. [Google Scholar] [CrossRef]
- Sun, C.; Nile, S.H.; Zhang, Y.; Qin, L.; El-Seedi, H.R.; Daglia, M.; Kai, G. Novel insight into utilization of flavonoid glycosides and biological properties of saffron (Crocus sativus L.) flower byproducts. J. Agric. Food Chem. 2020, 68, 10685–10696. [Google Scholar] [CrossRef]
- Tena, N.; Martín, J.; Asuero, A.G. State of the art of anthocyanins: Antioxidant activity, sources, bioavailability, and therapeutic effect in human health. Antioxidants 2020, 9, 451. [Google Scholar] [CrossRef]
- Sarici, G.; Cinar, S.; Armutcu, F.; Altınyazar, C.; Koca, R.; Tekin, N. Oxidative stress in Acne vulgaris. J. Eur. Acad. Dermatol. Venereol. 2009, 24, 763–767. [Google Scholar] [CrossRef]
- Lee, J.-C.; Kim, H.-R.; Kim, J.; Jang, Y.-S. Antioxidant property of an ethanol extract of the stem of Opuntia ficus-indica Var. Saboten. J. Agric. Food Chem. 2002, 50, 6490–6496. [Google Scholar] [CrossRef]
- Dimitrić Marković, J.M.; Pejin, B.; Milenković, D.; Amić, D.; Begović, N.; Mojović, M.; Marković, Z.S. Antiradical activity of delphinidin, pelargonidin and malvin towards hydroxyl and nitric oxide radicals: The energy requirements calculations as a prediction of the possible antiradical mechanisms. Food Chem. 2017, 218, 440–446. [Google Scholar] [CrossRef]
- Cai, J.; Peng, J.; Feng, J.; Li, R.; Ren, P.; Zang, X.; Wu, Z.; Lu, Y.; Luo, L.; Hu, Z.; et al. Antioxidant hepatic lipid metabolism can be promoted by orally administered inorganic nanoparticles. Nat. Commun. 2023, 14, 3643. [Google Scholar] [CrossRef] [PubMed]
- Casas-Grajales, S.; Muriel, P. Antioxidants in liver health. World J. Gastrointest. Pharmacol. Ther. 2015, 6, 59. [Google Scholar] [CrossRef]
- Merecz-Sadowska, A.; Sitarek, P.; Kucharska, E.; Kowalczyk, T.; Zajdel, K.; Cegliński, T.; Zajdel, R. Antioxidant properties of plant-derived phenolic compounds and their effect on skin fibroblast cells. Antioxidants 2021, 10, 726. [Google Scholar] [CrossRef] [PubMed]
- Park, R.-M.; Ahn, J.-Y.; Kim, S.Y.; Wee, J.-H.; Kim, Y.-H.; Min, J. Effect of alginate oligosaccharides on collagen expression in Hs27 human dermal fibroblast. J. Toxicol. Environ. Health Sci. 2020, 11, 327–334. [Google Scholar] [CrossRef]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of Oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef]
- Yang, S.; Lian, G. ROS and Diseases: Role in metabolism and energy supply. Mol. Cell Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef]
- Kruk, J.; Duchnik, E. Oxidative stress and skin diseases: Possible role of physical activity. Asian Pac. J. Cancer Prev. 2014, 15, 561–568. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Eble, J.A.; de Rezende, F.F. Redox-relevant aspects of the extracellular matrix and its cellular contacts via integrins. Antioxid. Redox Signal 2014, 20, 1977–1993. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Ramani, R.; Suraju, M.O. Polyphenol compounds and PKC signaling. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 2107–2121. [Google Scholar] [CrossRef] [PubMed]
- Bloom, D.A.; Jaiswal, A.K. Phosphorylation of Nrf2 at Ser40 by Protein Kinase C in response to antioxidants leads to the release of Nrf2 from INrf2, but is not required for Nrf2 Stabilization/accumulation in the nucleus and transcriptional activation of antioxidant response element-mediated NAD(P)H:Quinone Oxidoreductase-1 Gene expression. J. Biol. Chem. 2003, 278, 44675–44682. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.-C.D.; Wu, R.; Li, S.; Yang, A.Y.; Kong, A.-N. Anthocyanin delphinidin prevents neoplastic transformation of mouse skin JB6 P+ cells: Epigenetic re-activation of Nrf2-ARE pathway. AAPS J. 2019, 21, 83. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hu, Y.; Lu, L.; Zhao, Q.; Tao, X.; Ding, B.; Chen, S.; Zhu, J.; Guo, X.; Lin, Z. Myricetin attenuates hypoxic-ischemic brain damage in neonatal rats via NRF2 signaling pathway. Front. Pharmacol. 2023, 14, 1134464. [Google Scholar] [CrossRef]
