1. Introduction
Nutritional balance is closely related to human health, and fish is crucial for the human diet due to its rich nutritional content. Fish meat is a source of rich and high-quality protein that is more accessible and affordable than other animal proteins [
1,
2]. Since the 1950s, aquaculture production increased globally, while fishery production remained consistent since the early 1990s [
3]. Moreover, the production of aquaculture for human consumption exceeded that of the fisheries in 2016 [
3]. Aquaculture plays an important role in world food security by providing aquatic protein [
4]. According to a report by FAO [
5], the production of freshwater fish is at least five times that of marine fish, demonstrating the important role of freshwater fish farming in food security.
Micropterus salmoides (
M. salmoides) is a native to North America and was imported into China for farming as food fish.
M. salmoides has fast growth, strong adaptability, and delicious meat [
6]; therefore, its production increased from 243,196 to 802,486 tons during the last decade in China [
7]. However, the increased production necessitates sufficient feed and raw materials for culturing
M. salmoides. Fishmeal (FM) is one of the most important, expensive, and high-quality sources of protein due to its good palatability, amino acid balance, and high nutritional value [
8]. However, FM production and its price are projected to increase by 1% and 30%, respectively, by the year 2030 [
9], which may impede the sustainable development of the aquaculture industry. As a carnivorous fish,
M. salmoides requires 40–50% more FM than herbivorous and omnivorous fish [
10]. Therefore, reducing the proportion of FM in
M. salmoides feed is crucial to overcoming the FM production challenge, thereby promoting the sustainable farming of
M. salmoides.
To reduce the FM content, animal- and plant-based protein sources are used as important ingredients in aquatic animal feed. However, due to an imbalance of essential amino acids (EAAs), their extensive use results in several adverse effects [
11,
12,
13]. Therefore, it is important to balance the essential nutrients, including EAAs, in the feed to replace FM with animal- and plant-based proteins without affecting the production performance of the fish. A potential alternate protein source is poultry by-products, as they have similar protein contents, reasonable prices, and a stable supply [
14]. However, one of the limiting amino acids in poultry by-product feed is methionine [
15], which should be included in aquatic animal feed to reduce the adverse impact on growth, antioxidant capacity, and immune function resulting from reduced FM content [
16].
Supplementation with EAAs, particularly methionine (Met), was considered an effective way of replacing FM with animal- and plant-based proteins in aquatic feeds [
12,
17,
18]. Methionine products are commercially available in various forms, such as DL-methionine (DL-Met) or Met-Met. Met-Met has an advantage over DL-Met in that it is insoluble in water and absorbed easily [
19,
20]. Guo et al. found that Met-Met supplementation could improve the growth performance and antioxidant capacity of Nile tilapia (
Oreochromis niloticus) [
20]. Mamauag et al. reported similar utilization effects of DL-Met and Met-Met by Red Sea Bream (
Pagrus major) larvae and juveniles [
21]. Met-Met is mainly used to supplement the feed of
Litopenaeus vannamei (
L. vannamei) with a higher availability than DL-Met [
19]. Dietary supplementation with 0.34% of Met-Met could reduce the FM content from 18 to 6% without any adverse effect on the performance of
L. vannamei [
22]. Moreover, Met-Met supplementation ameliorates the negative effects on growth caused by limiting Met content in low-FM diets and effectively improves the immune and antioxidant capacity of
L. vannamei [
23]. However, considering the differences between shrimp and fish, the present study investigated the effects of Met-Met supplementation on the growth, intestinal antioxidant capacity, immunity, and microbiota of
M. salmoides. The outcomes would facilitate the further evaluation of the application of Met-Met in different species, thereby limiting the FM content to promote the sustainable development of