- Bjørklund, G.; Shanaida, M.; Lysiuk, R.; Butnariu, M.; Peana, M.; Sarac, I.; Strus, O.; Smetanina, K.; Chirumbolo, S. Natural compounds and products from an anti-aging perspective. Molecules 2022, 27, 7084. [Google Scholar] [CrossRef]
- Kanashiro, A.; Souza, J.G.; Kabeya, L.M.; Azzolini, A.E.C.S.; Lucisano-Valim, Y.M. Elastase release by stimulated neutrophils inhibited by flavonoids: Importance of the catechol group. Z. Naturforsch C J. Biosci. 2007, 62, 357–361. [Google Scholar] [CrossRef]
- Bralley, E.; Greenspan, P.; Hargrove, J.L.; Hartle, D.K. Inhibition of hyaluronidase activity by Vitis rotundifolia. (Muscadine) berry seeds and skins. Pharm. Biol. 2007, 45, 667–673. [Google Scholar] [CrossRef]
- Smit, N.; Vicanova, J.; Pavel, S. The hunt for natural skin whitening agents. Int. J. Mol. Sci. 2009, 10, 5326–5349. [Google Scholar] [CrossRef]
- Kubo, I.; Kinst-Hori, I. Flavonols from saffron flower: Tyrosinase inhibitory activity and inhibition mechanism. J. Agric. Food Chem. 1999, 47, 4121–4125. [Google Scholar] [CrossRef]
- Rodrigues, M.V.N.; Rodrigues-Silva, C.; Boaventura, S.; Oliveira, A.S.S.; Rath, S.; Cass, Q.B. On-flow LC-MS/MS method for screening of xanthine oxidase inhibitors. J. Pharm. Biomed. Anal. 2020, 181, 113097. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-M.; Cheng, S.-F.; Shieh, P.-C.; Lee, J.-C.; Chen, J.-J.; Ho, C.-T.; Kuo, S.-C.; Kuo, D.-H.; Huang, L.-J.; Way, T.-D. The methanol extract of Euonymus laxiflorus, Rubia lanceolata and Gardenia jasminoides inhibits xanthine oxidase and reduce serum uric acid level in rats. Food Chem. Toxicol. 2014, 70, 179–184. [Google Scholar] [CrossRef]
- Nagao, A.; Seki, M.; Kobayashi, H. Inhibition of xanthine oxidase by flavonoids. Biosci. Biotechnol. Biochem. 1999, 63, 1787–1790. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Pan, J.; Gong, D. Novel insights into the inhibitory mechanism of kaempferol on xanthine oxidase. J. Agric. Food Chem. 2015, 63, 526–534. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Phenols | Total Flavonoids | Total Anthocyanins | |
|---|---|---|---|
| % | 4.59 ± 0.09 | 1.21 ± 0.03 | 0.68 ± 0.01 |
| Nº | Tentative Annotation | tR (min) | [M-H]− | MS/MS | Aglycone |
|---|---|---|---|---|---|
| Flavonols–Kaempherol | |||||
| 3 | kaempferol 3-O-sophoroside-7-O-glucoside | 8.92 | 771.2001 | [M-Hex(162)-H]− = 609.1468 [M-2Hex(162)-H]− = 446.0850 [M-2Hex(180)-H]−= 429.0823 | [M-3Hex(162)-H]−= 284.0322 |
| 5 | kaempferol 3,7-di-O-glucoside | 9.78 | 609.1471 | [M-Hex(162)-H]−= 447.0934 | [M-2Hex(162)-H]− = 285.0406 [M-2Hex-H-CO]- = 255.0294 151.0035 |
| 6 | kaempferol 3,7,4’tri-O-glucoside | 10.32 | 771.2001 | [M-Hex(162)-H]− = 609.1468 [M-2Hex(162)-H]− = 446.0850 | [M-3Hex(162)-H]−= 284.0322 [M-2Hex-H-CO]- = 255.0294 |
| 9 | kaempferol 3-O-sophoroside | 10.65 | 609.1471 | [M-Hex(180)-H]− = 429.0817 | [M-2Hex(146)-H]−= 284.0329 [M-2Hex-H-CO]−= 255.0294 151.0035 |
| 11 | kaempferol 3-O-rutinoside | 10.8 | 593.1506 | [M-Hex(146)-H]−= 447.0936 [M-Hex-H2O-H]−= 429.0817 | [M-3Hex(162)-H]− = 284.0322 [M-2Hex-H-CO]- = 255.0294 |