M. salmoides.
4. Discussion
Fishmeal is widely used in aquatic animal feed due to its balanced amino acids [
30]. Compared to fishmeal, soy protein is known for its low levels of methionine [
31]. In the present study, the content of methionine in soybean meal is the lowest among several main ingredients in feed (as shown in the
Supplementary Materials). In addition, compared with the plant protein source used in this study, the essential amino acid content of poultry meal is more similar to that of fishmeal, except for lysine and methionine (as shown in the
Supplementary Materials), which are also the main limiting amino acids in poultry by-products [
32]. Therefore, in the present study, methionine was supplemented with a low-fishmeal diet to explore its application effect in
M. salmoides feed. Peptides have attracted widespread attention due to their unique transport mechanisms, leading to faster and more effective absorption rates than free-form crystalline amino acids in the intestines [
21,
33,
34]. A study on DL-Methionine (DL-Met) and Met-Met showed that dietary Met-Met supplementation was more effective than DL-Met in
L. vannamei [
35]. Our results showed that growth-related indicators and feed utilization were decreased by reducing FM from 40 to 25%. However, compared to 25% FM, the supplementation of 25% FM with 0.40% DL-Met or 0.10% Met-Met significantly improved FW, WGR, and SGR. These results suggested the role of DL-Met or Met-Met supplementation in alleviating the negative effects of low-FM diets. Previous studies showed that DL-Met or Met-Met could promote the growth of larvae, juvenile Red Sea Bream [
21], and
L. vannamei [
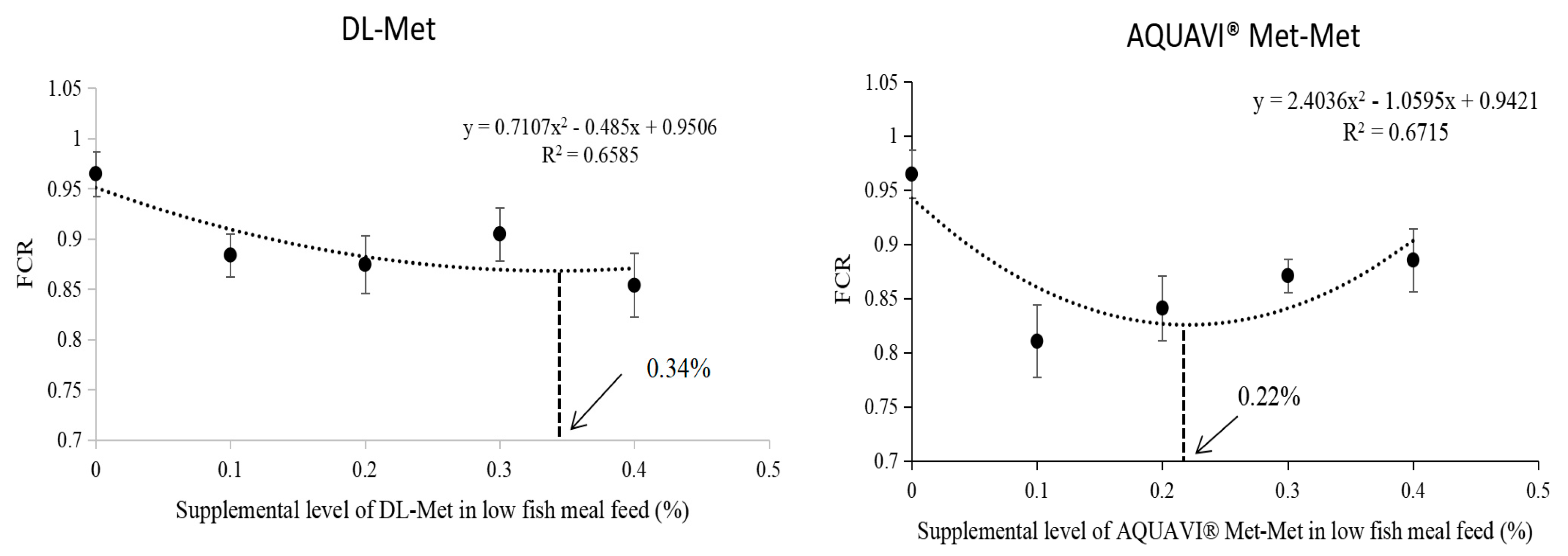
35]. According to the quadratic regression analysis of the FCR index, 0.34% DL-Met or 0.22% Met-Met added to LF meal resulted in similar growth and feed utilization by
M. salmoides as those feeding on high FM. Therefore, it can be concluded that the
M. salmoides diet with low FM required a higher level of DL-Met supplementation than that of Met-Met to induce desirable growth. In other words,
M. salmoides shows better utilization of Met-Met than DL-Met. In addition, Xie et al. reported that an improved growth performance of
L. vanname could be achieved by adding 0.10% of Met-Met or 0.30% of DL-Met to a low-FM diet [
35], which further validated our results. Moreover, in the present study, no significant difference was observed in the whole-body and amino acid composition of
M. salmoides among the groups. The results were similar to those reported in the previous studies on
L. vannamei [
36] and Red Sea Bream [
21].
Dietary low-FM contents lead to oxidative stress in
M. salmoides [
37], which is relieved by antioxidant enzymes and related genes mainly through the Nrf2 signaling pathway [
38,
39]. Oxidative stress results from the overproduction of reactive oxygen species (ROS), which are removed by the action of antioxidant enzymes (such as SOD, GSH-Px, and CAT) [
40]. One of the important indicators of oxidative damage in the body is the increased level of MDA [
16]. In the present study, relative to the HF diet, feeding with the LF diet significantly reduced the intestinal CAT, SOD, and GSH levels, whereas the MDA content was increased. This suggests that consuming a low-FM diet could reduce the antioxidant ability of the intestines of
M. salmoides, resulting in oxidative damage. Furthermore, in comparison to the LF group, CAT, SOD, and GSH levels in the intestines of
M. salmoides increased significantly when DL-Met or Met-Met were added to LF diets. Specifically, 0.20–0.40% of DL-Met or 0.10–0.40% of Met-Met significantly reduced MDA content relative to the LF group. Previous studies on Nile tilapia [
20] and
L. vannamei [
23] reported that 0.20% and 0.10–0.15% of Met-Met increased the antioxidant capacity, respectively. Ji et al. [
23] also reported that 0.10–0.25% of Met-Met significantly reduced the MDA content in
L. vannamei. Collectively, these results indicated that supplementing with DL-Met or Met-Met could effectively alleviate the intestinal oxidative damage caused by a low-FM diet. Particularly, 0.10% of Met-Met increased GSH and decreased MDA content in comparison to the HF group, suggesting a more effective role of Met-Met than that of DL-Met.
The nrf2-mediated gene expression of antioxidant enzymes reduces external oxidative stress [
41]. In the present study, relative to the HF group, the LF diet significantly reduced the expressions of
nrf2,
cat, and
sod genes, whereas the
keap1 gene expression levels were significantly increased. According to a study, dietary low FM could reduce the antioxidant capacity in juvenile golden pompano (
Trachinotus ovatus) by decreasing the
nrf2 expression level in the nrf2 signaling pathway, increasing the
keap1 expression level, thereby inhibiting the expression of oxidative stress-related genes, such as
sod,
cat, and
gsh-px [
42]. Similarly, a low-FM diet for
M. salmoides could downregulate the intestinal antioxidant capacity through the nrf2-mediated expression of antioxidant enzymes and related genes. Compared with the LF group, 0.20–0.40% of DL-Met supplementation significantly upregulated
nrf2 and
sod genes expression and downregulated
keap1 gene expression, whereas 0.10–0.40% of Met-Met the upregulated
nrf2,
cat, and
sod and downregulated the
keap1 gene expression in the present study. Conclusively, DL-Met or Met-Met improved the antioxidant capacity of