| 14 | kaempferol 3-O-glucoside | 11.33 | 447.0941 | -- | [M-Hex(182)-H]− = 285.0410 [M-2Hex-H-CO]−= 257.0457 151.0037 |
| 15 | kaempferol 7-O-glucoside | 11.59 | 447.0941 | -- | [M-Hex(182)-H]− = 284.0331 [M-2Hex-H-CO]−= 255.0301 151.0037 |
| 18 | kaempferol aglycone | 13.39 | 285.0411 | 239.0348, 229.0509, 185.0603, 159.0446, 143.0500, 107.0137 | |
| 3-Hydroxyflavonoids | |||||
| 1 | myricetin-3-O-glucoside | 8.10 | 479.0836 | -- | [M-Hex(162)-H]− = 317.0302 [M-Hex(162)-H2O-H]− = 299.0195 190,9988, 163.0037, 125.0247 |
| 2 | myricetin-3,7-di-O-glucoside | 8.41 | 641.1369 | [M-Hex(162)-H]−= 479.0836 [M-Hex-H2O-H]−= 461.0717 | [M-2Hex(162)-H]−= 317.0302 [M-2Hex(162)-H2O-H]− = 299.0195 [M-2Hex-2H2O-H]− = 281.0091, 190.9988 |
| 4 | quercetagetin 3,7-O-diglucoside | 9.23 | 641.1369 | [M-Hex(162)-H]− = 479.0836 461.0729, 435.0933 | [M-2Hex(162)-H]−= 317.0302 273.0405 |
| Flavonols–Quercetin | |||||
| 7 | quercetin 3,7-di-O-glucoside | 10.27 | 625.1435 | [M-Hex(162)-H]− = 463.0882 | [M-2Hex(162)-H]− = 301.0355 271.0247 |
| 8 | quercetin 3-O-sophoroside | 10.39 | 625.1435 | [M-Hex(162)-H]− = 463.0882 | [M-2Hex(162)-H]−= 301.0355 271.0247 |
| 10 | quercetin 3-O-glucoside | 11.15 | 463.0902 | -- | [M-Hex(162)-H]−= 301.0350 271.0247, 255.0296 |
| 13 | quercetin 7-O-glucoside | 11.57 | 463.0902 | -- | [M-Hex(162)-H]−= 301.0355 273.0409, 255.0297 |
| 17 | quercetin aglycone | 12.44 | 301.0359 | 273.0402, 178.9981, 151.0035, 121.0293, 107.0139 | |
| Flavonols–Isorhamnetin (7-methoxiquercetin) | |||||
| 12 | isorhamnetin 3-O-glucoside-7-O-rhamnoside | 10.90 | 623.1628 | [M-Hex(164)-H]−= 459.0926 | [M-2Hex(145)-H]− = 314.0432 [M-3Hex-CH3-H]−= 299.0195 285.04105, 271.0250, 151.0024 |
| 16 | isorhamnetin 3-O-glucoside | 11.79 | 477.1031 | -- | [M-Hex(162)-H]− = 314.0432 [M-2Hex-CH3-H]− = 299.0197 285.04105, 271.0236 |
| 19 | isorhamnetin aglycone | 13.56 | 315.0519 | [M-CH3-H]− = 300.0280, 283.0255.271.0250, 255.0310 | |
| Nº | Tentative Annotation | tR (min) | [M]+ | MS/MS | aglycone |
| Anthocyanins | |||||
| 20 | delphinidin 3,5-di-O-glucoside | 8.0 | 627.1550 | [M-Hex(162)]+ = 465.1024 | [M-2Hex(162)]+ = 303.0490 |
| 21 | delphinidin 3-O-sophoroside | 10.20 | 627.1550 | [M-Hex(162)]+ = 465.1022 | [M-2Hex(162)]+ = 303.0500 |
| 22 | delphinidin aglycone | 12.46 | 303.0492 | 285.0392, 257.0440, 229.0487, 201.0537 | |
| Radical | C. sativus Extract IC50 | Ascorbic Acid IC50 | Gallic Acid IC50 |
| DPPH | 887.87 ± 10.39 µg/mL | 21.18 ± 0.21 µg/mL | 0.803 ± 0.001 µg/mL |
| ABTS | 18.4 ± 0.22 µg/mL | 0.31 ± 0.01 µg/mL | 0.91 ± 0.02 µg/mL |
| O2−• | 58.46 ± 2.71 µg/mL | 11.64 ± 0.30 µg/mL | 2.30 ± 0.05 µg/mL |
| O2− | C. sativus Extract | Gallic Acid | Ascorbic Acid |
| IC50 (µg/mL) | 82.64 ± 3.32 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acero, N.; Muñoz-Mingarro, D.; Gradillas, A. Effects of Crocus sativus L. Floral Bio-Residues Related to Skin Protection. Antioxidants 2024, 13, 358. https://doi.org/10.3390/antiox13030358
Acero N, Muñoz-Mingarro D, Gradillas A. Effects of Crocus sativus L. Floral Bio-Residues Related to Skin Protection. Antioxidants. 2024; 13(3):358. https://doi.org/10.3390/antiox13030358
Chicago/Turabian StyleAcero, Nuria, Dolores Muñoz-Mingarro, and Ana Gradillas. 2024. "Effects of Crocus sativus L. Floral Bio-Residues Related to Skin Protection" Antioxidants 13, no. 3: 358. https://doi.org/10.3390/antiox13030358
APA StyleAcero, N., Muñoz-Mingarro, D., & Gradillas, A. (2024). Effects of Crocus sativus L. Floral Bio-Residues Related to Skin Protection. Antioxidants, 13(3), 358. https://doi.org/10.3390/antiox13030358