M. salmoides by regulating the activity of related enzymes and genes of the Nrf2 signaling pathway.
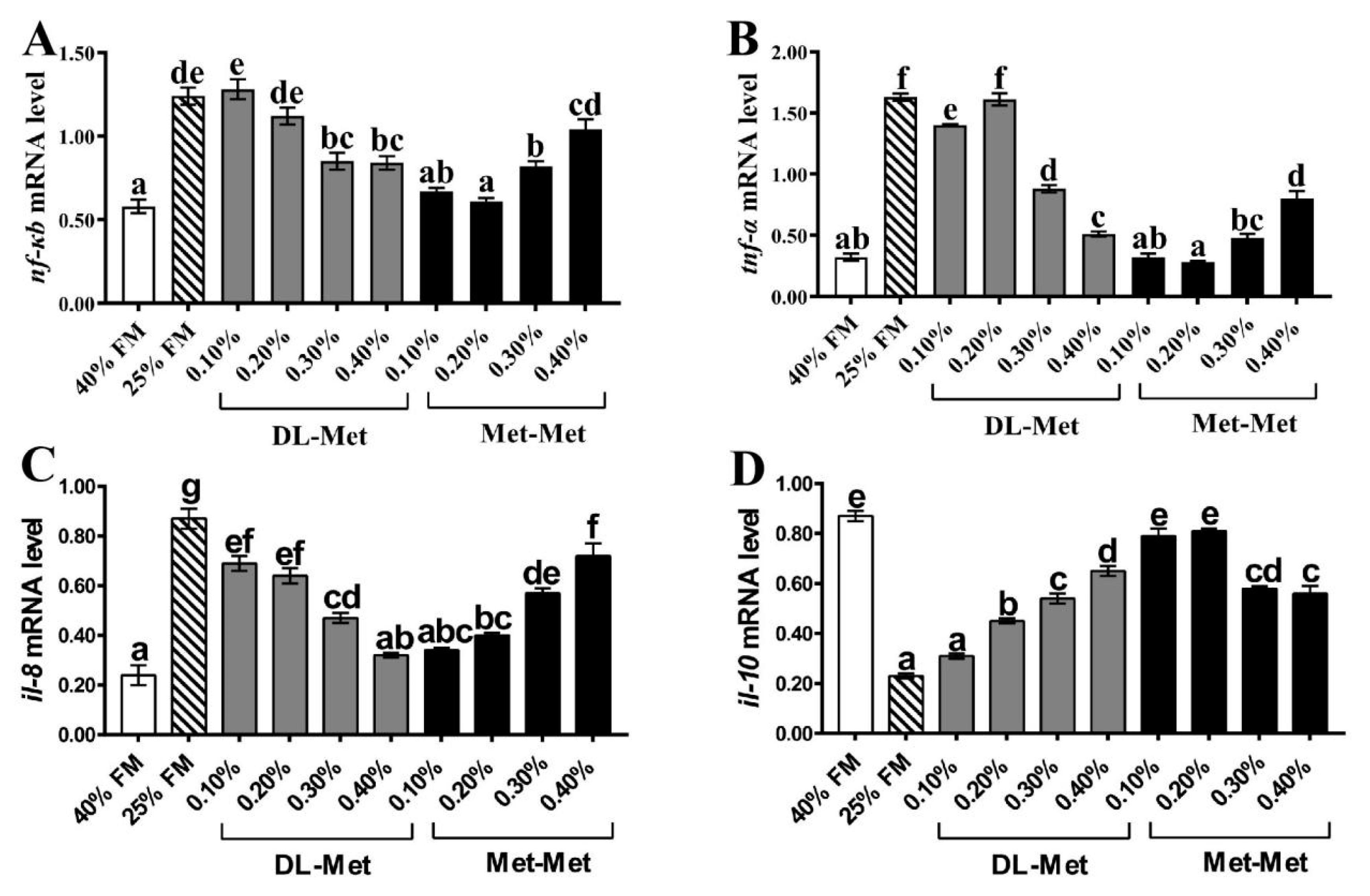
The nuclear factor (nf-κb) can regulate inflammation through cytokines. As a transcription factor, it regulates the expression of pro-inflammatory genes, including
tnf-α and
il-8, and anti-inflammatory genes, such as
il-10 [
43,
44]. In the present study, significantly upregulated levels of the
nf-κb,
tnf-α, and
il-8 genes and downregulated levels of
il-10 were induced by the LF diet relative to the HF group, which is consistent with the results of a previous study [
45]. In addition, our results showed that in comparison to the LF group, supplementation with DL-Met or Met-Met significantly down- and upregulated the pro- and anti-inflammatory genes, respectively, in the intestine of
M. salmoides. The above results indicated that consumption of a low-FM diet could lead to intestinal inflammation in
M. salmoides, and DL-Met or Met-Met supplementation could alleviate the inflammatory response.
The intestinal microbiome is crucial for human health and is substantially influenced by diet [
46]. Similarly, in fish, intestinal microbiota plays an important role in nutrition, immunity, and resistance to invading pathogens, and diet greatly influences its composition [
47]. According to a study on Nile tilapia, the addition of Met-Met to fish feed could influence the composition of intestinal microbiota [
20]. The core microbiota are closely related to the host genotype and remain unaffected by the external environment [
48,
49]. In the present study, the intestinal tissue samples of
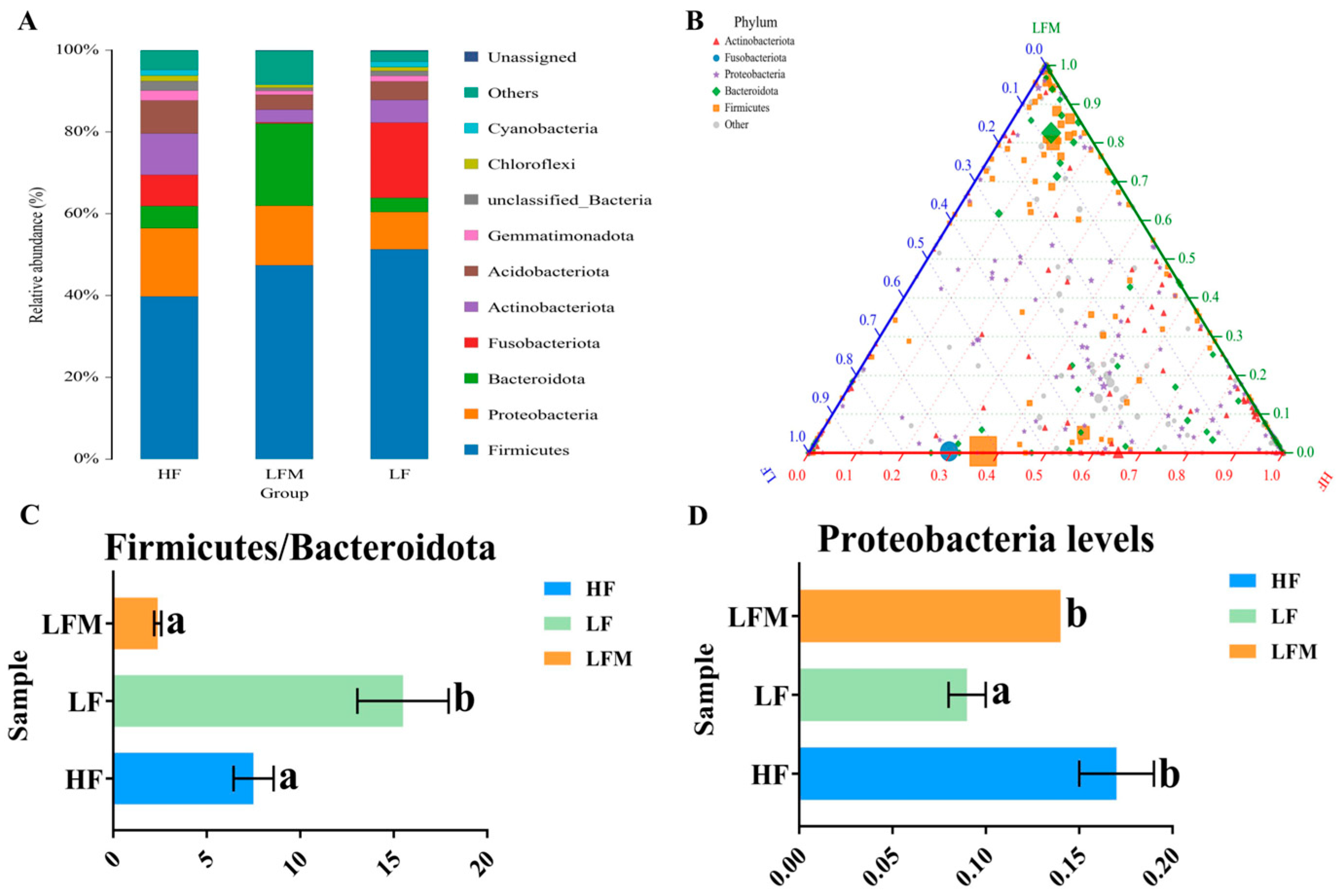
M. salmoides fed with the HF, LF, and LFM diets were used for microbiological analysis. The ACE and Simpson and Shannon indices in α-diversity are commonly used to evaluate the species richness and diversity. Our results did not show a significant difference in ACE among the groups, and the lowest values of the Simpson and Shannon index were observed in the LF group, which were significantly increased by Met-Met supplementation. This suggests the potential of Met-Met in enhancing the diversity of intestinal microbiota. PCoA, UPGMA, and sample clustering heatmaps for β-diversity analysis revealed that the microorganisms in the three random samples within the groups were similar; however, there were significant differences in the microbiota among the groups.
Nevertheless, an imbalanced Firmicutes-to-Bacteroidota ratio could lead to pathogenic invasion [
50]. In the present study, the Firmicutes-to-Bacteroidota ratio of the LF group was significantly higher than that of the HF and LFM groups. This suggested that the low-FM diet may lead to an imbalanced Firmicutes-to-Bacteroidota ratio in the intestine of
M. salmoides, which was more likely to promote pathogenic invasion, thereby reducing the immunity of fish. However, the supplementation of Met-Met in a low-FM diet could improve intestinal immunity by regulating the Firmicutes-to-Bacteroidota ratio. Furthermore, Proteobacteria is the core intestinal microbiota of fish [
51,
52]. Previous studies on the intestinal tissue samples of grass carp (
Ctenopharyngodon idella) revealed that Proteobacteria is positively correlated with the anti-inflammatory factor
tgf-β, suggesting its role in improving the anti-inflammatory ability of fish [
53]. Our results showed significantly lower levels of Proteobacteria in the LM group relative to the HF and LFM groups. Moreover, the upregulation of pro-inflammatory factors and downregulation of anti-inflammatory factors might be induced by the imbalanced Firmicutes-to-Bacteroidota ratio or reduced levels of Proteobacteria in a low-FM diet. However, the relevant regulatory mechanisms of bacterial abundance and inflammatory factors need further investigation. The results also demonstrated that Met-Met may improve intestinal health by increasing the abundance of beneficial bacteria. Additionally, the results of the ternary plot showed that Met-Met changed the bacterial diversity, whereas the LEfSe analysis revealed that the
unique Roseburia [
54,
55],
Lachnospiraceae_NK4A136_group [
56], and
unclassified_Oscillospiraceae [
57] in the LFM group were beneficial bacteria.
Roseburia was negatively correlated with MDA level and positively correlated with antioxidant enzymes, including SOD in the gut of mice [
58]. Moreover, it plays an anti-inflammatory role in alleviating colitis pathology, suggesting its potential anti-inflammatory effects [
59]. Similarly,
Lachnospiraceae_NK4A136_group found in the gut of mice was positively correlated with anti-inflammatory genes and negatively correlated with the pro-inflammatory and oxidative stress factors [
60]. Therefore, it can be concluded that changes in the abundance of
Roseburia and
Lachnospiraceae_NK4A136_group may have resulted from Met-Met supplementation, thereby enhancing the intestinal oxidation and anti-inflammatory capacity of
M. salmoides. Notably,
unclassified Oscillospiraceae produces butyrate after fermentation in the gut, which helps in the utilization of dietary fiber [
61]. The differences in genus abundance between the two groups were analyzed by Metastats. In the present study, compared with the LF and HF groups, the abundance of probiotic bacteria, such as
Caldicoprobacter,
Pseudogracilibacillus, and
Parasutterella, was increased in the LFM group. Moreover,
Caldicoprobacter could ferment complex carbohydrates in the intestine to produce lactic acid, thereby promoting the production of short-chain fatty acids to maintain intestinal health [
62,
63].
Pseudogracilibacilluse and
Parasutterella are considered beneficial bacteria [
64,
65]. In addition, anti-inflammatory effects may be exerted by the metabolites (7-ketodeoxycholic acid and haloperidol glucuronide) produced by
Parasutterella [
66]. This implies that the supplementation of Met-Met in a low-FM diet could improve the composition and abundance of beneficial bacteria in the gut, thereby improving intestinal health.



 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}